
Characterization of miR-34a-Induced Epithelial–Mesenchymal Transition in Non-Small Lung Cancer Cells Focusing on p53


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture
2.2. Transfection with the miR-34a Mimic or siRNA for p53
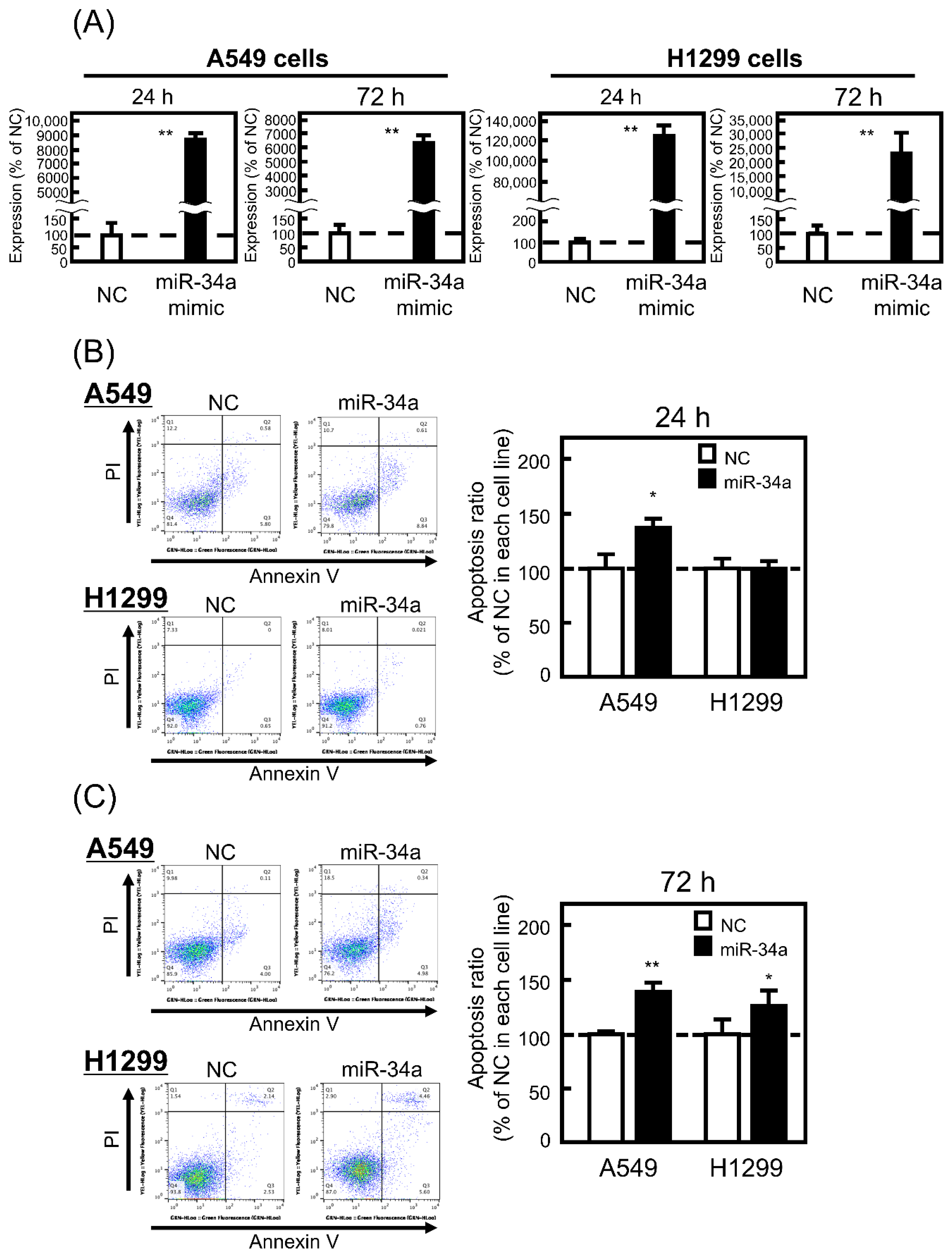
2.3. Apoptosis Assay
2.4. Scratch-Wound Assay
2.5. Detection of mRNA Expression Using Real-Time PCR Analysis
2.6. Detection of miRNA Expression Using Real-Time PCR Analysis
2.7. Detection of Protein Expression Using Western Blotting Analysis
2.8. Statistical Analysis
3. Results
3.1. Effect of miR-34a Overexpression on the Apoptosis Ratio of A549 and H1299 Cells
3.2. Effect of miR-34a Overexpression on EMT-Related Phenotypes in A549 and H1299 Cells
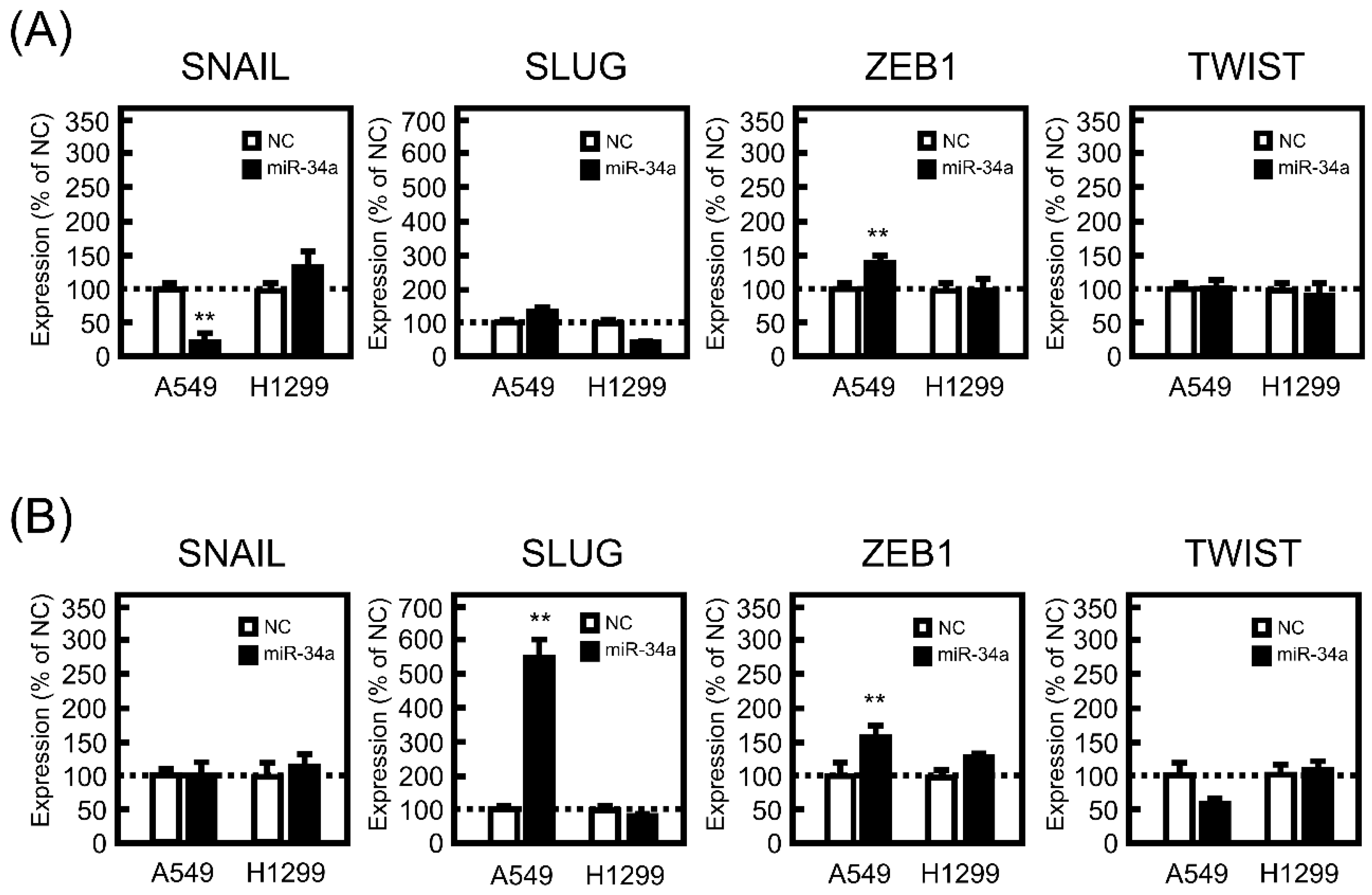
3.3. Effect of miR-34a Overexpression on the mRNA Expression Levels of EMT-Related Transcriptional Factors in A549 and H1299 Cells
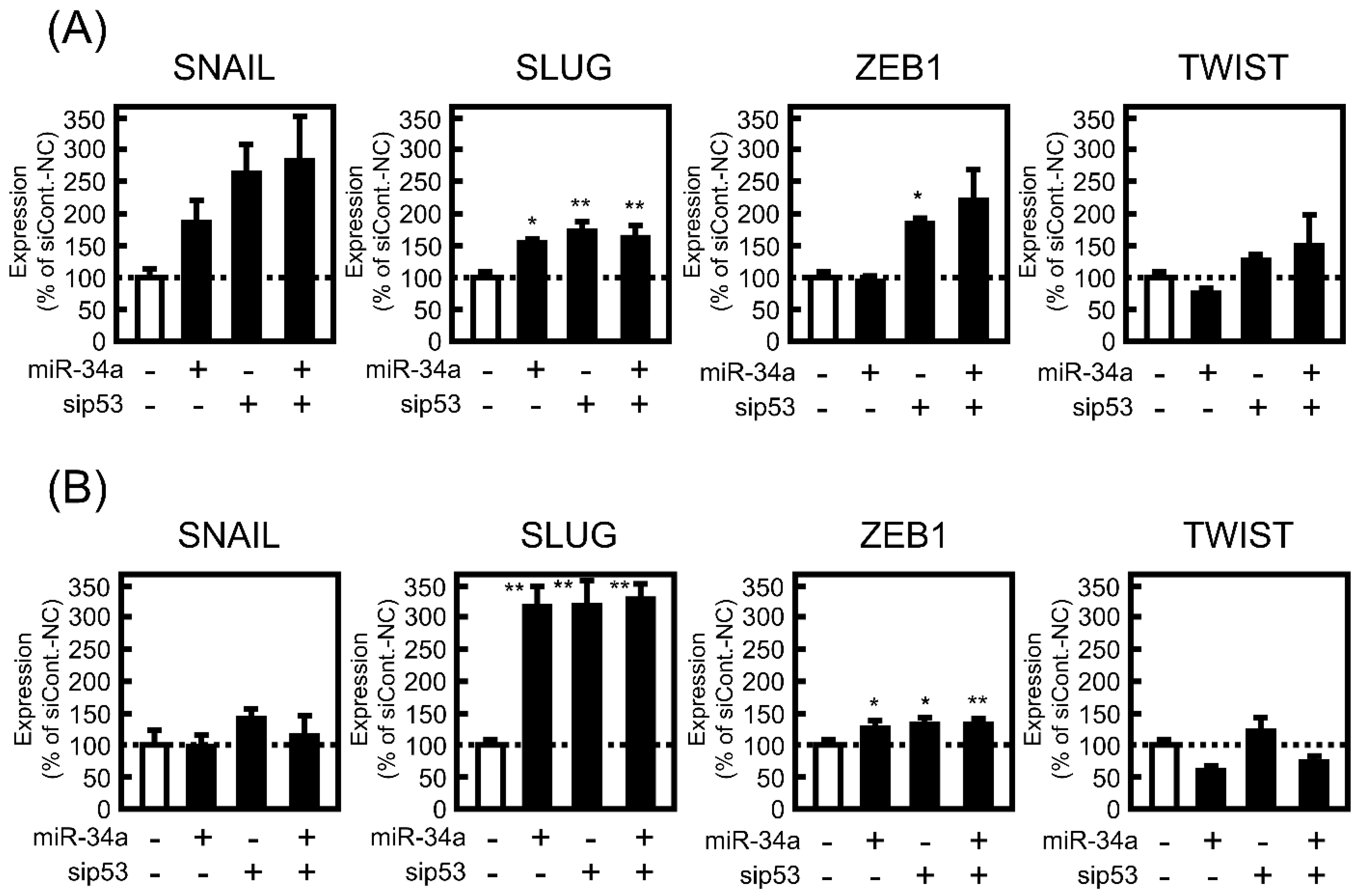
3.4. Effect of miR-34a Overexpression on EMT-Related Phenotypes under p53-Knockdown in A549 Cells
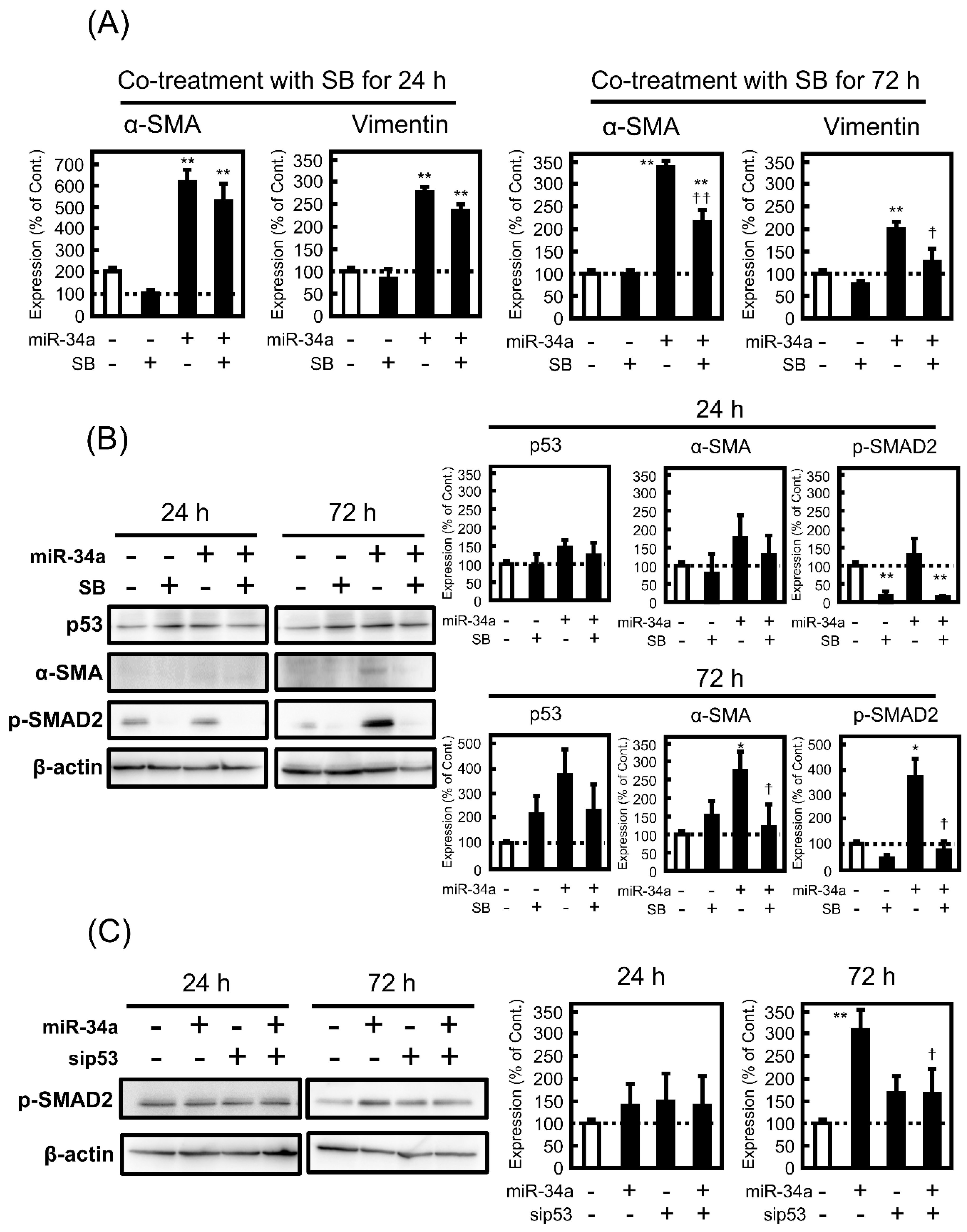
3.5. Effect of SB431542 (SB) on miR-34a-Induced EMT
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| epithelial–mesenchymal transition | EMT |
| microRNA | miRNA |
| non-small cell lung cancer | NSCLC |
References
- Takamizawa, J.; Konishi, H.; Yanagisawa, K.; Tomida, S.; Osada, H.; Endoh, H.; Harano, T.; Yatabe, Y.; Nagino, M.; Nimura, Y.; et al. Reduced Expression of the let-7 MicroRNAs in Human Lung Cancers in Association with Shortened Postoperative Survival. Cancer Res. 2004, 64, 3753–3756. [Google Scholar] [CrossRef] [Green Version]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Feng, Y.-L.; Liu, T.; Wang, J.-J.; Yu, J. MicroRNAs in organ fibrosis: From molecular mechanisms to potential therapeutic targets. Pathol.-Res. Pr. 2021, 225, 153588. [Google Scholar] [CrossRef]
- Hong, D.S.; Kang, Y.-K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.-L.; Kim, T.-Y.; et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- Bader, A.G. miR-34—A microRNA replacement therapy is headed to the clinic. Front. Genet. 2012, 3, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misso, G.; Di Martino, M.T.; De Rosa, G.; Farooqi, A.A.; Lombardi, A.; Campani, V.; Zarone, M.R.; Gullà, A.; Tagliaferri, P.; Tassone, P.; et al. Mir-34: A New Weapon Against Cancer? Mol. Ther.-Nucleic Acids 2014, 3, e195. [Google Scholar] [CrossRef]
- Hay, E.D. An Overview of Epithelio-Mesenchymal Transformation. Cells Tissues Organs 1995, 154, 8–20. [Google Scholar] [CrossRef]
- Dongre, A.; Weinberg, R.A. New insights into the mechanisms of epithelial–mesenchymal transition and implications for cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef]
- Takano, M.; Nekomoto, C.; Kawami, M.; Yumoto, R. Role of miR-34a in TGF-β1- and Drug-Induced Epithelial-Mesenchymal Transition in Alveolar Type II Epithelial Cells. J. Pharm. Sci. 2017, 106, 2868–2872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, A.; Kawami, M.; Konaka, T.; Takenaka, S.; Yumoto, R.; Takano, M. Anticancer Drug-Induced Epithelial-Mesenchymal Transition via p53/miR-34a axis in A549/ABCA3 Cells. J. Pharm. Pharm. Sci. 2019, 22, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Konaka, T.; Kawami, M.; Yamamoto, A.; Yumoto, R.; Takano, M. miR-484: A Possible Indicator of Drug-Induced Pulmonary Fibrosis. J. Pharm. Pharm. Sci. 2020, 23, 486–495. [Google Scholar] [CrossRef]
- Kawami, M.; Takenaka, S.; Kadekaru, Y.; Akai, M.; Konaka, T.; Yumoto, R.; Takano, M. Evaluation on epithelial-mesenchymal state and microRNAs focusing on isolated alveolar epithelial cells from bleomycin injured rat lung. Toxicology 2021, 461, 152903. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, J.; He, L.; Zhang, F. MicroRNA-34a Promotes EMT and Liver Fibrosis in Primary Biliary Cholangitis by Regulating TGF-β1/smad Pathway. J. Immunol. Res. 2021, 2021, 6890423. [Google Scholar] [CrossRef] [PubMed]
- Imani, S.; Wei, C.; Cheng, J.; Khan, A.; Fu, S.; Yang, L.; Tania, M.; Zhang, X.; Xiao, X.; Zhang, X.; et al. MicroRNA-34a targets epithelial to mesenchymal transition-inducing transcription factors (EMT-TFs) and inhibits breast cancer cell migration and invasion. Oncotarget 2017, 8, 21362–21379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, D.; Fu, J.; Chen, H.; Cheng, J.; Fu, J. Roles of MicroRNA-34a in Epithelial to Mesenchymal Transition, Competing Endogenous RNA Sponging and Its Therapeutic Potential. Int. J. Mol. Sci. 2019, 20, 861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; He, X.; Lim, L.P.; De Stanchina, E.; Xuan, Z.; Liang, Y.; Xue, W.; Zender, L.; Magnus, J.; Ridzon, D.; et al. A microRNA component of the p53 tumour suppressor network. Nat. Cell Biol. 2007, 447, 1130–1134. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.-C.; Wentzel, E.A.; Kent, O.; Ramachandran, K.; Mullendore, M.; Lee, K.H.; Feldmann, G.; Yamakuchi, M.; Ferlito, M.; Lowenstein, C.J.; et al. Transactivation of miR-34a by p53 Broadly Influences Gene Expression and Promotes Apoptosis. Mol. Cell 2007, 26, 745–752. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Antin, P.; Berx, G.; Blanpain, C.; Brabletz, T.; Bronner, M.; Campbell, K.; Cano, A.; Casanova, J.; Christofori, G.; et al. Guidelines and definitions for research on epithelial–mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2020, 21, 341–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, L.; Chen, J.; Ruan, L.; Tan, A.; Wang, P. miR-34a inhibits tumorigenesis of NSCLC via targeting SIRT6. Int. J. Clin. Exp. Pathol 2018, 11, 1135–1145. [Google Scholar] [PubMed]
- Ma, Z.-L.; Hou, P.-P.; Li, Y.; Wang, D.-T.; Yuan, T.-W.; Wei, J.-L.; Zhao, B.-T.; Lou, J.-T.; Zhao, X.-T.; Jin, Y.; et al. MicroRNA-34a inhibits the proliferation and promotes the apoptosis of non-small cell lung cancer H1299 cell line by targeting TGFβR2. Tumor Biol. 2014, 36, 2481–2490. [Google Scholar] [CrossRef]
- Rho, J.K.; Choi, Y.J.; Ryoo, B.-Y.; Na, I.I.; Yang, S.H.; Kim, C.H.; Lee, J.C. p53 Enhances Gefitinib-Induced Growth Inhibition and Apoptosis by Regulation of Fas in Non–Small Cell Lung Cancer. Cancer Res. 2007, 67, 1163–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojima, T.; Kawami, M.; Yumoto, R.; Takano, M. Differential mechanisms underlying methotrexate-induced cell death and epithelial-mesenchymal transition in A549 cells. Toxicol. Res. 2021, 37, 293–300. [Google Scholar] [CrossRef]
- Aleksakhina, S.; Kashyap, A.; Imyanitov, E.N. Mechanisms of acquired tumor drug resistance. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2019, 1872, 188310. [Google Scholar] [CrossRef] [PubMed]
- Brozovic, A. The relationship between platinum drug resistance and epithelial–mesenchymal transition. Arch. Toxicol. 2016, 91, 605–619. [Google Scholar] [CrossRef]
- Anggorowati, N.; Kurniasari, C.R.; Damayanti, K.; Cahyanti, T.; Widodo, I.; Ghozali, A.; Romi, M.M.; Sari, D.C.R.; Arfian, N. Histochemical and Immunohistochemical Study of α-SMA, Collagen, and PCNA in Epithelial Ovarian Neoplasm. Asian Pac. J. Cancer Prev. 2017, 18, 667–671. [Google Scholar] [CrossRef]
- Lee, H.W.; Park, Y.M.; Lee, S.J.; Cho, H.J.; Kim, D.-H.; Lee, J.-I.; Kang, M.-S.; Seol, H.J.; Shim, Y.M.; Nam, D.-H.; et al. Alpha-Smooth Muscle Actin (ACTA2) Is Required for Metastatic Potential of Human Lung Adenocarcinoma. Clin. Cancer Res. 2013, 19, 5879–5889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comer, K.A.; Dennis, P.A.; Armstrong, L.; Catino, J.J.; Kastan, M.B.; Kumar, C.C. Human smooth muscle α-actin gene is a transcriptional target of the p53 tumor suppressor protein. Oncogene 1998, 16, 1299–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.-H.; Li, C.-H.; Cheng, Y.-W.; Lee, C.-C.; Liao, P.-L.; Lin, C.-H.; Huang, S.-H.; Kang, J.-J. The inhibition of lung cancer cell migration by AhR-regulated autophagy. Sci. Rep. 2017, 7, srep41927. [Google Scholar] [CrossRef]
- Iida, K.; Sakai, R.; Yokoyama, S.; Kobayashi, N.; Togo, S.; Yoshikawa, H.Y.; Rawangkan, A.; Namiki, K.; Suganuma, M. Cell softening in malignant progression of human lung cancer cells by activation of receptor tyrosine kinase AXL. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-H.; Kim, D.-Y.; Jing, F.; Kim, H.; Yun, C.-O.; Han, D.-J.; Choi, E.Y. Del-1 overexpression potentiates lung cancer cell proliferation and invasion. Biochem. Biophys. Res. Commun. 2015, 468, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-P.; Wang, W.-L.; Chang, Y.-L.; Wu, C.-T.; Chao, Y.-C.; Kao, S.-H.; Yuan, A.; Lin, C.-W.; Yang, S.-C.; Chan, W.-K.; et al. p53 controls cancer cell invasion by inducing the MDM2-mediated degradation of Slug. Nat. Cell Biol. 2009, 11, 694–704. [Google Scholar] [CrossRef]
- Jung, C.-H.; Kim, J.; Park, J.K.; Hwang, S.-G.; Moon, S.-K.; Kim, W.-J.; Um, H.-D. Mdm2 increases cellular invasiveness by binding to and stabilizing the Slug mRNA. Cancer Lett. 2013, 335, 270–277. [Google Scholar] [CrossRef]
- Colak, S.; Ten Dijke, P. Targeting TGF-beta Signaling in Cancer. Trends Cancer 2017, 3, 56–71. [Google Scholar] [CrossRef]
- Kim, K.K.; Sheppard, D.; Chapman, H.A. TGF-β1 Signaling and Tissue Fibrosis. Cold Spring Harb. Perspect. Biol. 2018, 10, a022293. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.I. MicroRNA Control of TGF-β Signaling. Int. J. Mol. Sci. 2018, 19, 1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooqi, A.A.; Tabassum, S.; Ahmad, A. MicroRNA-34a: A Versatile Regulator of Myriads of Targets in Different Cancers. Int. J. Mol. Sci. 2017, 18, 2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slabáková, E.; Culig, Z.; Remšík, J.; Souček, K. Alternative mechanisms of miR-34a regulation in cancer. Cell Death Dis. 2017, 8, e3100. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Accession Number | Sequence |
|---|---|---|
| α-SMA | NC_000010.11 | Foward: (5′)GCTGTTTCCCATCCATTGT(3′) |
| Reverse: (5′)TTTGCTCTGTGCTTCGTCAC(3′) | ||
| CDKN1A | NC_000006.12 | Foward: (5′)GTGGACCTGTCACTGTCTTG(3′) |
| Reverse: (5′)GGCGTTTGGAGTGGTAGAAA(3′) | ||
| GAPDH | NC_000012.12 | Foward: (5′)ACGGGAAGCTTGTCATCAAT(3′) |
| Reverse: (5′)TGGACTCCACGACGTACTCA(3′) | ||
| N-cadherin | NC_000018.10 | Foward: (5′)AATCAGTGGCGGAGATCCTA(3′) |
| Reverse: (5′)CCTTGGCTAATGGCACTTGA(3′) | ||
| SNAIL | NC_000020.11 | Foward: (5′)GAGTGGTTCTTCTGCGCTAC(3′) |
| Reverse: (5′)TAGGGCTGCTGGAAGGTAAA(3′) | ||
| SLUG | NC_000008.11 | Foward: (5′)TATTCGGACCCACACATTAC(3′) |
| Reverse: (5′)CAGATTTGACCTGTCTGCAA(3′) | ||
| TWIST | NC_000007.14 | Foward: (5′)AGCTATGTGGCTCACGAG(3′) |
| Reverse: (5′)TGTCCATTTTCTCCTTCTCTGG(3′) | ||
| Vimentin | NC_000010.11 | Foward: (5′)TCAGAGAGAGGAAGCCGAAA(3′) |
| Reverse: (5′)CAAAAAGGCAATCTCTTCTTGC(3′) | ||
| ZEB1 | NC_000010.11 | Foward: (5′)AGCTGCCAATAAGCAAACGA(3′) |
| Reverse: (5′)TTTGGGCGGTGTAGAATCAG(3′) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawami, M.; Takenaka, S.; Akai, M.; Yumoto, R.; Takano, M. Characterization of miR-34a-Induced Epithelial–Mesenchymal Transition in Non-Small Lung Cancer Cells Focusing on p53. Biomolecules 2021, 11, 1853. https://doi.org/10.3390/biom11121853
Kawami M, Takenaka S, Akai M, Yumoto R, Takano M. Characterization of miR-34a-Induced Epithelial–Mesenchymal Transition in Non-Small Lung Cancer Cells Focusing on p53. Biomolecules. 2021; 11(12):1853. https://doi.org/10.3390/biom11121853
Chicago/Turabian StyleKawami, Masashi, Shinnosuke Takenaka, Mizuki Akai, Ryoko Yumoto, and Mikihisa Takano. 2021. "Characterization of miR-34a-Induced Epithelial–Mesenchymal Transition in Non-Small Lung Cancer Cells Focusing on p53" Biomolecules 11, no. 12: 1853. https://doi.org/10.3390/biom11121853
APA StyleKawami, M., Takenaka, S., Akai, M., Yumoto, R., & Takano, M. (2021). Characterization of miR-34a-Induced Epithelial–Mesenchymal Transition in Non-Small Lung Cancer Cells Focusing on p53. Biomolecules, 11(12), 1853. https://doi.org/10.3390/biom11121853