A Shift in Glycerolipid Metabolism Defines the Follicular Fluid of IVF Patients with Unexplained Infertility

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Materials and Reagents
2.3. Sample Preparation for LC-MS Analysis
2.4. Ultra-HPLC Quadrupole Time-of-Flight MS Analysis
2.5. Data Analysis
3. Results
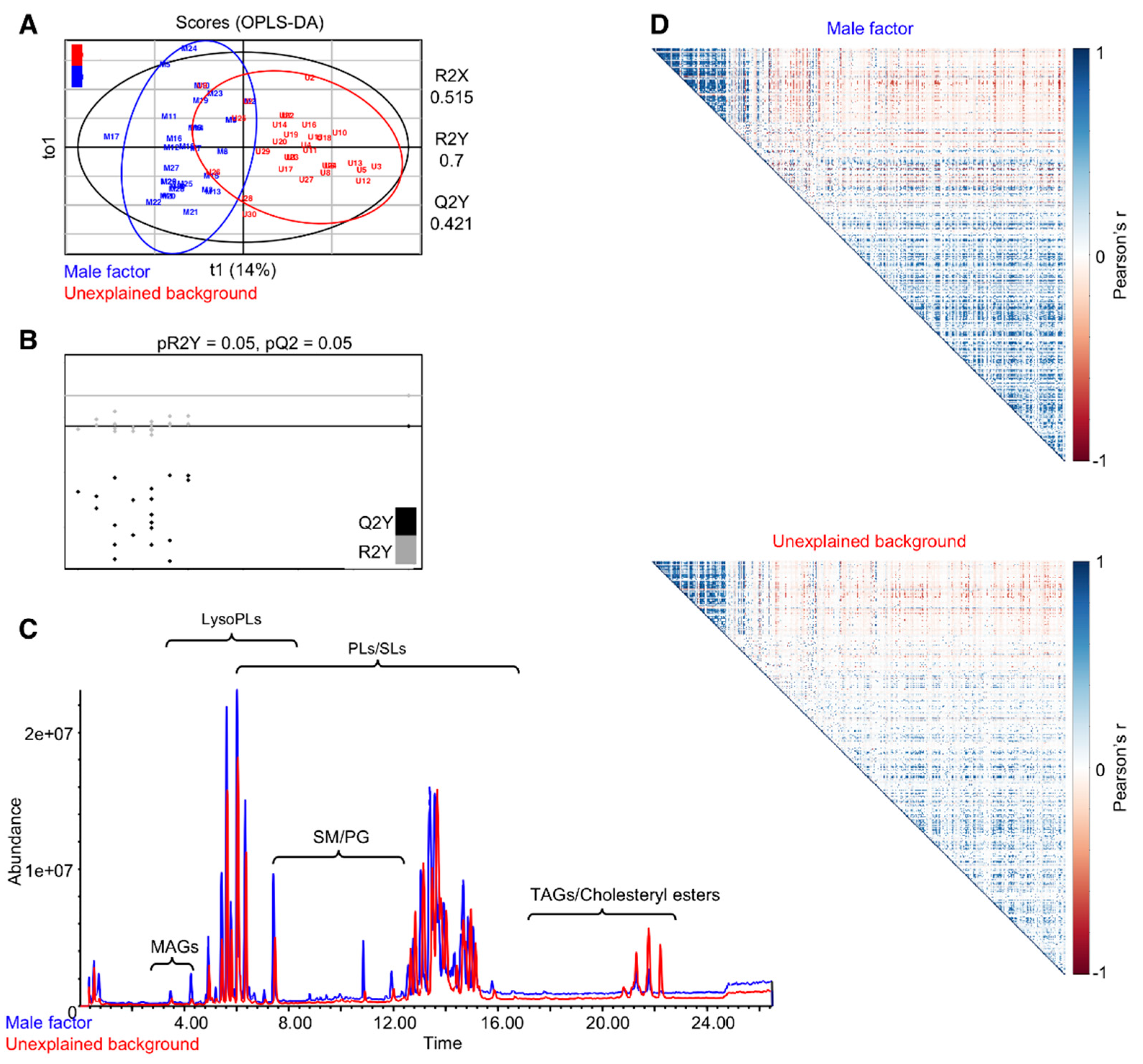
3.1. Collection of FF Samples from IVF Patients with Unexplained Infertility or Male Factor Background
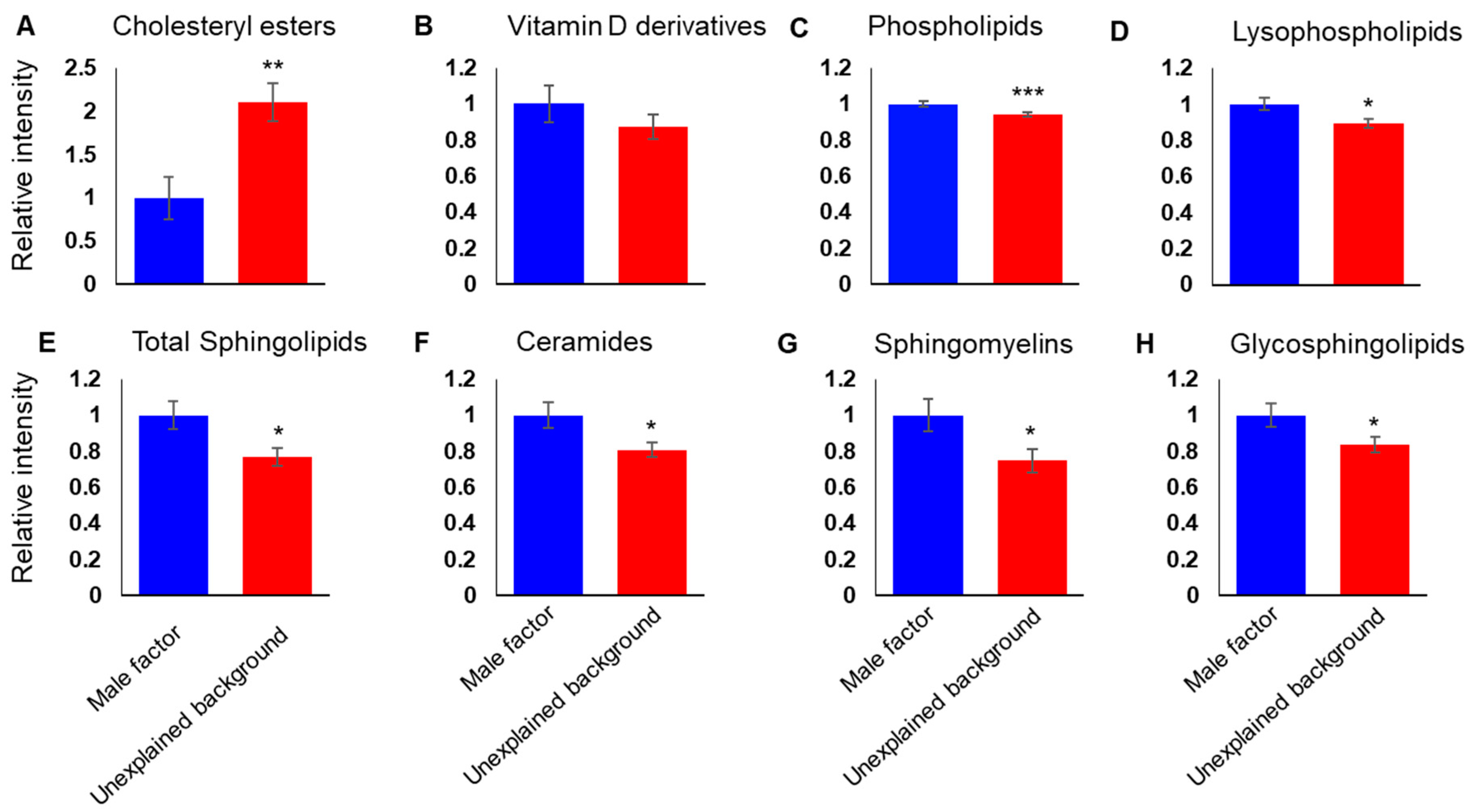
3.2. A Differential Lipid Composition of the FF of IVF Patients with Unexplained Infertility
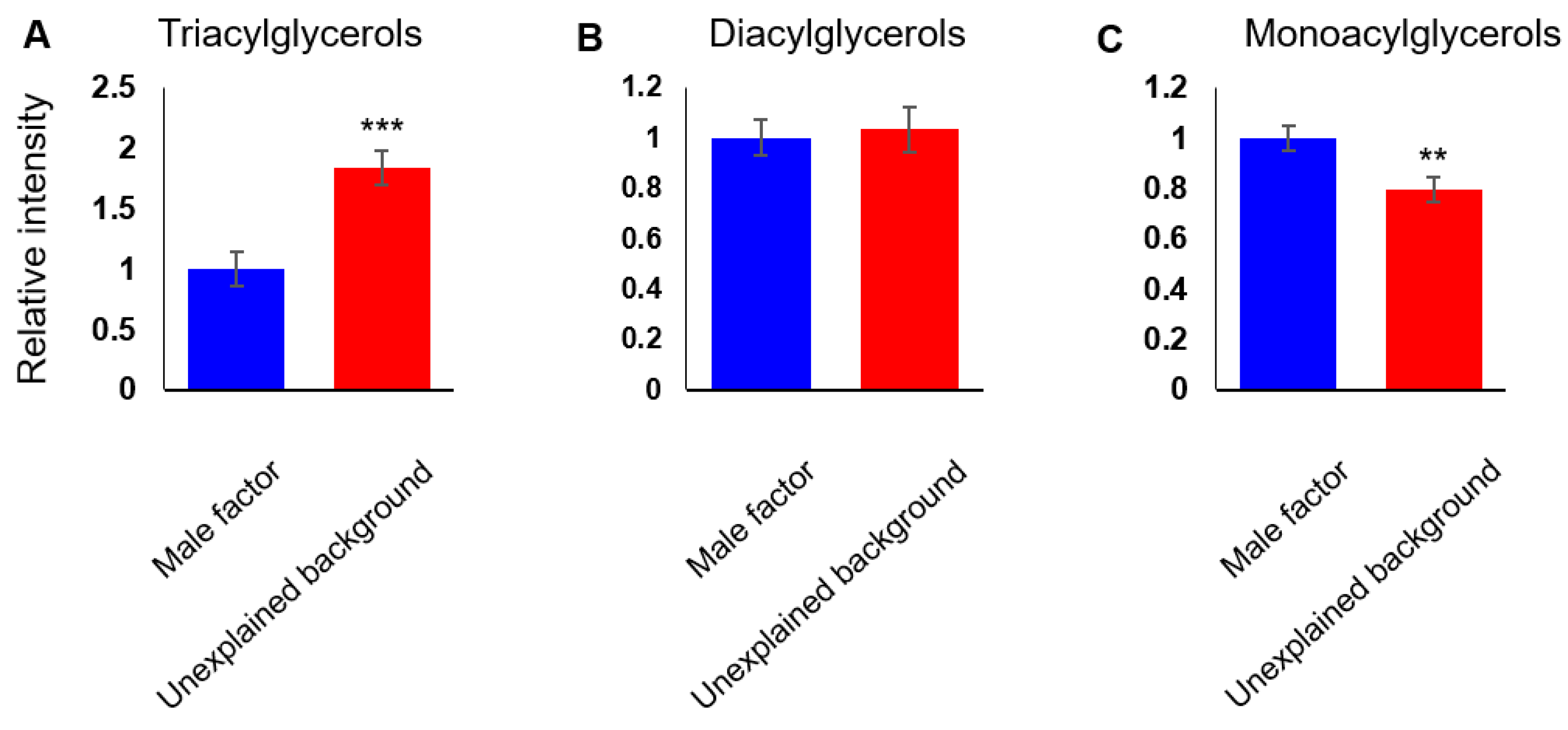
3.3. Glycerolipid Metabolism Shifts From Monoacylglycerols to Triacylglycerols
3.4. Potential Lipid Biomarkers for Pregnancy Outcome Show Discriminant Accumulation in the FF of IVF Patients with Unexplained Infertility
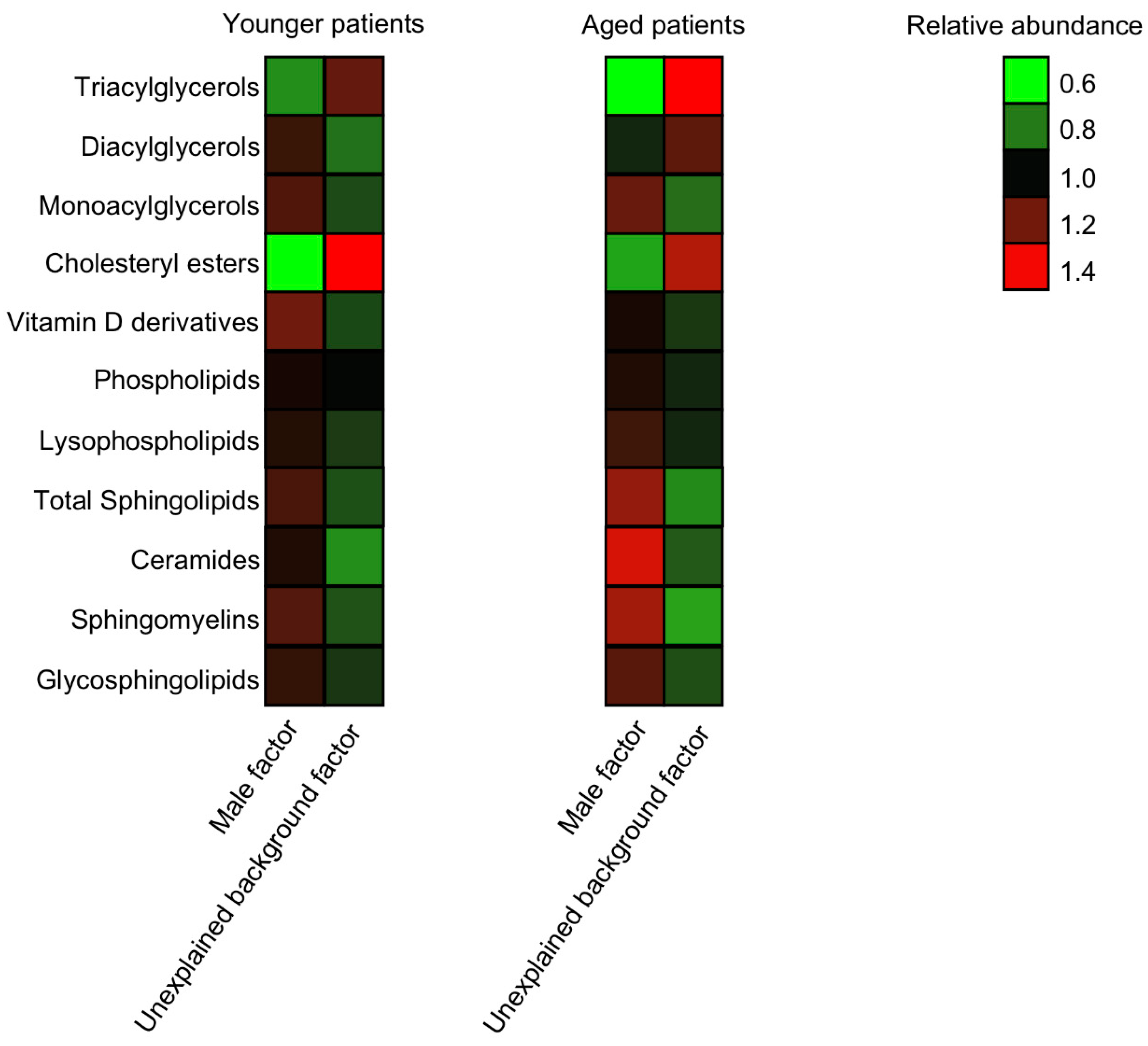
3.5. The Lipid Signature of Patients with Unexplained Infertility is not Age- or BMI-Dependent
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Revelli, A.; Delle Piane, L.; Casano, S.; Molinari, E.; Massobrio, M.; Rinaudo, P. Follicular fluid content and oocyte quality: From single biochemical markers to metabolomics. Reprod. Biol. Endocrinol. 2009, 7, 40–53. [Google Scholar] [CrossRef] [PubMed]
- O’Gorman, A.; Wallace, M.; Cottell, E.; Gibney, M.J.; McAuliffe, F.M.; Wingfield, M.; Brennan, L. Metabolic profiling of human follicular fluid identifies potential biomarkers of oocyte developmental competence. Reproduction 2013, 146, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Bender, K.; Walsh, S.; Evans, A.C.; Fair, T.; Brennan, L. Metabolite concentrations in follicular fluid may explain differences in fertility between heifers and lactating cows. Reproduction 2010, 139, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, F.B.; Montani, D.A.; Pilau, E.J.; Gozzo, F.C.; Fraietta, R.; Turco, E.G.L. Ovarian environment aging: Follicular fluid lipidomic and related metabolic pathways. J. Assist. Reprod. Genet. 2018, 35, 1385–1393. [Google Scholar] [CrossRef]
- de la Barca, J.M.C.; Boueilh, T.; Simard, G.; Boucret, L.; Ferre-L’Hotellier, V.; Tessier, L.; Gadras, C.; Bouet, P.E.; Descamps, P.; Procaccio, V.; et al. Targeted metabolomics reveals reduced levels of polyunsaturated choline plasmalogens and a smaller dimethylarginine/arginine ratio in the follicular fluid of patients with a diminished ovarian reserve. Hum. Reprod. 2017, 32, 2269–2278. [Google Scholar] [CrossRef]
- Valckx, S.D.; De Pauw, I.; De Neubourg, D.; Inion, I.; Berth, M.; Fransen, E.; Bols, P.E.; Leroy, J.L. BMI-related metabolic composition of the follicular fluid of women undergoing assisted reproductive treatment and the consequences for oocyte and embryo quality. Hum. Reprod. 2012, 27, 3531–3539. [Google Scholar] [CrossRef]
- Pacella, L.; Zander-Fox, D.L.; Armstrong, D.T.; Lane, M. Women with reduced ovarian reserve or advanced maternal age have an altered follicular environment. Fertil. Steril. 2012, 98, 986–994. [Google Scholar] [CrossRef]
- Von Wald, T.; Monisova, Y.; Hacker, M.R.; Yoo, S.W.; Penzias, A.S.; Reindollar, R.R.; Usheva, A. Age-related variations in follicular apolipoproteins may influence human oocyte maturation and fertility potential. Fertil. Steril. 2010, 93, 2354–2361. [Google Scholar] [CrossRef]
- Valckx, S.D.; Arias-Alvarez, M.; De Pauw, I.; Fievez, V.; Vlaeminck, B.; Fransen, E.; Bols, P.E.; Leroy, J.L. Fatty acid composition of the follicular fluid of normal weight, overweight and obese women undergoing assisted reproductive treatment: A descriptive cross-sectional study. Reprod. Biol. Endocrinol. 2014, 12, 13–31. [Google Scholar] [CrossRef]
- Mirabi, P.; Chaichi, M.J.; Esmaeilzadeh, S.; Ali Jorsaraei, S.G.; Bijani, A.; Ehsani, M.; Hashemi Karooee, S.F. The role of fatty acids on ICSI outcomes: A prospective cohort study. Lipids Health Dis. 2017, 16, 18–27. [Google Scholar] [CrossRef]
- Mirabi, P.; Chaichi, M.J.; Esmaeilzadeh, S.; Jorsaraei, S.G.A.; Bijani, A.; Ehsani, M. Does different BMI influence oocyte and embryo quality by inducing fatty acid in follicular fluid? Taiwan. J. Obstet. Gynecol. 2017, 56, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Gesink Law, D.C.; Maclehose, R.F.; Longnecker, M.P. Obesity and time to pregnancy. Hum. Reprod. 2007, 22, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Rich-Edwards, J.W.; Goldman, M.B.; Willett, W.C.; Hunter, D.J.; Stampfer, M.J.; Colditz, G.A.; Manson, J.E. Adolescent body mass index and infertility caused by ovulatory disorder. Am. J. Obstet. Gynecol. 1994, 171, 171–177. [Google Scholar] [CrossRef]
- Jungheim, E.S.; Moley, K.H. Current knowledge of obesity’s effects in the pre- and periconceptional periods and avenues for future research. Am. J. Obstet. Gynecol. 2010, 203, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Liu, H.; Gu, X.; Boots, C.; Moley, K.H.; Wang, Q. Metabolic control of oocyte development: Linking maternal nutrition and reproductive outcomes. Cell. Mol. Life Sci. 2015, 72, 251–271. [Google Scholar] [CrossRef]
- Wu, L.L.; Dunning, K.R.; Yang, X.; Russell, D.L.; Lane, M.; Norman, R.J.; Robker, R.L. High-fat diet causes lipotoxicity responses in cumulus-oocyte complexes and decreased fertilization rates. Endocrinology 2010, 151, 5438–5445. [Google Scholar] [CrossRef]
- Luzzo, K.M.; Wang, Q.; Purcell, S.H.; Chi, M.; Jimenez, P.T.; Grindler, N.; Schedl, T.; Moley, K.H. High fat diet induced developmental defects in the mouse: Oocyte meiotic aneuploidy and fetal growth retardation/brain defects. PLoS ONE 2012, 7, e49217. [Google Scholar] [CrossRef]
- Yang, X.; Wu, L.L.; Chura, L.R.; Liang, X.; Lane, M.; Norman, R.J.; Robker, R.L. Exposure to lipid-rich follicular fluid is associated with endoplasmic reticulum stress and impaired oocyte maturation in cumulus-oocyte complexes. Fertil. Steril. 2012, 97, 1438–1443. [Google Scholar] [CrossRef]
- Nandi, S.; Kumar, V.G.; Manjunatha, B.M.; Gupta, P.S. Biochemical composition of ovine follicular fluid in relation to follicle size. Dev. Growth Differ. 2007, 49, 61–66. [Google Scholar] [CrossRef]
- Gonzalez, M.B.; Lane, M.; Knight, E.J.; Robker, R.L. Inflammatory markers in human follicular fluid correlate with lipid levels and Body Mass Index. J. Reprod. Immunol. 2018, 130, 25–29. [Google Scholar] [CrossRef]
- Cataldi, T.; Cordeiro, F.B.; Costa Ldo, V.; Pilau, E.J.; Ferreira, C.R.; Gozzo, F.C.; Eberlin, M.N.; Bertolla, R.P.; Cedenho, A.P.; Turco, E.G. Lipid profiling of follicular fluid from women undergoing IVF: Young poor ovarian responders versus normal responders. Hum. Fertil. (Camb.) 2013, 16, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Wallace, M.; Cottell, E.; Gibney, M.J.; McAuliffe, F.M.; Wingfield, M.; Brennan, L. An investigation into the relationship between the metabolic profile of follicular fluid, oocyte developmental potential, and implantation outcome. Fertil. Steril. 2012, 97, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Fayezi, S.; Darabi, M.; Darabi, M.; Nouri, M.; Rahimipour, A.; Mehdizadeh, A. Analysis of follicular fluid total phospholipids in women undergoing in-vitro fertilisation. J. Obstet. Gynaecol. 2014, 34, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Irani, M.; Merhi, Z. Role of vitamin D in ovarian physiology and its implication in reproduction: A systematic review. Fertil. Steril. 2014, 102, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, S.; Jindal, S.; Greenseid, K.; Shu, J.; Zeitlian, G.; Hickmon, C.; Pal, L. Replete vitamin D stores predict reproductive success following in vitro fertilization. Fertil. Steril. 2010, 94, 1314–1319. [Google Scholar] [CrossRef]
- Ciepiela, P.; Duleba, A.J.; Kowaleczko, E.; Chelstowski, K.; Kurzawa, R. Vitamin D as a follicular marker of human oocyte quality and a serum marker of in vitro fertilization outcome. J. Assist. Reprod. Genet. 2018, 35, 1265–1276. [Google Scholar] [CrossRef]
- Liu, L.; Yin, T.L.; Chen, Y.; Li, Y.; Yin, L.; Ding, J.; Yang, J.; Feng, H.L. Follicular dynamics of glycerophospholipid and sphingolipid metabolisms in polycystic ovary syndrome patients. J. Steroid Biochem. Mol. Biol. 2018, 185, 142–149. [Google Scholar] [CrossRef]
- Shehadeh, A.; Bruck-Haimson, R.; Saidemberg, D.; Zacharia, A.; Herzberg, S.; Ben-Meir, A.; Moussaieff, A. A shift in follicular fluid from triacylglycerols to membrane lipids is associated with positive pregnancy outcome. FASEB J. 2019, 33, 10291–10299. [Google Scholar] [CrossRef]
- Thevenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the human adult urinary metabolome variations with age, body mass index and gender by implementing a comprehensive workflow for univariate and OPLS statistical analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef]
- Batushansky, A.; Toubiana, D.; Fait, A. Correlation-Based Network Generation, Visualization, and Analysis as a Powerful Tool in Biological Studies: A Case Study in Cancer Cell Metabolism. BioMed Res. Int. 2016, 2016, 1–9. [Google Scholar] [CrossRef]
- Wie, T.; Simko, V. R package “corrplot”: Visualization of a Correlation Matrix (Version 0.84). Available online: https://github.com/taiyun/corrplot (accessed on 22 June 2020).
- Batushansky, A.; Matsuzaki, S.; Newhardt, M.F.; West, M.S.; Griffin, T.M.; Humphries, K.M. GC-MS metabolic profiling reveals fructose-2,6-bisphosphate regulates branched chain amino acid metabolism in the heart during fasting. Metabolomics 2019, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Donovan, E.L.; Lopes, E.B.P.; Batushansky, A.; Kinter, M.; Griffin, T.M. Independent effects of dietary fat and sucrose content on chondrocyte metabolism and osteoarthritis pathology in mice. Dis. Model Mech. 2018, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kotze, H.L.; Armitage, E.G.; Sharkey, K.J.; Allwood, J.W.; Dunn, W.B.; Williams, K.J.; Goodacre, R. A novel untargeted metabolomics correlation-based network analysis incorporating human metabolic reconstructions. BMC Syst. Biol. 2013, 7, 107. [Google Scholar] [CrossRef] [PubMed]
- Perez de Souza, L.; Alseekh, S.; Brotman, Y.; Fernie, A.R. Network based strategies in metabolomics data analysis and interpretation: From molecular networking to biological interpretation. Expert Rev. Proteom. 2020, 17, 243–255. [Google Scholar] [CrossRef]
- Vander Borght, M.; Wyns, C. Fertility and infertility: Definition and epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef]
- Steuer, R. Review: On the analysis and interpretation of correlations in metabolomic data. Brief. Bioinform. 2006, 7, 151–158. [Google Scholar] [CrossRef]
- Vance, D.E.; Vance, J.E. Biochemistry of Lipids, Lipoproteins and Membranes, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2008. [Google Scholar] [CrossRef]
- Ferguson, E.M.; Leese, H.J. A potential role for triglyceride as an energy source during bovine oocyte maturation and early embryo development. Mol. Reprod. Dev. 2006, 73, 1195–1201. [Google Scholar] [CrossRef]
- Ferguson, E.M.; Leese, H.J. Triglyceride content of bovine oocytes and early embryos. J. Reprod. Fertil. 1999, 116, 373–378. [Google Scholar] [CrossRef]
- Dunning, K.R.; Russell, D.L.; Robker, R.L. Lipids and oocyte developmental competence: The role of fatty acids and beta-oxidation. Reproduction 2014, 148, 15–27. [Google Scholar] [CrossRef]
- Gautier, T.; Becker, S.; Drouineaud, V.; Menetrier, F.; Sagot, P.; Nofer, J.R.; von Otte, S.; Lagrost, L.; Masson, D.; Tietge, U.J. Human luteinized granulosa cells secrete apoB100-containing lipoproteins. J. Lipid Res. 2010, 51, 2245–2252. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Male Factor Infertility (n = 30) | Unexplained Infertility (n = 30) | p-Value |
|---|---|---|---|
| Age (years) | 32 ± 5.2 | 38 ± 5.2 | <0.001 |
| BMI (kg/m2) | 23.5 ± 3.6 | 27.2 ± 6.9 | 0.02 |
| Previous Pregnancies | 0.83 ± 1.1 | 0.7 ± 1.1 | 0.65 |
| Previous Deliveries | 0.53 ± 1.02 | 0.13 ± 0.34 | 0.05 |
| Previous Miscarriages | 0.27 ± 0.51 | 0.43 ± 0.88 | 0.38 |
| No. of previous IVF cycles | 2 ± 1.6 | 1.9 ± 1.1 | 0.78 |
| No. of retrieved oocytes | 11.83 ± 8.39 | 6.5 ± 5.7 | 0.006 |
| Maximal serum estradiol (pM) | 7434 ± 4109 | 4828 ± 2989 | 0.009 |
| Sperm Parameters: | |||
| Volume | 2.8 ± 1.3 | 2.8 ± 0.9 | 0.94 |
| Concentration | 6.4 ± 9.9 | 53.2 ± 31.1 | <0.01 |
| Motility | 16.6 ± 16.1 | 32.8 ± 19.7 | <0.01 |
| Morphology | 3.2 ± 4.9 | 4.7 ± 3.6 | 0.56 |
| Putative Identification | Molecular Ion/Precursor Ion | Empirical Formula (Molecular Ion) | log2 (FC) | q-Value * |
|---|---|---|---|---|
| TG(16:1/18:0/20:0) | 906.84 (M+NH4) | C57H108O6 | 2.04 | 0.00002 |
| TG(14:1/16:0/20:0) | 850.78 (M+NH4) | C53H100O6 | 2.04 | 0.00003 |
| TG(16:0/16:0/16:1) ** | 822.75 (M+NH4) | C51H96O6 | 1.96 | 0.00003 |
| TG(18:1/14:0/22:1) | 904.82 (M+NH4) | C57H106O6 | 1.47 | 0.00004 |
| TG(18:0/16:0/18:0) | 904.83 (M+ACN+H) | C55H106O6 | 1.66 | 0.00004 |
| TG(14:1/19:0/22:1) | 918.84 (M+NH4) | C58H108O6 | 1.85 | 0.00004 |
| TG(14:1/20:0/21:0) | 920.86 (M+NH4) | C58H110O6 | 2.56 | 0.00008 |
| TG(18:1/16:0/18:0) | 878.81 (M+NH4) | C55H104O6 | 2.11 | 0.00015 |
| TG(15:1/24:1/18:2) ** | 1008.89 (M+IsoProp+Na+H) | C60H108O6 | 1.53 | 0.00027 |
| TG(16:0/16:1/16:1) | 820.73 (M+NH4) | C51H94O6 | 1.49 | 0.00041 |
| LysoPC(18:2) | 520.61 (M+H) | C26H50NO7P | −2.16 | 0.00181 |
| TG(14:0/16:0/16:1) | 794.72 (M+NH4) | C49H92O6 | 1.76 | 0.00185 |
| PC(P-16:0/20:2) | 1562.18 (2M+Na) | C44H84NO7P | −1.44 | 0.00241 |
| Unidentified | 959.13 | 1.71 | 0.00241 | |
| Unidentified | 1044.7055 | −2.14 | 0.01089 | |
| Unidentified | 1005.69 | −2.12 | 0.01388 | |
| TG(16:1/18:1/18:1) | 876.80(M+NH4) | C53H102O6 | 2.18 | 0.01467 |
| Unidentified | 524.907 | −2.30 | 0.01625 | |
| Unidentified | 783.9095 | −2.02 | 0.01625 | |
| Unidentified | 759.4597 | −2.14 | 0.01625 | |
| Unidentified | 1036.93 | 1.84 | 0.01625 | |
| LysoPC(18:0) | 524.371 (M+H) | C26H54NO7P | −2.24 | 0.01774 |
| Lyso PC(18:1) | 1043.702 (2M+H) | C26H52NO7P | −2.20 | 0.02053 |
| LysoPC(18:1) | 522.35 (M+H) | C26H52NO7P | −2.24 | 0.02191 |
| Unidentified | 783.4730 | −2.10 | 0.02191 | |
| Unidentified | 1056.89 | 1.28 | 0.02226 | |
| TG(14:0/18:3/16:0) | 818.72 (M+NH4) | C51H92O6 | 0.82 | 0.02600 |
| TG(18:0/24:0/20:4) | 1058.9 (M+ACN+Na) | C65H118O6 | 1.19 | 0.02882 |
| TG(20:0/20:3/22:0) | 1060.92 (M+ACN+Na) | C65H120O6 | 1.45 | 0.03621 |
| TG(20:3/20:1/22:0) ** | 1058.90 (M+ACN+Na) | C65H118O6 | 1.13 | 0.04079 |
| SM(d18:1/16:0) ** | 1406.141 (2M+H) | C39H79N2O6P | −1.31 | 0.05886 |
| TG(20:0/22:3/22:2) | 1084.92 (M+ACN+Na) | C67H120O6 | 1.16 | 0.08095 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batushansky, A.; Zacharia, A.; Shehadeh, A.; Bruck-Haimson, R.; Saidemberg, D.; Kogan, N.M.; Thomas Mannully, C.; Herzberg, S.; Ben-Meir, A.; Moussaieff, A. A Shift in Glycerolipid Metabolism Defines the Follicular Fluid of IVF Patients with Unexplained Infertility. Biomolecules 2020, 10, 1135. https://doi.org/10.3390/biom10081135
Batushansky A, Zacharia A, Shehadeh A, Bruck-Haimson R, Saidemberg D, Kogan NM, Thomas Mannully C, Herzberg S, Ben-Meir A, Moussaieff A. A Shift in Glycerolipid Metabolism Defines the Follicular Fluid of IVF Patients with Unexplained Infertility. Biomolecules. 2020; 10(8):1135. https://doi.org/10.3390/biom10081135
Chicago/Turabian StyleBatushansky, Albert, Anish Zacharia, Alaa Shehadeh, Reut Bruck-Haimson, Daniel Saidemberg, Natalya M. Kogan, Chanchal Thomas Mannully, Shmuel Herzberg, Assaf Ben-Meir, and Arieh Moussaieff. 2020. "A Shift in Glycerolipid Metabolism Defines the Follicular Fluid of IVF Patients with Unexplained Infertility" Biomolecules 10, no. 8: 1135. https://doi.org/10.3390/biom10081135
APA StyleBatushansky, A., Zacharia, A., Shehadeh, A., Bruck-Haimson, R., Saidemberg, D., Kogan, N. M., Thomas Mannully, C., Herzberg, S., Ben-Meir, A., & Moussaieff, A. (2020). A Shift in Glycerolipid Metabolism Defines the Follicular Fluid of IVF Patients with Unexplained Infertility. Biomolecules, 10(8), 1135. https://doi.org/10.3390/biom10081135