
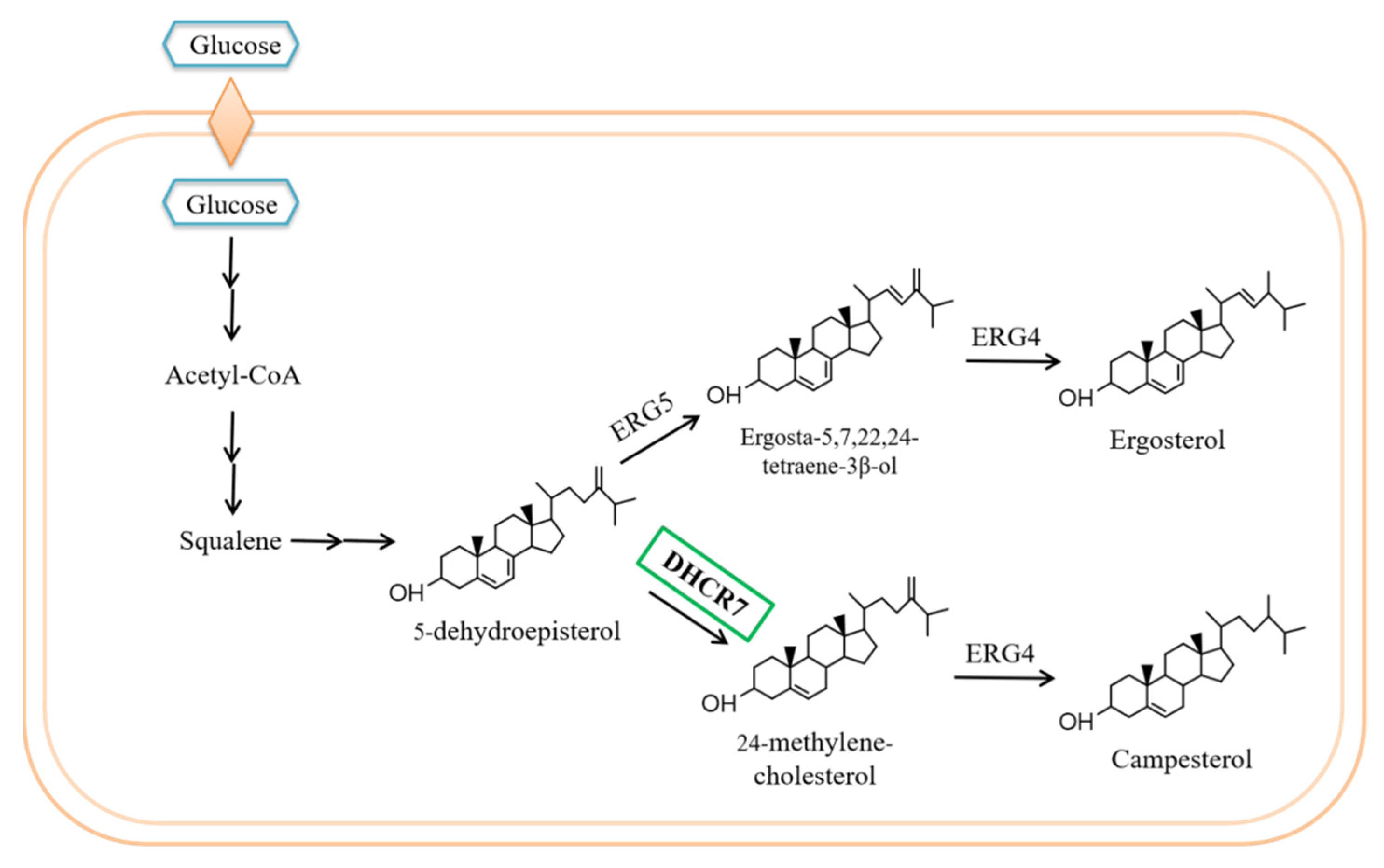
Engineering of Saccharomyces cerevisiae for 24-Methylene-Cholesterol Production


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Yang, J.; Li, C.; Zhang, Y. Engineering of Saccharomyces cerevisiae for 24-Methylene-Cholesterol Production. Biomolecules 2021, 11, 1710. https://doi.org/10.3390/biom11111710
Yang J, Li C, Zhang Y. Engineering of Saccharomyces cerevisiae for 24-Methylene-Cholesterol Production. Biomolecules. 2021; 11(11):1710. https://doi.org/10.3390/biom11111710
Chicago/Turabian StyleYang, Jiao, Changfu Li, and Yansheng Zhang. 2021. "Engineering of Saccharomyces cerevisiae for 24-Methylene-Cholesterol Production" Biomolecules 11, no. 11: 1710. https://doi.org/10.3390/biom11111710
APA StyleYang, J., Li, C., & Zhang, Y. (2021). Engineering of Saccharomyces cerevisiae for 24-Methylene-Cholesterol Production. Biomolecules, 11(11), 1710. https://doi.org/10.3390/biom11111710