The Antimicrobial Cathelicidin CRAMP Augments Platelet Activation during Psoriasis in Mice

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Imiquimod-Induced “Psoriasis-Like” Skin Inflammation
2.2. Tail Bleeding Assay
2.3. Mouse Blood Collection and Platelet Preparation
2.4. Flow Cytometry-Based Assays
2.5. Enzyme-Linked Immunosorbent Assays (ELISA)
2.6. Statistical Analysis
3. Results
3.1. Characterisation of the IMQ Mouse Model of Psoriasis
3.2. Haemostasis Is Not Affected in IMQ-Treated Mice
3.3. IMQ Does Not Directly Affect Platelet Activation
3.4. mCRAMP Is Elevated in the Skin and Plasma of IMQ-Treated Mice
3.5. The Level of Soluble P-Selectin Is Elevated in Psoriatic Mouse Plasma
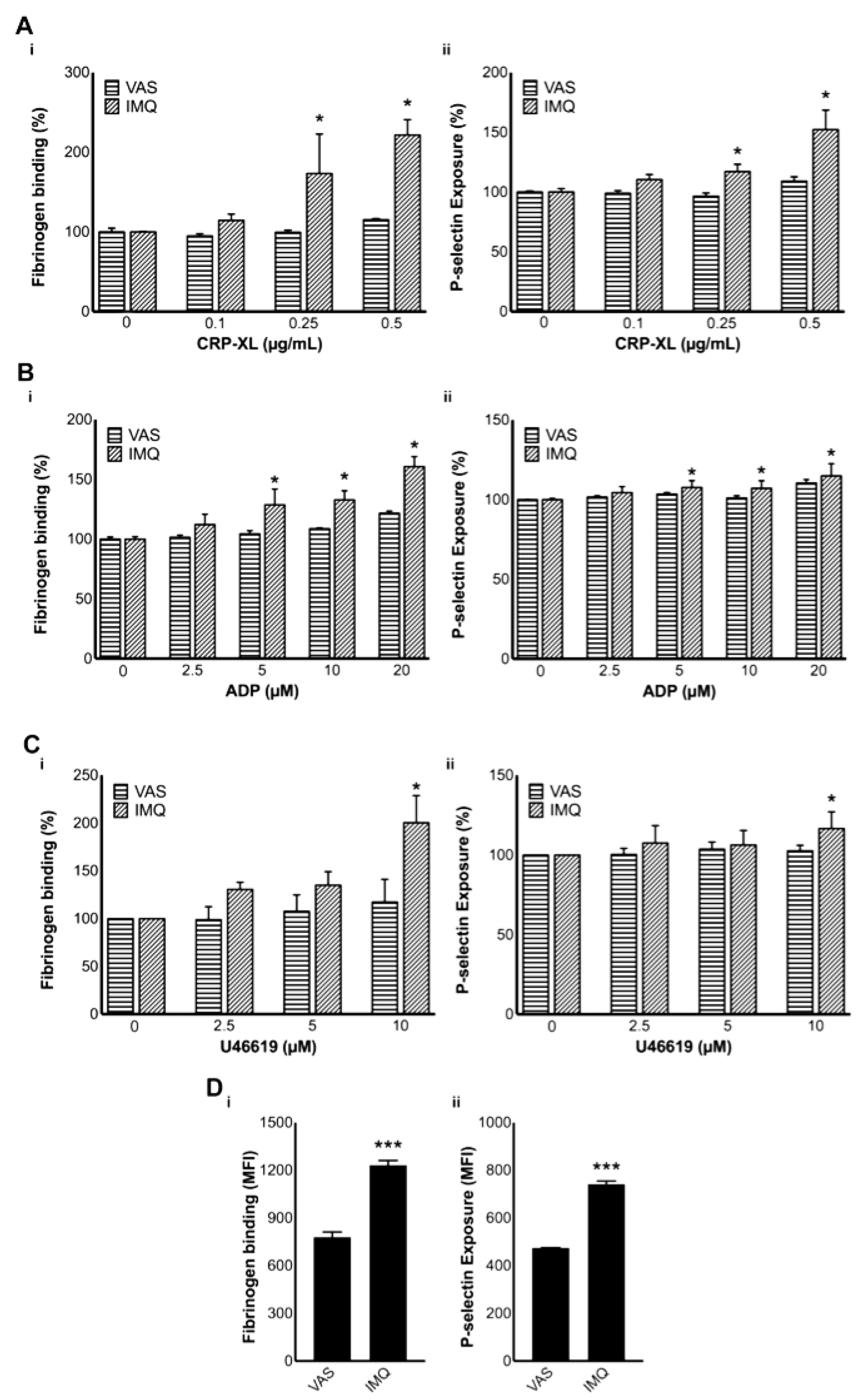
3.6. Platelet Activation Is Increased in IMQ-Treated Mice
3.7. Psoriatic Mouse Plasma Activates Healthy Mouse Platelets
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Boehncke, W.H.; Schon, M.P. Psoriasis. Lancet 2015, 386, 983–994. [Google Scholar] [CrossRef]
- Ashcroft, D.M.; Wan, P.A.L.; Williams, H.C.; Griffiths, C.E. Clinical measures of disease severity and outcome in psoriasis: A critical appraisal of their quality. Br. J. Dermatol. 1999, 141, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Marcinkiewicz, M.; Majewski, S. The role of antimicrobial peptides in chronic inflammatory skin diseases. Postepy Dermatol. Alergol. 2016, 33, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009, 30, 131–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morizane, S.; Gallo, R.L. Antimicrobial peptides in the pathogenesis of psoriasis. J. Dermatol. 2012, 39, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Ong, P.Y.; Ohtake, T.; Brandt, C.; Strickland, I.; Boguniewicz, M.; Ganz, T.; Gallo, R.L.; Leung, D.Y. Endogenous antimicrobial peptides and skin infections in atopic dermatitis. N. Engl. J. Med. 2002, 347, 1151–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verjans, E.T.; Zels, S.; Luyten, W.; Landuyt, B.; Schoofs, L. Molecular mechanisms of LL-37-induced receptor activation: An overview. Peptides 2016, 85, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Salamah, M.F.; Ravishankar, D.; Kodji, X.; Moraes, L.A.; Williams, H.F.; Vallance, T.M.; Albadawi, D.A.; Vaiyapuri, R.; Watson, K.; Gibbins, J.M.; et al. The endogenous antimicrobial cathelicidin LL37 induces platelet activation and augments thrombus formation. Blood Adv. 2018, 2, 2973–2985. [Google Scholar] [CrossRef] [PubMed]
- De, Y.; Chen, Q.; Schmidt, A.P.; Anderson, G.M.; Wang, J.M.; Wooters, J.; Oppenheim, J.J.; Chertov, O. LL-37, the neutrophil granule- and epithelial cell-derived cathelicidin, utilizes formyl peptide receptor-like 1 (FPRL1) as a receptor to chemoattract human peripheral blood neutrophils, monocytes, and T cells. J. Exp. Med. 2000, 192, 1069–1074. [Google Scholar]
- Doolin, T.; Amir, H.M.; Duong, L.; Rosenzweig, R.; Urban, L.A.; Bosch, M.; Pol, A.; Gross, S.P.; Siryaporn, A. Mammalian histones facilitate antimicrobial synergy by disrupting the bacterial proton gradient and chromosome organization. Nat. Commun. 2020, 11, 3888. [Google Scholar] [CrossRef]
- Bandurska, K.; Berdowska, A.; Barczyńska-Felusiak, R.; Krupa, P. Unique features of human cathelicidin LL-37. Biofactors 2015, 41, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Kahlenberg, J.M.; Kaplan, M.J. Little peptide, big effects: The role of LL-37 in inflammation and autoimmune disease. J. Immunol. 2013, 191, 4895–4901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, K.; Schauber, J.; Coda, A.; Lin, H.; Dorschner, R.A.; Schechter, N.M.; Bonnart, C.; Descargues, P.; Hovnanian, A.; Gallo, R.L. Kallikrein-mediated proteolysis regulates the antimicrobial effects of cathelicidins in skin. FASEB J. 2006, 20, 2068–2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusaka, S.; Nishida, A.; Takahashi, K.; Bamba, S.; Yasui, H.; Kawahara, M.; Inatomi, O.; Sugimoto, M.; Andoh, A. Expression of human cathelicidin peptide LL-37 in inflammatory bowel disease. Clin. Exp. Immunol. 2018, 191, 96–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulsen, F.; Pufe, T.; Conradi, L.; Varoga, D.; Tsokos, M.; Papendieck, J.; Petersen, W. Antimicrobial peptides are expressed and produced in healthy and inflamed human synovial membranes. J. Pathol. 2002, 198, 369–377. [Google Scholar] [CrossRef]
- Li, D.; Li, J.; Duan, Y.; Zhou, X. Expression of LL-37, human beta defensin-2, and CCR6 mRNA in patients with psoriasis vulgaris. J. Huazhong Univ. Sci. Technolog. Med. Sci. 2004, 24, 404–406. [Google Scholar]
- Leung, T.F.; Ching, K.W.; Kong, A.P.; Wong, G.W.; Chan, J.C.; Hon, K.L. Circulating LL-37 is a biomarker for eczema severity in children. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 518–522. [Google Scholar] [CrossRef]
- Döring, Y.; Drechsler, M.; Wantha, S.; Kemmerich, K.; Lievens, D.; Vijayan, S.; Gallo, R.L.; Weber, C.; Soehnlein, O. Lack of neutrophil-derived CRAMP reduces atherosclerosis in mice. Circ. Res. 2012, 110, 1052–1056. [Google Scholar] [CrossRef] [Green Version]
- Edfeldt, K.; Agerberth, B.; Rottenberg, M.E.; Gudmundsson, G.H.; Wang, X.B.; Mandal, K.; Xu, Q.; Yan, Z.Q. Involvement of the antimicrobial peptide LL-37 in human atherosclerosis. Arterioscler Thromb. Vasc. Biol. 2006, 26, 1551–1557. [Google Scholar] [CrossRef] [Green Version]
- Islam, D.; Bandholtz, L.; Nilsson, J.; Wigzell, H.; Christensson, B.; Agerberth, B.; Gudmundsson, G. Downregulation of bactericidal peptides in enteric infections: A novel immune escape mechanism with bacterial DNA as a potential regulator. Nat. Med. 2001, 7, 180–185. [Google Scholar] [CrossRef]
- An, L.L.; Ma, X.T.; Yang, Y.H.; Lin, Y.M.; Song, Y.H.; Wu, K.F. Marked reduction of LL-37/hCAP-18, an antimicrobial peptide, in patients with acute myeloid leukemia. Int. J. Hematol. 2005, 81, 45–47. [Google Scholar] [CrossRef] [PubMed]
- Heilborn, J.D.; Nilsson, M.F.; Kratz, G.; Weber, G.; Sørensen, O.; Borregaard, N.; Ståhle-Bäckdahl, M. The cathelicidin anti-microbial peptide LL-37 is involved in re-epithelialization of human skin wounds and is lacking in chronic ulcer epithelium. J. Investig. Dermatol. 2003, 120, 379–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pestonjamasp, V.K.; Huttner, K.H.; Gallo, R.L. Processing site and gene structure for the murine antimicrobial peptide CRAMP. Peptides 2001, 22, 1643–1650. [Google Scholar] [CrossRef]
- Hwang, Y.J.; Jung, H.J.; Kim, M.J.; Roh, N.K.; Jung, J.W.; Lee, Y.W.; Choe, Y.B.; Ahn, K.J. Serum levels of LL-37 and inflammatory cytokines in plaque and guttate psoriasis. Mediators Inflamm. 2014, 2014, 268257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pircher, J.; Czermak, T.; Ehrlich, A.; Eberle, C.; Gaitzsch, E.; Margraf, A.; Grommes, J.; Saha, P.; Titova, A.; Ishikawa-Ankerhold, H.; et al. Cathelicidins prime platelets to mediate arterial thrombosis and tissue inflammation. Nat. Commun. 2018, 9, 1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbins, J.M. Platelet adhesion signalling and the regulation of thrombus formation. J. Cell Sci. 2004, 117, 3415–3425. [Google Scholar] [CrossRef] [Green Version]
- Tamagawa-Mineoka, R.; Katoh, N.; Kishimoto, S. Platelet activation in patients with psoriasis: Increased plasma levels of platelet-derived microparticles and soluble P-selectin. J. Am. Acad. Dermatol. 2010, 62, 621–626. [Google Scholar] [CrossRef]
- Middleton, E.; Rondina, M.T. Platelets in infectious disease. Hematol. Am. Soc. Hematol. Educ. Program 2016, 2016, 256–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodji, X.; Arkless, K.L.; Kee, Z.; Cleary, S.J.; Aubdool, A.A.; Evans, E.; Caton, P.; Pitchford, S.C.; Brain, S.D. Sensory nerves mediate spontaneous behaviors in addition to inflammation in a murine model of psoriasis. FASEB J 2019, 33, 1578–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemény, Á.; Kodji, X.; Horváth, S.; Komlódi, R.; Szőke, É.; Sándor, Z.; Perkecz, A.; Gyömörei, C.; Sétáló, G.; Kelemen, B.; et al. TRPA1 Acts in a Protective Manner in Imiquimod-Induced Psoriasiform Dermatitis in Mice. J. Investig. Dermatol. 2018, 138, 1774–1784. [Google Scholar] [CrossRef] [Green Version]
- Vaiyapuri, S.; Sage, T.; Rana, R.H.; Schenk, M.P.; Ali, M.S.; Unsworth, A.J.; Jones, C.I.; Stainer, A.R.; Kriek, N.; Moraes, L.A.; et al. EphB2 regulates contact-dependent and contact-independent signaling to control platelet function. Blood 2015, 125, 720–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravishankar, D.; Salamah, M.; Attina, A.; Pothi, R.; Vallance, T.M.; Javed, M.; Williams, H.F.; Alzahrani, E.M.S.; Kabova, E.; Vaiyapuri, R.; et al. Ruthenium-conjugated chrysin analogues modulate platelet activity, thrombus formation and haemostasis with enhanced efficacy. Sci. Rep. 2017, 7, 5738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufton, N.; Hannon, R.; Brancaleone, V.; Dalli, J.; Patel, H.B.; Gray, M.; D’Acquisto, F.; Buckingham, J.C.; Perretti, M.; Flower, R.J. Anti-inflammatory role of the murine formyl-peptide receptor 2: Ligand-specific effects on leukocyte responses and experimental inflammation. J. Immunol. 2010, 184, 2611–2619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schön, M.P.; Schön, M.; Klotz, K.N. The small antitumoral immune response modifier imiquimod interacts with adenosine receptor signaling in a TLR7- and TLR8-independent fashion. J. Investig. Dermatol. 2006, 126, 1338–1347. [Google Scholar] [CrossRef] [Green Version]
- Nerurkar, L.; McColl, A.; Graham, G.; Cavanagh, J. The Systemic Response to Topical Aldara Treatment is Mediated Through Direct TLR7 Stimulation as Imiquimod Enters the Circulation. Sci. Rep. 2017, 7, 16570. [Google Scholar] [CrossRef]
- Puig, L. PASI90 response: The new standard in therapeutic efficacy for psoriasis. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 645–648. [Google Scholar] [CrossRef]
- Feldman, S.R.; Krueger, G.G. Psoriasis assessment tools in clinical trials. Ann. Rheum. Dis. 2005, 64, 69–73. [Google Scholar] [CrossRef]
- Fuentes-Duculan, J.; Bonifacio, K.M.; Hawkes, J.E.; Kunjravia, N.; Cueto, I.; Li, X.; Gonzalez, J.; Garcet, S.; Krueger, J.G. Autoantigens ADAMTSL5 and LL37 are significantly upregulated in active Psoriasis and localized with keratinocytes, dendritic cells and other leukocytes. Exp. Dermatol. 2017, 26, 1075–1082. [Google Scholar] [CrossRef]
- Tamagawa-Mineoka, R.; Katoh, N.; Ueda, E.; Masuda, K.; Kishimoto, S. Elevated platelet activation in patients with atopic dermatitis and psoriasis: Increased plasma levels of beta-thromboglobulin and platelet factor 4. Allergol. Int. 2008, 57, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Garbaraviciene, J.; Diehl, S.; Varwig, D.; Bylaite, M.; Ackermann, H.; Ludwig, R.J.; Boehncke, W.H. Platelet P-selectin reflects a state of cutaneous inflammation: Possible application to monitor treatment efficacy in psoriasis. Exp. Dermatol. 2010, 19, 736–741. [Google Scholar] [CrossRef]
- Chandrashekar, L.; Rajappa, M.; Revathy, G.; Sundar, I.; Munisamy, M.; Ananthanarayanan, P.H.; Thappa, D.M.; Basu, D. Is enhanced platelet activation the missing link leading to increased cardiovascular risk in psoriasis? Clin. Chim. Acta 2015, 446, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Semenov, A.V.; Romanov, Y.A.; Loktionova, S.A.; Tikhomirov, O.Y.; Khachikian, M.V.; Vasil’ev, S.A.; Mazurov, A.V. Production of soluble P-selectin by platelets and endothelial cells. Biochemistry 1999, 64, 1326–1335. [Google Scholar] [PubMed]
- Canpolat, F.; Akpinar, H.; Eskioğlu, F. Mean platelet volume in psoriasis and psoriatic arthritis. Clin. Rheumatol. 2010, 29, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Magen, E.; Mishal, J.; Zeldin, Y.; Feldman, V.; Kidon, M.; Schlesinger, M.; Sthoeger, Z. Increased mean platelet volume and C-reactive protein levels in patients with chronic urticaria with a positive autologous serum skin test. Am. J. Med. Sci. 2010, 339, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Willoughby, S.; Holmes, A.; Loscalzo, J. Platelets and cardiovascular disease. Eur. J. Cardiovasc. Nurs. 2002, 1, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Von Hundelshausen, P.; Weber, C. Platelets as immune cells: Bridging inflammation and cardiovascular disease. Circ. Res. 2007, 100, 27–40. [Google Scholar] [CrossRef]
- Ahlehoff, O.; Gislason, G.H.; Lindhardsen, J.; Charlot, M.G.; Jørgensen, C.H.; Olesen, J.B.; Bretler, D.M.; Skov, L.; Torp-Pedersen, C.; Hansen, P.R. Psoriasis carries an increased risk of venous thromboembolism: A Danish nationwide cohort study. PLoS ONE 2011, 6, e18125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lande, R.; Gregorio, J.; Facchinetti, V.; Chatterjee, B.; Wang, Y.H.; Homey, B.; Cao, W.; Wang, Y.H.; Su, B.; Nestle, F.O.; et al. Plasmacytoid dendritic cells sense self-DNA coupled with antimicrobial peptide. Nature 2007, 449, 564–569. [Google Scholar] [CrossRef]
- Wantha, S.; Alard, J.E.; Megens, R.T.; van der Does, A.M.; Döring, Y.; Drechsler, M.; Pham, C.T.; Wang, M.W.; Wang, J.M.; Gallo, R.L.; et al. Neutrophil-derived cathelicidin promotes adhesion of classical monocytes. Circ. Res. 2013, 112, 792–801. [Google Scholar] [CrossRef] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salamah, M.F.; Vallance, T.M.; Kodji, X.; Ravishankar, D.; Williams, H.F.; Brain, S.D.; Vaiyapuri, S. The Antimicrobial Cathelicidin CRAMP Augments Platelet Activation during Psoriasis in Mice. Biomolecules 2020, 10, 1267. https://doi.org/10.3390/biom10091267
Salamah MF, Vallance TM, Kodji X, Ravishankar D, Williams HF, Brain SD, Vaiyapuri S. The Antimicrobial Cathelicidin CRAMP Augments Platelet Activation during Psoriasis in Mice. Biomolecules. 2020; 10(9):1267. https://doi.org/10.3390/biom10091267
Chicago/Turabian StyleSalamah, Maryam F., Thomas M. Vallance, Xenia Kodji, Divyashree Ravishankar, Harry F. Williams, Susan D. Brain, and Sakthivel Vaiyapuri. 2020. "The Antimicrobial Cathelicidin CRAMP Augments Platelet Activation during Psoriasis in Mice" Biomolecules 10, no. 9: 1267. https://doi.org/10.3390/biom10091267