
Iron Deficiency in Obesity and after Bariatric Surgery

 ,
,  ,
,  , ,
, ,  , ,
, ,  ,
,  and
and 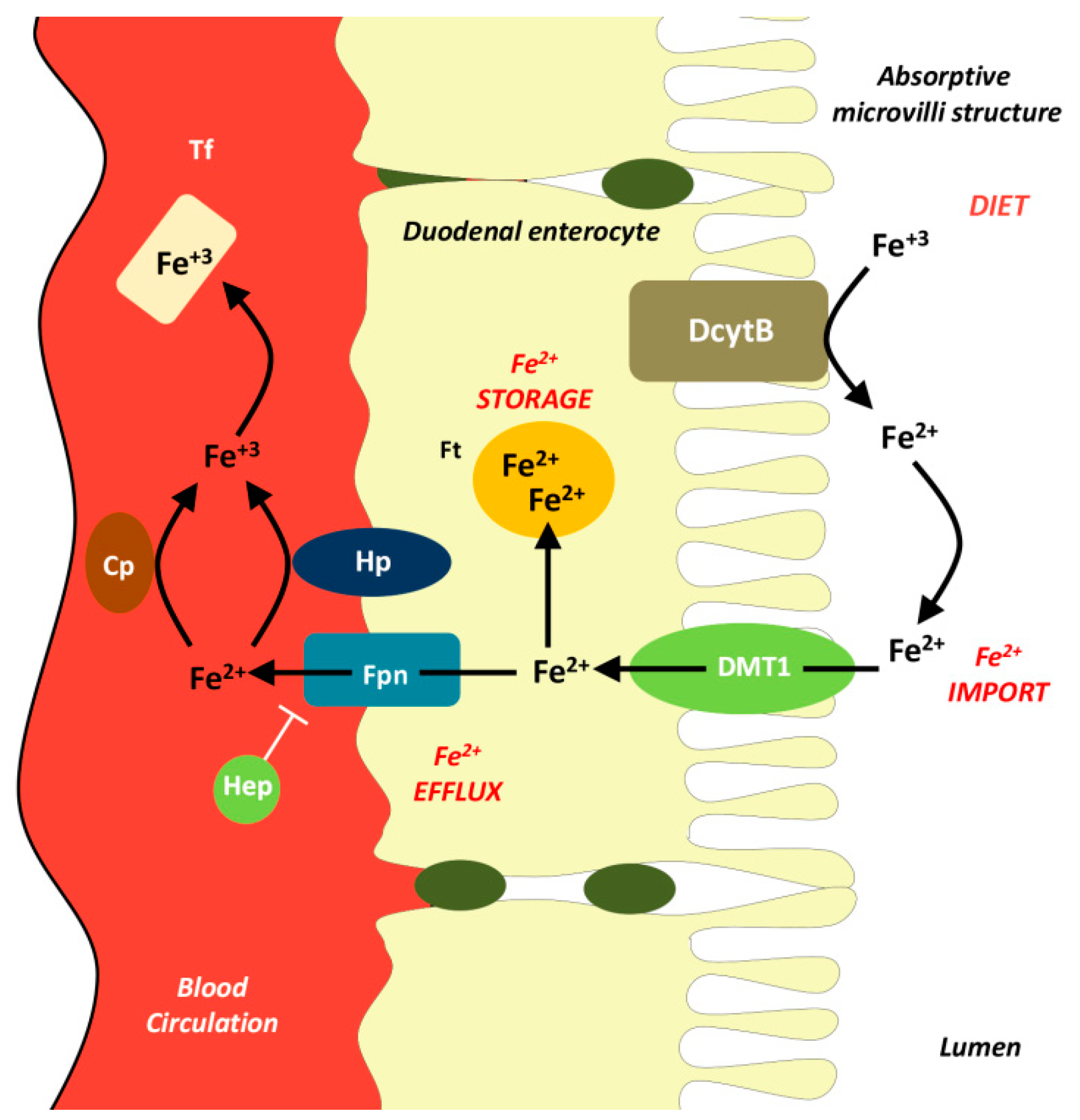{kind=link}
Abstract
:1. Introduction
- decrease in iron intake due to its low absorption
- poor tolerance to iron reach foods
- low adherence of patients to iron-containing drugs
- decreased hydrochloric acid secretion due to the presence of a shunt or resection of the greater curvature of the stomach
- decrease in the absorbing surface due to a duodenal shunt or damage to the intestinal villi.
2. Iron Metabolism
3. Biomarkers of Iron Status
4. Iron Deficiency and Anemia in Obesity
5. Iron Deficiency, Inflammation, and Obesity
6. Bariatric Surgery and Postoperative Iron Status
7. Iron Supplementation after Bariatric Surgery
8. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Li, Y.; Yang, W.; Dong, D.; Jiang, S.; Yang, Z.; Wang, Y. Effect of different sources and levels of iron in the diet of sows on iron status in neonatal pigs. Anim. Nutr. 2018, 4, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Beard, J.L. Why iron deficiency is important in infant development. J. Nutr. 2008, 138, 2534–2536. [Google Scholar] [CrossRef] [PubMed]
- Zoroddu, M.A.; Aaseth, J.; Crisponi, G.; Medici, S.; Peana, M.; Nurchi, V.M. The essential metals for humans: A brief overview. J. Inorg. Biochem. 2019, 195, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-L.; Ghosh, M.C.; Rouault, T.A. The physiological functions of iron regulatory proteins in iron homeostasis-an update. Front. Pharmacol. 2014, 5, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbaspour, N.; Hurrell, R.; Kelishadi, R. Review on iron and its importance for human health. J. Res. Med. Sci. 2014, 19, 164. [Google Scholar] [PubMed]
- Harigae, H. Iron metabolism and related diseases: An overview. Int. J. Hematol. 2018, 107, 5–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebastiani, G.; Pantopoulos, K. Disorders associated with systemic or local iron overload: From pathophysiology to clinical practice. Metallomics 2011, 3, 971–986. [Google Scholar] [CrossRef]
- Bjørklund, G.; Dadar, M.; Peana, M.; Rahaman, M.S.; Aaseth, J. Interactions between iron and manganese in neurotoxicity. Arch Toxicol. 2020, 94, 725–734. [Google Scholar] [CrossRef]
- Hanif, N.; Anwer, F. Chronic Iron Deficiency; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Nikonorov, A.A.; Skalnaya, M.G.; Tinkov, A.A.; Skalny, A.V. Mutual interaction between iron homeostasis and obesity pathogenesis. J. Trace Elem. Med. Biol. 2015, 30, 207–214. [Google Scholar] [CrossRef]
- Gonzalez-Dominguez, A.; Visiedo-Garcia, F.M.; Dominguez-Riscart, J.; Gonzalez-Dominguez, R.; Mateos, R.M.; Lechuga-Sancho, A.M. Iron Metabolism in Obesity and Metabolic Syndrome. Int. J. Mol. Sci. 2020, 21, 5529. [Google Scholar] [CrossRef] [PubMed]
- Lachowicz, J.I.; Nurchi, V.M.; Fanni, D.; Gerosa, C.; Peana, M.; Zoroddu, M.A. Nutritional iron deficiency: The role of oral iron supplementation. Curr. Med. Chem. 2014, 21, 3775–3784. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.; Emmanuel, J. Global concept of iron deficiency. In Iron Deficiency and Overload; Yehuda, S., Mostofsky, D., Eds.; Humana Press: Totowa, NJ, USA, 2009; pp. 299–312. [Google Scholar]
- Htet, M.K.; Fahmida, U.; Dillon, D.; Akib, A.; Utomo, B.; Thurnham, D.I. Is Iron Supplementation Influenced by Sub-Clinical Inflammation? A Randomized Controlled Trial Among Adolescent Schoolgirls in Myanmar. Nutrients 2019, 11, 918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, G.; Goodnough, L.T. Anemia of chronic disease. N. Engl. J. Med. 2005, 352, 1011–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, C.N.; Andrews, N.C. Anemia of inflammation: The hepcidin link. Curr. Opin. Hematol. 2005, 12, 107–111. [Google Scholar] [CrossRef]
- Verma, S.; Cherayil, B.J. Iron and inflammation—the gut reaction. Metallomics 2017, 9, 101–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwiczek, S.; Aigner, E.; Theurl, I.; Weiss, G. Cytokine-mediated regulation of iron transport in human monocytic cells. Blood 2003, 101, 4148–4154. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Hernandez, H.; Simental-Mendia, L.E.; Rodriguez-Ramirez, G.; Reyes-Romero, M.A. Obesity and inflammation: Epidemiology, risk factors, and markers of inflammation. Int. J. Endocrinol. 2013, 2013, 678159. [Google Scholar] [CrossRef] [Green Version]
- Ortíz Pérez, M.; Vázquez López, M.A.; Ibáñez Alcalde, M.; Galera Martínez, R.; Martín González, M.; Lendínez Molinos, F.; Bonillo Perales, A. Relationship between Obesity and Iron Deficiency in Healthy Adolescents. Child. Obes. 2020, 16, 440–447. [Google Scholar] [CrossRef]
- Aigner, E.; Feldman, A.; Datz, C. Obesity as an emerging risk factor for iron deficiency. Nutrients 2014, 6, 3587–3600. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, X.; Shen, Y.; Fang, X.; Wang, Y.; Wang, F. Obesity and iron deficiency: A quantitative meta-analysis. Obes. Rev. 2015, 16, 1081–1093. [Google Scholar] [CrossRef]
- Cepeda-Lopez, A.C.; Aeberli, I.; Zimmermann, M.B. Does Obesity Increase Risk for Iron Deficiency? A Review of the Literature and the Potential Mechanisms. Int. J. Vitam. Nutr. Res. 2010, 80, 263–270. [Google Scholar] [CrossRef]
- McClung, J.P.; Karl, J.P. Iron deficiency and obesity: The contribution of inflammation and diminished iron absorption. Nutr. Rev. 2009, 67, 100–104. [Google Scholar] [CrossRef]
- Yanoff, L.B.; Menzie, C.M.; Denkinger, B.; Sebring, N.G.; McHugh, T.; Remaley, A.T.; Yanovski, J.A. Inflammation and iron deficiency in the hypoferremia of obesity. Int. J. Obes. 2007, 31, 1412–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, I.C.; Tseng, S.-H.; Aulia, B.; Shih, C.-K.; Bai, C.-H.; Chang, J.-S. Can diet-induced weight loss improve iron homoeostasis in patients with obesity: A systematic review and meta-analysis. Obes. Rev. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zuo, D.; Xiao, X.; Yang, S.; Gao, Y.; Wang, G.; Ning, G. Effects of bariatric surgery in Chinese with obesity and type 2 diabetes mellitus: A 3-year follow-up. Medicine 2020, 99, e21673. [Google Scholar] [CrossRef] [PubMed]
- Steenackers, N.; Van der Schueren, B.; Mertens, A.; Lannoo, M.; Grauwet, T.; Augustijns, P.; Matthys, C. Iron deficiency after bariatric surgery: What is the real problem? Proc. Nutr. Soc. 2018, 77, 445–455. [Google Scholar] [CrossRef] [Green Version]
- Vazquez Prado, A.; Montalva Oron, E.M.; de Tursi Rispoli, L.C. Assessment of morbid-obesity-associated comorbidities progression after surgical treatment with the duodenal crossing technique. Nutr. Hosp. 2007, 22, 596–601. [Google Scholar]
- Ernst, B.; Thurnheer, M.; Schmid, S.M.; Schultes, B. Evidence for the necessity to systematically assess micronutrient status prior to bariatric surgery. Obes. Surg. 2009, 19, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Toh, S.Y.; Zarshenas, N.; Jorgensen, J. Prevalence of nutrient deficiencies in bariatric patients. Nutrition 2009, 25, 1150–1156. [Google Scholar] [CrossRef]
- Skroubis, G.; Sakellaropoulos, G.; Pouggouras, K.; Mead, N.; Nikiforidis, G.; Kalfarentzos, F. Comparison of Nutritional Deficiencies after Rouxen-Y Gastric Bypass and after Biliopancreatic Diversion with Roux-en-Y Gastric Bypass. Obes. Surg. 2002, 12, 551–558. [Google Scholar] [CrossRef]
- Coupaye, M.; Puchaux, K.; Bogard, C.; Msika, S.; Jouet, P.; Clerici, C.; Larger, E.; Ledoux, S. Nutritional consequences of adjustable gastric banding and gastric bypass: A 1-year prospective study. Obes. Surg. 2009, 19, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Salinas, A.; Salinas, H.M.; Santiago, E.; Garcia, W.; Ferro, Q.; Antor, M. Silastic ring vertical gastric bypass: Cohort study with 83% rate of 5-year follow-up. Surg. Obes. Relat. Dis. 2009, 5, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Kitagishi, H.; Kano, K. Supramolecular Iron Porphyrin/Cyclodextrin Dimer Complex that Mimics the Functions of Hemoglobin and Methemoglobin. Angew. Chem. Int. Ed. 2013, 52, 6894–6897. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C. Essential functions of iron-requiring proteins in DNA replication, repair and cell cycle control. Protein Cell 2014, 5, 750–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjørklund, G.; Aaseth, J.; Skalny, A.V.; Suliburska, J.; Skalnaya, M.G.; Nikonorov, A.A.; Tinkov, A.A. Interactions of iron with manganese, zinc, chromium, and selenium as related to prophylaxis and treatment of iron deficiency. J. Trace Elem. Med. Biol. 2017, 41, 41–53. [Google Scholar] [CrossRef]
- Neidlein, S.; Wirth, R.; Pourhassan, M. Iron deficiency, fatigue and muscle strength and function in older hospitalized patients. Eur. J. Clin. Nutr. 2020. [Google Scholar] [CrossRef]
- Grubić Kezele, T.; Ćurko-Cofek, B. Age-Related Changes and Sex-Related Differences in Brain Iron Metabolism. Nutrients 2020, 12, 2601. [Google Scholar] [CrossRef]
- Listabarth, S.; König, D.; Vyssoki, B.; Hametner, S. Does thiamine protect the brain from iron overload and alcohol-related dementia? Alzheimers Dement 2020, 16. [Google Scholar] [CrossRef]
- Tippairote, T.; Bjørklund, G.; Peana, M.; Roytrakul, S. The Proteomics Study of Compounded HFE/TF/TfR2/HJV Genetic Variations in a Thai Family with Iron Overload, Chronic Anemia, and Motor Neuron Disorder. J. Mol. Neurosci. 2020. [Google Scholar] [CrossRef]
- Sheikh, N.; Farrukh, A.; Abbas, A. Hepatic iron regulatory gene expression influenced due to disturbed essential trace elements level. J. Anim. Plant Sci. 2017, 27, 1024–1030. [Google Scholar]
- Peirano, P.D.; Algarín, C.R.; Chamorro, R.; Garrido, M.I.; Lozoff, B. Sleep and neurofunctions throughout child development: Lasting effects of early iron deficiency. J. Pediatric Gastroenterol. Nutr. 2009, 48, S8. [Google Scholar] [CrossRef] [Green Version]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abboud, S.; Haile, D.J. A novel mammalian iron-regulated protein involved in intracellular iron metabolism. J. Biol. Chem. 2000, 275, 19906–19912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, D.F. The Regulation of Iron Absorption and Homeostasis. Clin. Biochem. Rev. 2016, 37, 51–62. [Google Scholar] [PubMed]
- Mackenzie, B.; Garrick, M.D. Iron Imports. II. Iron uptake at the apical membrane in the intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 289, G981–G986. [Google Scholar] [CrossRef] [Green Version]
- Latunde-Dada, G.O.; Simpson, R.J.; McKie, A.T. Duodenal cytochrome B expression stimulates iron uptake by human intestinal epithelial cells. J. Nutr. 2008, 138, 991–995. [Google Scholar] [CrossRef]
- Goodnough, L.T.; Nemeth, E.; Ganz, T. Detection, evaluation, and management of iron-restricted erythropoiesis. Blood 2010, 116, 4754–4761. [Google Scholar] [CrossRef]
- Little, H.C.; Rodriguez, S.; Lei, X.; Tan, S.Y.; Stewart, A.N.; Sahagun, A.; Sarver, D.C.; Wong, G.W. Myonectin deletion promotes adipose fat storage and reduces liver steatosis. FASEB J. 2019, 33, 8666–8687. [Google Scholar] [CrossRef]
- Datz, C.; Felder, T.K.; Niederseer, D.; Aigner, E. Iron homeostasis in the metabolic syndrome. Eur. J. Clin. Investig. 2013, 43, 215–224. [Google Scholar] [CrossRef]
- Beard, J.L.; Murray-Kolb, L.E.; Rosales, F.J.; Solomons, N.W.; Angelilli, M.L. Interpretation of serum ferritin concentrations as indicators of total-body iron stores in survey populations: The role of biomarkers for the acute phase response. Am. J. Clin. Nutr. 2006, 84, 1498–1505. [Google Scholar] [CrossRef] [Green Version]
- Kuriyama-Matsumura, K.; Sato, H.; Yamaguchi, M.; Bannai, S. Regulation of ferritin synthesis and iron regulatory protein 1 by oxygen in mouse peritoneal macrophages. Biochem. Biophys. Res. Commun. 1998, 249, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.C.; Schaffer, K.J.; Hazegh-Azam, M.; Zhou, C.Y.; Tran, T.N.; Nagel, G.M. Serum ferritin: Does it differ from tissue ferritin? J. Gastroenterol. Hepatol. 1996, 11, 1033–1036. [Google Scholar] [CrossRef] [PubMed]
- Thurnham, D.I.; McCabe, L.D.; Haldar, S.; Wieringa, F.T.; Northrop-Clewes, C.A.; McCabe, G.P. Adjusting plasma ferritin concentrations to remove the effects of subclinical inflammation in the assessment of iron deficiency: A meta-analysis. Am. J. Clin. Nutr. 2010, 92, 546–555. [Google Scholar] [CrossRef] [Green Version]
- DePalma, R.G.; Hayes, V.W.; Chow, B.K.; Shamayeva, G.; May, P.E.; Zacharski, L.R. Ferritin levels, inflammatory biomarkers, and mortality in peripheral arterial disease: A substudy of the Iron (Fe) and Atherosclerosis Study (FeAST) Trial. J. Vasc. Surg. 2010, 51, 1498–1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Knovich, M.A.; Coffman, L.G.; Torti, F.M.; Torti, S.V. Serum ferritin: Past, present and future. Biochim. Biophys. Acta 2010, 1800, 760–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kernan, K.F.; Carcillo, J.A. Hyperferritinemia and inflammation. Int. Immunol. 2017, 29, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Zaritsky, J.; Young, B.; Wang, H.-J.; Westerman, M.; Olbina, G.; Nemeth, E.; Ganz, T.; Rivera, S.; Nissenson, A.R.; Salusky, I.B. Hepcidin—A potential novel biomarker for iron status in chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2009, 4, 1051–1056. [Google Scholar] [CrossRef]
- Ganz, T.; Nemeth, E. Iron Balance and the Role of Hepcidin in Chronic Kidney Disease. Semin. Nephrol. 2016, 36, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Ganz, T.; Nemeth, E. Hepcidin and iron homeostasis. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 1434–1443. [Google Scholar] [CrossRef] [Green Version]
- Blanchette, N.L.; Manz, D.H.; Torti, F.M.; Torti, S.V. Modulation of hepcidin to treat iron deregulation: Potential clinical applications. Expert Rev. Hematol. 2016, 9, 169–186. [Google Scholar] [CrossRef]
- Beguin, Y. Soluble transferrin receptor for the evaluation of erythropoiesis and iron status. Clin. Chim. Acta 2003, 329, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Rohner, F.; Namaste, S.M.; Larson, L.M.; Addo, O.Y.; Mei, Z.; Suchdev, P.S.; Williams, A.M.; Sakr Ashour, F.A.; Rawat, R.; Raiten, D.J.; et al. Adjusting soluble transferrin receptor concentrations for inflammation: Biomarkers Reflecting Inflammation and Nutritional Determinants of Anemia (BRINDA) project. Am. J. Clin. Nutr. 2017, 106, 372S–382S. [Google Scholar] [CrossRef] [PubMed]
- Sierpinski, R.; Josiak, K.; Suchocki, T.; Wojtas-Polc, K.; Mazur, G.; Butrym, A.; Rozentryt, P.; van der Meer, P.; Comin-Colet, J.; von Haehling, S.; et al. High soluble transferrin receptor in patients with heart failure: A measure of iron deficiency and a strong predictor of mortality. Eur. J. Heart Fail. 2020. [Google Scholar] [CrossRef] [PubMed]
- Harms, K.; Kaiser, T. Beyond soluble transferrin receptor: Old challenges and new horizons. Best Pract. Res. Clin. Endocrinol. Metab 2015, 29, 799–810. [Google Scholar] [CrossRef]
- Horinouchi, Y.; Ikeda, Y.; Tamaki, T. Body iron accumulation in obesity, diabetes and its complications, and the possibility of therapeutic application by iron regulation. Nippon Yakurigaku Zasshi 2019, 154, 316. [Google Scholar] [CrossRef] [PubMed]
- Sanad, M.; Osman, M.; Gharib, A. Obesity modulate serum hepcidin and treatment outcome of iron deficiency anemia in children: A case control study. Ital. J. Pediatric 2011, 37, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnweber, T.; Ress, C.; Nairz, M.; Theurl, I.; Schroll, A.; Murphy, A.T.; Wroblewski, V.; Witcher, D.R.; Moser, P.; Ebenbichler, C.F.; et al. High-fat diet causes iron deficiency via hepcidin-independent reduction of duodenal iron absorption. J. Nutr. Biochem. 2012, 23, 1600–1608. [Google Scholar] [CrossRef]
- Mujica-Coopman, M.F.; Brito, A.; de Romana, D.L.; Pizarro, F.; Olivares, M. Body mass index, iron absorption and iron status in childbearing age women. J. Trace Elem. Med. Biol. 2015, 30, 215–219. [Google Scholar] [CrossRef]
- Zimmermann, M.; Zeder, C.; Muthayya, S.; Winichagoon, P.; Chaouki, N.; Aeberli, I.; Hurrell, R. Adiposity in women and children from transition countries predicts decreased iron absorption, iron deficiency and a reduced response to iron fortification. Int. J. Obes. 2008, 32, 1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benotti, P.N.; Wood, G.C.; Still, C.D.; Gerhard, G.S.; Rolston, D.D.; Bistrian, B.R. Metabolic surgery and iron homeostasis. Obes. Rev. 2019, 20, 612–620. [Google Scholar] [CrossRef]
- Dixon, J.B.; Zimmet, P.; Alberti, K.G.; Rubino, F.; International Diabetes Federation Taskforce on Epidemiology and Prevention. Bariatric surgery: An IDF statement for obese Type 2 diabetes. Diabet. Med. 2011, 28, 628–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haluzik, M. Bariatric/metabolic surgery: The diabetologists view. Rozhl. Chir. 2019, 98, 46–51. [Google Scholar]
- Schauer, P.R.; Rubino, F. International Diabetes Federation position statement on bariatric surgery for type 2 diabetes: Implications for patients, physicians, and surgeons. Surg. Obes. Relat. Dis. 2011, 7, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Brito, J.P.; Montori, V.M.; Davis, A.M. Metabolic Surgery in the Treatment Algorithm for Type 2 Diabetes: A Joint Statement by International Diabetes Organizations. JAMA 2017, 317, 635–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.; Bryant, C.; Cook, R.; O’connor, H.; Rooney, K.; Steinbeck, K. The relationship between obesity and hypoferraemia in adults: A systematic review. Obes. Rev. 2012, 13, 150–161. [Google Scholar] [CrossRef]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A.R. Adipocytokines: Mediators linking adipose tissue, inflammation and immunity. Nat. Rev. Immunol. 2006, 6, 772–783. [Google Scholar] [CrossRef]
- Gasmi, A.; Peana, M.; Pivina, L.; Srinath, S.; Benahmed, A.G.; Semenova, Y.; Menzel, A.; Dadar, M.; Bjørklund, G. Interrelations between COVID-19 and other disorders. Clin. Immunol. 2020, 108651. [Google Scholar] [CrossRef]
- Schwenger, K.J.; Allard, J.P. Clinical approaches to non-alcoholic fatty liver disease. World J. Gastroenterol. 2014, 20, 1712. [Google Scholar] [CrossRef]
- Amato, A.; Santoro, N.; Calabro, P.; Grandone, A.; Swinkels, D.; Perrone, L.; Del Giudice, E.M. Effect of body mass index reduction on serum hepcidin levels and iron status in obese children. Int. J. Obes. 2010, 34, 1772. [Google Scholar] [CrossRef] [Green Version]
- Gong, L.; Yuan, F.; Teng, J.; Li, X.; Zheng, S.; Lin, L.; Deng, H.; Ma, G.; Sun, C.; Li, Y. Weight loss, inflammatory markers, and improvements of iron status in overweight and obese children. J. Pediatrics 2014, 164, 795–800.e2. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-Y.; Sabrina, N.; Chien, Y.-W.; Chen, Y.-C.; Lin, S.-H.; Chang, J.-S. A Moderate Interleukin-6 Reduction, Not a Moderate Weight Reduction, Improves the Serum Iron Status in Diet-Induced Weight Loss with Fish Oil Supplementation. Mol. Nutr. Food Res. 2018, 62, 1800243. [Google Scholar] [CrossRef] [PubMed]
- Aktas, G.; Alcelik, A.; Yalcin, A.; Karacay, S.; Kurt, S.; Akduman, M.; Savli, H. Treatment of iron deficiency anemia induces weight loss and improves metabolic parameters. Clin. Ter. 2014, 165, e87–e89. [Google Scholar] [PubMed]
- Pivina, L.; Semenova, Y.; Doşa, M.D.; Dauletyarova, M.; Bjørklund, G. Iron Deficiency, Cognitive Functions, and Neurobehavioral Disorders in Children. J. Mol. Neurosci. 2019, 68, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kanamori, Y.; Murakami, M.; Sugiyama, M.; Hashimoto, O.; Matsui, T.; Funaba, M. Hepcidin and IL-1beta. Vitam. Horm. 2019, 110, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Leitch, H.A. Optimizing therapy for iron overload in the myelodysplastic syndromes: Recent developments. Drugs 2011, 71, 155–177. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Hepcidin--A peptide hormone at the interface of innate immunity and iron metabolism. Curr. Top. Microbiol. Immunol. 2006, 306, 183–198. [Google Scholar] [CrossRef]
- Nemeth, E.; Valore, E.V.; Territo, M.; Schiller, G.; Lichtenstein, A.; Ganz, T. Hepcidin, a putative mediator of anemia of inflammation, is a type II acute-phase protein. Blood 2003, 101, 2461–2463. [Google Scholar] [CrossRef]
- Nairz, M.; Weiss, G. Iron in infection and immunity. Mol. Aspects Med. 2020, 75, 100864. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860. [Google Scholar] [CrossRef]
- Gasmi, A.; Noor, S.; Menzel, A.; Dosa, A.; Pivina, L.; Bjørklund, G. Obesity and Insulin Resistance: Associations with Chronic Inflammation, Genetic and Epigenetic Factors. Curr. Med. Chem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Stolarczyk, E. Adipose tissue inflammation in obesity: A metabolic or immune response? Curr. Opin. Pharmacol. 2017, 37, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Real, J.M.; Ricart, W. Insulin resistance and chronic cardiovascular inflammatory syndrome. Endocr. Rev. 2003, 24, 278–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawano, Y.; Nakae, J.; Watanabe, N.; Kikuchi, T.; Tateya, S.; Tamori, Y.; Kaneko, M.; Abe, T.; Onodera, M.; Itoh, H. Colonic Pro-inflammatory Macrophages Cause Insulin Resistance in an Intestinal Ccl2/Ccr2-Dependent Manner. Cell Metab. 2016, 24, 295–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.H.; Cho, M.H.; Kim, J.Y.; Kwon, M.S.; Peak, J.J.; Kang, S.W.; Yoon, S.Y.; Song, Y. Impaired macrophage autophagy induces systemic insulin resistance in obesity. Oncotarget 2016, 7, 35577–35591. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Sethi, J.K.; Hotamisligil, G.S. Transmembrane Tumor Necrosis Factor (TNF)-α Inhibits Adipocyte Differentiation by Selectively Activating TNF Receptor 1. J. Biol. Chem. 1999, 274, 26287–26295. [Google Scholar] [CrossRef] [Green Version]
- Bulló, M.; García-Lorda, P.; Peinado-Onsurbe, J.; Hernández, M.; Del Castillo, D.; Argilés, J.M.; Salas-Salvadó, J. TNFα expression of subcutaneous adipose tissue in obese and morbid obese females: Relationship to adipocyte LPL activity and leptin synthesis. Int. J. Obes. 2002, 26, 652–658. [Google Scholar] [CrossRef] [Green Version]
- Lyon, C.J.; Law, R.E.; Hsueh, W.A. Minireview: Adiposity, inflammation, and atherogenesis. Endocrinology 2003, 144, 2195–2200. [Google Scholar] [CrossRef]
- Mohamed-Ali, V.; Goodrick, S.; Rawesh, A.; Katz, D.R.; Miles, J.M.; Yudkin, J.S.; Klein, S.; Coppack, S.W. Subcutaneous Adipose Tissue Releases Interleukin-6, But Not Tumor Necrosis Factor-α, in Vivo1. J. Clin. Endocrinol. Metab. 1997, 82, 4196–4200. [Google Scholar] [CrossRef] [Green Version]
- Banos, G.; Medina-Campos, O.N.; Maldonado, P.D.; Zamora, J.; Perez, I.; Pavon, N.; Pedraza-Chaverri, J. Antioxidant enzymes in hypertensive and hypertriglyceridemic rats: Effect of gender. Clin. Exp. Hypertens 2005, 27, 45–57. [Google Scholar] [CrossRef]
- Fernandez-Real, J.M.; Vayreda, M.; Richart, C.; Gutierrez, C.; Broch, M.; Vendrell, J.; Ricart, W. Circulating interleukin 6 levels, blood pressure, and insulin sensitivity in apparently healthy men and women. J. Clin. Endocrinol. Metab. 2001, 86, 1154–1159. [Google Scholar] [CrossRef] [PubMed]
- Keaney, J.F., Jr.; Larson, M.G.; Vasan, R.S.; Wilson, P.W.; Lipinska, I.; Corey, D.; Massaro, J.M.; Sutherland, P.; Vita, J.A.; Benjamin, E.J. Obesity and systemic oxidative stress: Clinical correlates of oxidative stress in the Framingham Study. Arter. Thromb. Vasc. Biol. 2003, 23, 434–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambhojan, C.; Larifla, L.; Clepier, J.; Bouaziz-Amar, E.; Velayoudom-Cephise, F.-L.; Blanchet-Deverly, A.; Armand, C.; Plumasseau, J.; Lacorte, J.-M.; Foucan, L. Vitamin D status, insulin resistance, leptin-to-adiponectin ratio in adolescents: Results of a 1-year lifestyle intervention. Open Access Maced. J. Med. Sci. 2016, 4, 596. [Google Scholar] [CrossRef] [Green Version]
- Hukshorn, C.J.; Lindeman, J.H.; Toet, K.H.; Saris, W.H.; Eilers, P.H.; Westerterp-Plantenga, M.S.; Kooistra, T. Leptin and the proinflammatory state associated with human obesity. J. Clin. Endocrinol. Metab. 2004, 89, 1773–1778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fantuzzi, G.; Faggioni, R. Leptin in the regulation of immunity, inflammation, and hematopoiesis. J. Leukoc. Biol. 2000, 68, 437–446. [Google Scholar] [PubMed]
- Bełtowski, J. Apelin and visfatin: Unique. Med. Sci. Monit. 2006, 12, RA112–RA119. [Google Scholar] [PubMed]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative stress in obesity: A critical component in human diseases. Int. J. Mol. Sci. 2015, 16, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Martos-Moreno, G.Á.; Kratzsch, J.; Körner, A.; Barrios, V.; Hawkins, F.; Kiess, W.; Argente, J. Serum visfatin and vaspin levels in prepubertal children: Effect of obesity and weight loss after behavior modifications on their secretion and relationship with glucose metabolism. Int. J. Obes. 2011, 35, 1355. [Google Scholar] [CrossRef] [Green Version]
- Pihl, E.; Zilmer, K.; Kullisaar, T.; Kairane, C.; Mägi, A.; Zilmer, M. Atherogenic inflammatory and oxidative stress markers in relation to overweight values in male former athletes. Int. J. Obes. 2006, 30, 141. [Google Scholar] [CrossRef] [Green Version]
- Chrysohoou, C.; Panagiotakos, D.B.; Pitsavos, C.; Skoumas, I.; Papademetriou, L.; Economou, M.; Stefanadis, C. The implication of obesity on total antioxidant capacity in apparently healthy men and women: The ATTICA study. Nutr. Metab. Cardiovasc. Dis. 2007, 17, 590–597. [Google Scholar] [CrossRef]
- le Roux, C.W.; Heneghan, H.M. Bariatric Surgery for Obesity. Med. Clin. N. Am. 2018, 102, 165–182. [Google Scholar] [CrossRef] [PubMed]
- Salgado, W., Jr.; Modotti, C.; Nonino, C.B.; Ceneviva, R. Anemia and iron deficiency before and after bariatric surgery. Surg. Obes. Relat. Dis. 2014, 10, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Khanbhai, M.; Dubb, S.; Patel, K.; Ahmed, A.; Richards, T. The prevalence of iron deficiency anaemia in patients undergoing bariatric surgery. Obes. Res. Clin. Pract. 2015, 9, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Ten Broeke, R.; Bravenboer, B.; Smulders, F.J. Iron deficiency before and after bariatric surgery: The need for iron supplementation. Neth. J. Med. 2013, 71, 412–417. [Google Scholar]
- Careaga, M.; Moize, V.; Flores, L.; Deulofeu, R.; Andreu, A.; Vidal, J. Inflammation and iron status in bariatric surgery candidates. Surg. Obes. Relat. Dis. 2015, 11, 906–911. [Google Scholar] [CrossRef] [PubMed]
- Gesquiere, I.; Lannoo, M.; Augustijns, P.; Matthys, C.; Van der Schueren, B.; Foulon, V. Iron deficiency after Roux-en-Y gastric bypass: Insufficient iron absorption from oral iron supplements. Obes. Surg. 2014, 24, 56–61. [Google Scholar] [CrossRef] [Green Version]
- Boyce, S.G.; Goriparthi, R.; Clark, J.; Cameron, K.; Roslin, M.S. Can Composite Nutritional Supplement Based on the Current Guidelines Prevent Vitamin and Mineral Deficiency after Weight Loss Surgery? Obes. Surg. 2016, 26, 966–971. [Google Scholar] [CrossRef]
- Del Villar Madrigal, E.; Neme-Yunes, Y.; Clavellina-Gaytan, D.; Sanchez, H.A.; Mosti, M.; Herrera, M.F. Anemia After Roux-en-Y Gastric Bypass. How Feasible to Eliminate the Risk by Proper Supplementation? Obes. Surg. 2015, 25, 80–84. [Google Scholar] [CrossRef]
- Montano-Pedroso, J.C.; Garcia, E.B.; Omonte, I.R.; Rocha, M.G.; Ferreira, L.M. Hematological variables and iron status in abdominoplasty after bariatric surgery. Obes. Surg. 2013, 23, 7–16. [Google Scholar] [CrossRef]
- Monaco-Ferreira, D.V.; Leandro-Merhi, V.A. Status of iron metabolism 10 years after Roux-en-Y gastric bypass. Obes. Surg. 2017, 27, 1993–1999. [Google Scholar] [CrossRef]
- Ferreira, M.J.S.; Salazar, D.A.; Neves, J.S.; Pedro, J.M.P.; Guerreiro, V.A.; Viana, S.e.S.; Mendonça, F.; Silva, M.M.; Belo, S.P.; Sande, A.V.; et al. Which Factors Are Associated with a Higher Prevalence of Anemia Following Bariatric Surgery? Results from a Retrospective Study Involving 1999 Patients. Obes. Surg. 2020, 30, 3496–3502. [Google Scholar] [CrossRef] [PubMed]
- Bailly, L.; Schiavo, L.; Sebastianelli, L.; Fabre, R.; Pradier, C.; Iannelli, A. Anemia and Bariatric Surgery: Results of a National French Survey on Administrative Data of 306,298 Consecutive Patients Between 2008 and 2016. Obes. Surg. 2018, 28, 2313–2320. [Google Scholar] [CrossRef] [PubMed]
- Tussing-Humphreys, L.M.; Nemeth, E.; Fantuzzi, G.; Freels, S.; Holterman, A.X.L.; Galvani, C.; Ayloo, S.; Vitello, J.; Braunschweig, C. Decreased serum hepcidin and improved functional iron status 6 months after restrictive bariatric surgery. Obesity 2010, 18, 2010–2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cepeda-Lopez, A.C.; Allende-Labastida, J.; Melse-Boonstra, A.; Osendarp, S.J.; Herter-Aeberli, I.; Moretti, D.; Rodriguez-Lastra, R.; Gonzalez-Salazar, F.; Villalpando, S.; Zimmermann, M.B. The effects of fat loss after bariatric surgery on inflammation, serum hepcidin, and iron absorption: A prospective 6-mo iron stable isotope study. Am. J. Clin. Nutr. 2016, 104, 1030–1038. [Google Scholar] [CrossRef] [PubMed]
- Enani, G.; Bilgic, E.; Lebedeva, E.; Delisle, M.; Vergis, A.; Hardy, K. The incidence of iron deficiency anemia post-Roux-en-Y gastric bypass and sleeve gastrectomy: A systematic review. Surg. Endosc. 2020, 34, 3002–3010. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.; Kim, H.J.; Lo Menzo, E.; Park, S.; Szomstein, S.; Rosenthal, R.J. Anemia, iron and vitamin B12 deficiencies after sleeve gastrectomy compared to Roux-en-Y gastric bypass: A meta-analysis. Surg. Obes. Relat. Dis. 2014, 10, 589–597. [Google Scholar] [CrossRef]
- Casgrain, A.; Collings, R.; Harvey, L.J.; Hooper, L.; Fairweather-Tait, S.J. Effect of iron intake on iron status: A systematic review and meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2012, 96, 768–780. [Google Scholar] [CrossRef] [Green Version]
- Gera, T.; Sachdev, H.S.; Boy, E. Effect of iron-fortified foods on hematologic and biological outcomes: Systematic review of randomized controlled trials. Am. J. Clin. Nutr. 2012, 96, 309–324. [Google Scholar] [CrossRef] [Green Version]
- Ben-Porat, T.; Elazary, R.; Sherf-Dagan, S.; Weiss, R.; Levin, G.; Rottenstreich, M.; Sakran, N.; Rottenstreich, A. Factors Associated with the Development of Anemia During Pregnancy after Sleeve Gastrectomy. Obes. Surg. 2020, 30, 3884–3890. [Google Scholar] [CrossRef]
- Sandvik, J.; Hole, T.; Klockner, C.A.; Kulseng, B.E.; Wibe, A. Intravenous Iron Treatment in the Prevention of Iron Deficiency and Anaemia after Roux-en-Y Gastric Bypass. Obes. Surg. 2020, 30, 1745–1752. [Google Scholar] [CrossRef] [Green Version]
- Rozanska-Waledziak, A.; Bartnik, P.; Kacperczyk-Bartnik, J.; Czajkowski, K.; Waledziak, M. The Impact of Bariatric Surgery on Menstrual Abnormalities-a Cross-Sectional Study. Obes. Surg. 2020, 30, 4505–4509. [Google Scholar] [CrossRef]
- Teitelman, M.; Grotegut, C.A.; Williams, N.N.; Lewis, J.D. The impact of bariatric surgery on menstrual patterns. Obes. Surg. 2006, 16, 1457–1463. [Google Scholar] [CrossRef]
- Tan, O.; Carr, B.R. The impact of bariatric surgery on obesity-related infertility and in vitro fertilization outcomes. Semin. Reprod. Med. 2012, 30, 517–528. [Google Scholar] [CrossRef]
- Mischler, R.A.; Armah, S.M.; Wright, B.N.; Mattar, S.G.; Rosen, A.D.; Gletsu-Miller, N. Influence of diet and supplements on iron status after gastric bypass surgery. Surg. Obes. Relat. Dis. 2016, 12, 651–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechanick, J.I.; Youdim, A.; Jones, D.B.; Garvey, W.T.; Hurley, D.L.; McMahon, M.M.; Heinberg, L.J.; Kushner, R.; Adams, T.D.; Shikora, S.; et al. Clinical practice guidelines for the perioperative nutritional, metabolic, and nonsurgical support of the bariatric surgery patient—2013 update: Cosponsored by American Association of Clinical Endocrinologists, The Obesity Society, and American Society for Metabolic & Bariatric Surgery. Obesity 2013, 21 (Suppl. 1), S1–S27. [Google Scholar] [CrossRef]
- Mischler, R.A.; Armah, S.M.; Craig, B.A.; Rosen, A.D.; Banerjee, A.; Selzer, D.J.; Choi, J.N.; Gletsu-Miller, N. Comparison of Oral Iron Supplement Formulations for Normalization of Iron Status Following Roux-EN-y Gastric Bypass Surgery: A Randomized Trial. Obes. Surg. 2018, 28, 369–377. [Google Scholar] [CrossRef]
- Snell-Sparapany, H.; Choi, J.; Rosen, A.; Reddy, M.; Gletsu-Miller, N. Bioavailability of Iron Supplement Formulations by Bariatric Surgery Type: A Pilot Study (P23-013-19). Curr. Dev. Nutr. 2019, 3. [Google Scholar] [CrossRef] [Green Version]
- Palermo, A.; Naciu, A.M.; Tabacco, G.; Manfrini, S.; Trimboli, P.; Vescini, F.; Falchetti, A. Calcium citrate: From biochemistry and physiology to clinical applications. Rev. Endocr. Metab. Disord. 2019, 20, 353–364. [Google Scholar] [CrossRef]
- Parrott, J.; Frank, L.; Rabena, R.; Craggs-Dino, L.; Isom, K.A.; Greiman, L. American Society for Metabolic and Bariatric Surgery Integrated Health Nutritional Guidelines for the Surgical Weight Loss Patient 2016 Update: Micronutrients. Surg. Obes. Relat. Dis. 2017, 13, 727–741. [Google Scholar] [CrossRef]
- Bjørklund, G.; Semenova, Y.; Pivina, L.; Costea, D.O. Follow-up after bariatric surgery: A review. Nutrition 2020, 78, 110831. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bjørklund, G.; Peana, M.; Pivina, L.; Dosa, A.; Aaseth, J.; Semenova, Y.; Chirumbolo, S.; Medici, S.; Dadar, M.; Costea, D.-O. Iron Deficiency in Obesity and after Bariatric Surgery. Biomolecules 2021, 11, 613. https://doi.org/10.3390/biom11050613
Bjørklund G, Peana M, Pivina L, Dosa A, Aaseth J, Semenova Y, Chirumbolo S, Medici S, Dadar M, Costea D-O. Iron Deficiency in Obesity and after Bariatric Surgery. Biomolecules. 2021; 11(5):613. https://doi.org/10.3390/biom11050613
Chicago/Turabian StyleBjørklund, Geir, Massimiliano Peana, Lyudmila Pivina, Alexandru Dosa, Jan Aaseth, Yuliya Semenova, Salvatore Chirumbolo, Serenella Medici, Maryam Dadar, and Daniel-Ovidiu Costea. 2021. "Iron Deficiency in Obesity and after Bariatric Surgery" Biomolecules 11, no. 5: 613. https://doi.org/10.3390/biom11050613
APA StyleBjørklund, G., Peana, M., Pivina, L., Dosa, A., Aaseth, J., Semenova, Y., Chirumbolo, S., Medici, S., Dadar, M., & Costea, D. -O. (2021). Iron Deficiency in Obesity and after Bariatric Surgery. Biomolecules, 11(5), 613. https://doi.org/10.3390/biom11050613