
Role of Biomolecules in Osteoclasts and Their Therapeutic Potential for Osteoporosis


Abstract
:1. Introduction
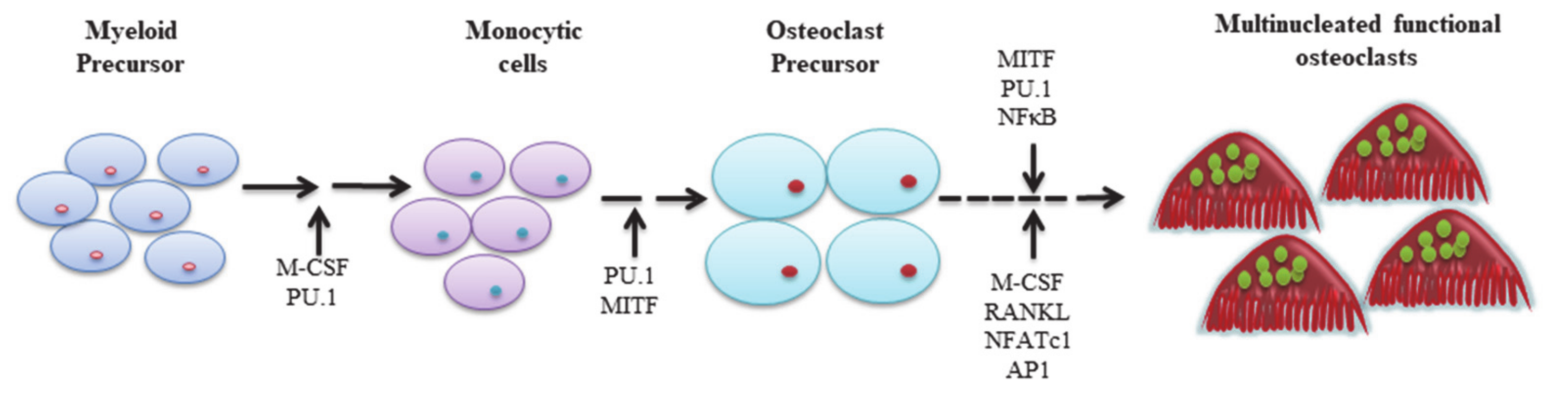
2. The Role of Molecules in Osteoclast Regulation
2.1. Cytokines
2.1.1. PU.1
2.1.2. M-CSF
2.1.3. c-Fos
2.1.4. C/EBPα
2.1.5. RANK/RANKL/Osteoprotegerin (OPG)
2.1.6. NFATc1
2.1.7. MITF
2.2. Hormone
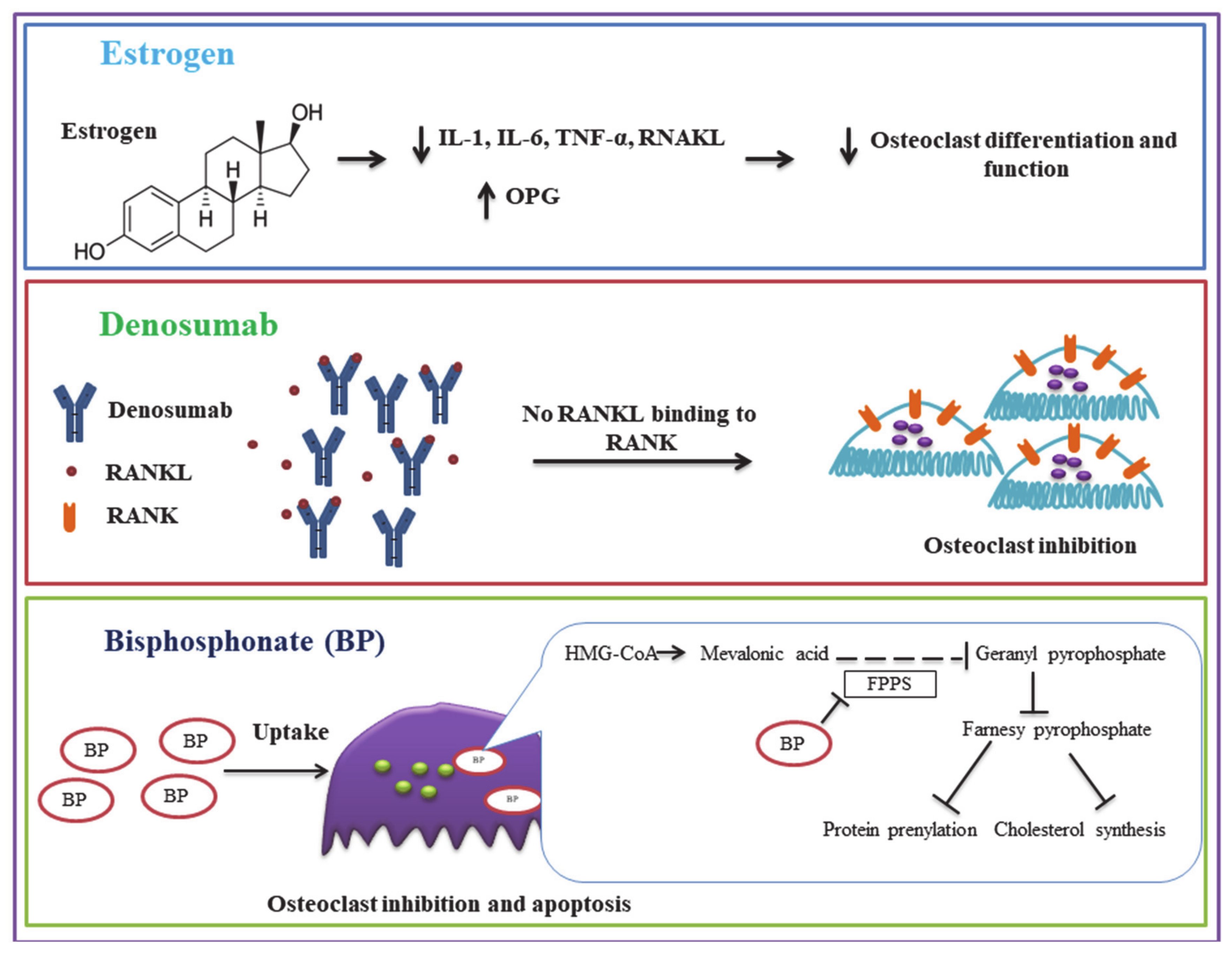
2.2.1. Estrogen
2.2.2. Parathyroid Hormone (PTH)
2.2.3. Calcitonin
2.3. V-ATPase
2.4. Non-Coding RNA
2.5.1. MicroRNA (MiRNA)
2.5.2. Long Non-Coding RNA (LncRNA)
2.5.3. Circular RNA (CircRNA)
3. The Role of Osteoclasts in Bone Resorption
4. Osteoporosis
Osteoclast Targeted Osteoporosis Treatments
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hadjidakis, D.J.; Androulakis, I.I. Bone remodeling. Ann. N. Y. Acad. Sci. 2006, 1092, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, J.A.; Partridge, N.C. Physiological Bone Remodeling: Systemic Regulation and Growth Factor Involvement. Physiology 2016, 31, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast-osteoclast interactions. Connect. Tissue Res. 2018, 59, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Raggatt, L.J.; Partridge, N.C. Cellular and molecular mechanisms of bone remodeling. J. Biol. Chem. 2010, 285, 25103–25108. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, K.; Takeshita, S. The role of osteoclast differentiation and function in skeletal homeostasis. J. Biochem. 2016, 159, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Lane, J.M.; Russell, L.; Khan, S.N. Osteoporosis. Clin. Orthopaed. Relat. Res. 2000, 139–150. [Google Scholar] [CrossRef]
- Binkley, N.; Krueger, D. Osteoporosis in men. WMJ Off. Publ. St. Med. Soc. Wis. 2002, 101, 28–32. [Google Scholar] [CrossRef] [Green Version]
- Sözen, T.; Özışık, L.; Başaran, N.Ç. An overview and management of osteoporosis. Eur. J. Rheumatol. 2017, 4, 46–56. [Google Scholar] [CrossRef]
- Teitelbaum, S.L.; Ross, F.P. Genetic regulation of osteoclast development and function. Nat. Rev. Genet. 2003, 4, 638–649. [Google Scholar] [CrossRef]
- Dahl, R.; Walsh, J.C.; Lancki, D.; Laslo, P.; Iyer, S.R.; Singh, H.; Simon, M.C. Regulation of macrophage and neutrophil cell fates by the PU.1:C/EBPalpha ratio and granulocyte colony-stimulating factor. Nat. Immunol. 2003, 4, 1029–1036. [Google Scholar] [CrossRef]
- Yang, D.Q.; Feng, S.; Chen, W.; Zhao, H.; Paulson, C.; Li, Y.P. V-ATPase subunit ATP6AP1 (Ac45) regulates osteoclast differentiation, extracellular acidification, lysosomal trafficking, and protease exocytosis in osteoclast-mediated bone resorption. J. Bone Miner. Res. 2012, 27, 1695–1707. [Google Scholar] [CrossRef] [Green Version]
- Oikawa, T.; Yamada, T. Molecular biology of the Ets family of transcription factors. Gene 2003, 303, 11–34. [Google Scholar] [CrossRef]
- Tondravi, M.M.; McKercher, S.R.; Anderson, K.; Erdmann, J.M.; Quiroz, M.; Maki, R.; Teitelbaum, S.L. Osteopetrosis in mice lacking haematopoietic transcription factor PU.1. Nature 1997, 386, 81–84. [Google Scholar] [CrossRef]
- Ishiyama, K.; Yashiro, T.; Nakano, N.; Kasakura, K.; Miura, R.; Hara, M.; Kawai, F.; Maeda, K.; Tamura, N.; Okumura, K.; et al. Involvement of PU.1 in NFATc1 promoter function in osteoclast development. Allergol. Int. 2015, 64, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, M.; Kogawa, M.; Wada, S.; Takayanagi, H.; Tsujimoto, M.; Katayama, S.; Hisatake, K.; Nogi, Y. Essential role of p38 mitogen-activated protein kinase in cathepsin K gene expression during osteoclastogenesis through association of NFATc1 and PU.1. J. Biol. Chem. 2004, 279, 45969–45979. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.M.; Bronisz, A.; Hu, R.; Patel, K.; Mansky, K.C.; Sif, S.; Ostrowski, M.C. MITF and PU.1 recruit p38 MAPK and NFATc1 to target genes during osteoclast differentiation. J. Biol. Chem. 2007, 282, 15921–15929. [Google Scholar] [CrossRef] [Green Version]
- Izawa, N.; Kurotaki, D.; Nomura, S.; Fujita, T.; Omata, Y.; Yasui, T.; Hirose, J.; Matsumoto, T.; Saito, T.; Kadono, Y.; et al. Cooperation of PU.1 With IRF8 and NFATc1 Defines Chromatin Landscapes During RANKL-Induced Osteoclastogenesis. J. Bone Miner. Res. 2019, 34, 1143–1154. [Google Scholar] [CrossRef]
- Jin, X.; Kruth, H.S. Culture of Macrophage Colony-stimulating Factor Differentiated Human Monocyte-derived Macrophages. J. Vis. Exp. JoVE 2016. [Google Scholar] [CrossRef]
- Ushach, I.; Zlotnik, A. Biological role of granulocyte macrophage colony-stimulating factor (GM-CSF) and macrophage colony-stimulating factor (M-CSF) on cells of the myeloid lineage. J. Leukocyte Biol. 2016, 100, 481–489. [Google Scholar] [CrossRef]
- Owens, J.; Chambers, T.J. Macrophage colony-stimulating factor (M-CSF) induces migration in osteoclasts in vitro. Biochem. Biophys. Res. Commun. 1993, 195, 1401–1407. [Google Scholar] [CrossRef]
- Umeda, S.; Takahashi, K.; Naito, M.; Shultz, L.D.; Takagi, K. Neonatal changes of osteoclasts in osteopetrosis (op/op) mice defective in production of functional macrophage colony-stimulating factor (M-CSF) protein and effects of M-CSF on osteoclast development and differentiation. J. Submicrosc. Cytol. Pathol. 1996, 28, 13–26. [Google Scholar]
- Hodge, J.M.; Collier, F.M.; Pavlos, N.J.; Kirkland, M.A.; Nicholson, G.C. M-CSF potently augments RANKL-induced resorption activation in mature human osteoclasts. PLoS ONE 2011, 6, e21462. [Google Scholar] [CrossRef] [Green Version]
- Hattersley, G.; Owens, J.; Flanagan, A.M.; Chambers, T.J. Macrophage colony stimulating factor (M-CSF) is essential for osteoclast formation in vitro. Biochem. Biophys. Res. Commun. 1991, 177, 526–531. [Google Scholar] [CrossRef]
- Ross, F.P.; Teitelbaum, S.L. alphavbeta3 and macrophage colony-stimulating factor: Partners in osteoclast biology. Immunol. Rev. 2005, 208, 88–105. [Google Scholar] [CrossRef]
- Ross, F.P. M-CSF, c-Fms, and signaling in osteoclasts and their precursors. Ann. N. Y. Acad. Sci. 2006, 1068, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Fujiwara, T.; Ye, S.; Li, X.; Zhao, H. Ubiquitin E3 Ligase LNX2 is Critical for Osteoclastogenesis In Vitro by Regulating M-CSF/RANKL Signaling and Notch2. Calcif. Tissue Int. 2015, 96, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Wagner, E.F. Bone development and inflammatory disease is regulated by AP-1 (Fos/Jun). Ann. Rheum. Dis. 2010, 69, i86–i88. [Google Scholar] [CrossRef]
- Miyamoto, T.; Ohneda, O.; Arai, F.; Iwamoto, K.; Okada, S.; Takagi, K.; Anderson, D.M.; Suda, T. Bifurcation of osteoclasts and dendritic cells from common progenitors. Blood 2001, 98, 2544–2554. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, K.; Ray, N. Osteoclasts, mononuclear phagocytes, and c-Fos: New insight into osteoimmunology. Keio J. Med. 2004, 53, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Zhu, G.; Hao, L.; Wu, M.; Ci, H.; Li, Y.P. C/EBPalpha regulates osteoclast lineage commitment. Proc. Natl. Acad. Sci. USA 2013, 110, 7294–7299. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Zhu, G.; Tang, J.; Zhou, H.D.; Li, Y.P. C/ebpα controls osteoclast terminal differentiation, activation, function, and postnatal bone homeostasis through direct regulation of Nfatc1. J. Pathol. 2018, 244, 271–282. [Google Scholar] [CrossRef]
- Jules, J.; Chen, W.; Feng, X.; Li, Y.P. CCAAT/Enhancer-binding Protein α (C/EBPα) Is Important for Osteoclast Differentiation and Activity. J. Biol. Chem. 2016, 291, 16390–16403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jules, J.; Chen, W.; Feng, X.; Li, Y.P. C/EBPα transcription factor is regulated by the RANK cytoplasmic (535)IVVY(538) motif and stimulates osteoclastogenesis more strongly than c-Fos. J. Biol. Chem. 2018, 293, 1480–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigl, V.; Jones, L.P.; Penninger, J.M. RANKL/RANK: From bone loss to the prevention of breast cancer. Open Biol. 2016, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyce, B.F.; Xing, L. Biology of RANK, RANKL, and osteoprotegerin. Arthritis Res. Ther. 2007, 9, S1. [Google Scholar] [CrossRef] [Green Version]
- Fata, J.E.; Kong, Y.-Y.; Li, J.; Sasaki, T.; Irie-Sasaki, J.; Moorehead, R.A.; Elliott, R.; Scully, S.; Voura, E.B.; Lacey, D.L. The osteoclast differentiation factor osteoprotegerin-ligand is essential for mammary gland development. Cell 2000, 103, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Sobacchi, C.; Menale, C.; Villa, A. The RANKL-RANK Axis: A Bone to Thymus Round Trip. Front. Immunol. 2019, 10, 629. [Google Scholar] [CrossRef] [Green Version]
- Nagy, V.; Penninger, J.M. The RANKL-RANK Story. Gerontology 2015, 61, 534–542. [Google Scholar] [CrossRef]
- Boyce, B.F.; Xing, L. Functions of RANKL/RANK/OPG in bone modeling and remodeling. Arch. Biochem. Biophys. 2008, 473, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Grimaud, E.; Soubigou, L.; Couillaud, S.; Coipeau, P.; Moreau, A.; Passuti, N.; Gouin, F.; Redini, F.; Heymann, D. Receptor activator of nuclear factor kappaB ligand (RANKL)/osteoprotegerin (OPG) ratio is increased in severe osteolysis. Am. J. Pathol. 2003, 163, 2021–2031. [Google Scholar] [CrossRef]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, X. Receptor activator of nuclear factor-kappaB ligand (RANKL)/RANK/osteoprotegerin system in bone and other tissues (review). Mol. Med. Rep. 2015, 11, 3212–3218. [Google Scholar] [CrossRef] [Green Version]
- Ikebuchi, Y.; Aoki, S.; Honma, M.; Hayashi, M.; Sugamori, Y.; Khan, M.; Kariya, Y.; Kato, G.; Tabata, Y.; Penninger, J.M.; et al. Coupling of bone resorption and formation by RANKL reverse signalling. Nature 2018, 561, 195–200. [Google Scholar] [CrossRef]
- Yasuda, H.; Shima, N.; Nakagawa, N.; Yamaguchi, K.; Kinosaki, M.; Mochizuki, S.; Tomoyasu, A.; Yano, K.; Goto, M.; Murakami, A.; et al. Osteoclast differentiation factor is a ligand for osteoprotegerin/osteoclastogenesis-inhibitory factor and is identical to TRANCE/RANKL. Proc. Natl. Acad. Sci. USA 1998, 95, 3597–3602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacey, D.L.; Timms, E.; Tan, H.L.; Kelley, M.J.; Dunstan, C.R.; Burgess, T.; Elliott, R.; Colombero, A.; Elliott, G.; Scully, S.; et al. Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 1998, 93, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kim, K.; Youn, B.U.; Jin, H.M.; Kim, J.-Y.; Moon, J.B.; Ko, A.; Seo, S.-B.; Lee, K.-Y.; Kim, N. RANKL induces NFATc1 acetylation and stability via histone acetyltransferases during osteoclast differentiation. Biochem. J. 2011, 436, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Yang, Z.; Ma, Y.; Yue, Z.; Lin, H.; Qu, G.; Huang, J.; Dai, W.; Li, C.; Zheng, C.; et al. LGR4 is a receptor for RANKL and negatively regulates osteoclast differentiation and bone resorption. Nat. Med. 2016, 22, 539–546. [Google Scholar] [CrossRef]
- Asagiri, M.; Sato, K.; Usami, T.; Ochi, S.; Nishina, H.; Yoshida, H.; Morita, I.; Wagner, E.F.; Mak, T.W.; Serfling, E.; et al. Autoamplification of NFATc1 expression determines its essential role in bone homeostasis. J. Exp. Med. 2005, 202, 1261–1269. [Google Scholar] [CrossRef] [Green Version]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.; et al. Induction and activation of the transcription factor NFATc1 (NFAT2) integrate RANKL signaling in terminal differentiation of osteoclasts. Dev. Cell 2002, 3, 889–901. [Google Scholar] [CrossRef] [Green Version]
- Takayanagi, H. The role of NFAT in osteoclast formation. Ann. N. Y. Acad. Sci. 2007, 1116, 227–237. [Google Scholar] [CrossRef]
- Crabtree, G.R.; Olson, E.N. NFAT signaling: Choreographing the social lives of cells. Cell 2002, 109, S67–S79. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, K.; Galson, D.L.; Zhao, C.; Peng, L.; Laplace, C.; Wang, K.Z.; Bachler, M.A.; Amano, H.; Aburatani, H.; Ishikawa, H. Nuclear factor of activated T-cells (NFAT) rescues osteoclastogenesis in precursors lacking c-Fos. J. Biol. Chem. 2004, 279, 26475–26480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crotti, T.N.; Flannery, M.; Walsh, N.C.; Fleming, J.D.; Goldring, S.R.; McHugh, K.P. NFATc1 regulation of the human β3 integrin promoter in osteoclast differentiation. Gene 2006, 372, 92–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bronisz, A.; Sharma, S.M.; Hu, R.; Godlewski, J.; Tzivion, G.; Mansky, K.C.; Ostrowski, M.C. Microphthalmia-associated transcription factor interactions with 14-3-3 modulate differentiation of committed myeloid precursors. Mol. Biol. Cell 2006, 17, 3897–3906. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, T.; Murphy, S.; Sohn, C.; Mansky, K.C. C-TAK1 interacts with microphthalmia-associated transcription factor, Mitf, but not the related family member Tfe3. Biochem. Biophys. Res. Commun. 2010, 394, 890–895. [Google Scholar] [CrossRef] [Green Version]
- Bronisz, A.; Carey, H.A.; Godlewski, J.; Sif, S.; Ostrowski, M.C.; Sharma, S.M. The multifunctional protein fused in sarcoma (FUS) is a coactivator of microphthalmia-associated transcription factor (MITF). J. Biol. Chem. 2014, 289, 326–334. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Cheng, T.; Steer, J.H.; Joyce, D.A.; Pavlos, N.J.; Leong, C.; Kular, J.; Liu, J.; Feng, X.; Zheng, M.H.; et al. Myocyte enhancer factor 2 and microphthalmia-associated transcription factor cooperate with NFATc1 to transactivate the V-ATPase d2 promoter during RANKL-induced osteoclastogenesis. J. Biol. Chem. 2009, 284, 14667–14676. [Google Scholar] [CrossRef] [Green Version]
- Pang, M.; Rodríguez-Gonzalez, M.; Hernandez, M.; Recinos, C.C.; Seldeen, K.L.; Troen, B.R. AP-1 and Mitf interact with NFATc1 to stimulate cathepsin K promoter activity in osteoclast precursors. J. Cell. Biochem. 2019, 120, 12382–12392. [Google Scholar] [CrossRef]
- Khosla, S.; Oursler, M.J.; Monroe, D.G. Estrogen and the skeleton. Trends Endocrinol. Metab. 2012, 23, 576–581. [Google Scholar] [CrossRef] [Green Version]
- Väänänen, H.K.; Härkönen, P.L. Estrogen and bone metabolism. Maturitas 1996, 23, S65–S69. [Google Scholar] [CrossRef]
- Nakamura, T.; Imai, Y.; Matsumoto, T.; Sato, S.; Takeuchi, K.; Igarashi, K.; Harada, Y.; Azuma, Y.; Krust, A.; Yamamoto, Y.; et al. Estrogen prevents bone loss via estrogen receptor alpha and induction of Fas ligand in osteoclasts. Cell 2007, 130, 811–823. [Google Scholar] [CrossRef]
- Krum, S.A.; Chang, J.; Miranda-Carboni, G.; Wang, C.Y. Novel functions for NFkappaB: Inhibition of bone formation. Nat. Rev. Rheumatol. 2010, 6, 607–611. [Google Scholar] [CrossRef]
- Kameda, T.; Mano, H.; Yuasa, T.; Mori, Y.; Miyazawa, K.; Shiokawa, M.; Nakamaru, Y.; Hiroi, E.; Hiura, K.; Kameda, A.; et al. Estrogen inhibits bone resorption by directly inducing apoptosis of the bone-resorbing osteoclasts. J. Exp. Med. 1997, 186, 489–495. [Google Scholar] [CrossRef] [Green Version]
- Bord, S.; Ireland, D.C.; Beavan, S.R.; Compston, J.E. The effects of estrogen on osteoprotegerin, RANKL, and estrogen receptor expression in human osteoblasts. Bone 2003, 32, 136–141. [Google Scholar] [CrossRef]
- Silva, B.; Costa, A.; Cusano, N.; Kousteni, S.; Bilezikian, J. Catabolic and anabolic actions of parathyroid hormone on the skeleton. J. Endocrinol. Invest. 2011, 34, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.K.; Mohamad, N.-V.; Giaze, T.R.; Chin, K.-Y.; Mohamed, N.; Ima-Nirwana, S. Prostate Cancer and Bone Metastases: The Underlying Mechanisms. Int. J. Mol. Sci. 2019, 20, 2587. [Google Scholar] [CrossRef] [Green Version]
- Martin, T.J.; Johnson, R.W. Multiple actions of PTHrP in breast cancer bone metastasis. Br. J. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Sun, P.; Wang, M.; Yin, G.Y. Endogenous parathyroid hormone (PTH) signals through osteoblasts via RANKL during fracture healing to affect osteoclasts. Biochem. Biophys. Res. Commun. 2020, 525, 850–856. [Google Scholar] [CrossRef]
- Liu, S.; Zhu, W.; Li, S.; Ma, J.; Zhang, H.; Li, Z.; Zhang, L.; Zhang, B.; Li, Z.; Liang, X.; et al. Bovine parathyroid hormone enhances osteoclast bone resorption by modulating V-ATPase through PTH1R. Int. J. Mol. Med. 2016, 37, 284–292. [Google Scholar] [CrossRef] [Green Version]
- Copp, D.H.; Cameron, E.C.; Cheney, B.A.; Davidson, A.G.; Henze, K.G. Evidence for calcitonin--a new hormone from the parathyroid that lowers blood calcium. Endocrinology 1962, 70, 638–649. [Google Scholar] [CrossRef]
- Nicholson, G.; Moseley, J.; Sexton, P.; Mendelsohn, F.; Martin, T. Abundant calcitonin receptors in isolated rat osteoclasts. Biochemical and autoradiographic characterization. J. Clin. Invest. 1986, 78, 355–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okumura, S.; Mizoguchi, T.; Sato, N.; Yamaki, M.; Kobayashi, Y.; Yamauchi, H.; Ozawa, H.; Udagawa, N.; Takahashi, N. Coordination of microtubules and the actin cytoskeleton is important in osteoclast function, but calcitonin disrupts sealing zones without affecting microtubule networks. Bone 2006, 39, 684–693. [Google Scholar] [CrossRef] [PubMed]
- Zaidi, M.; Datta, H.K.; Moonga, B.S.; MacIntyre, I. Evidence that the action of calcitonin on rat osteoclasts is mediated by two G proteins acting via separate post-receptor pathways. J. Endocrinol. 1990, 126, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Keller, J.; Catala-Lehnen, P.; Huebner, A.K.; Jeschke, A.; Heckt, T.; Lueth, A.; Krause, M.; Koehne, T.; Albers, J.; Schulze, J.; et al. Calcitonin controls bone formation by inhibiting the release of sphingosine 1-phosphate from osteoclasts. Nat. Commun. 2014, 5, 5215. [Google Scholar] [CrossRef]
- Blair, H.C.; Teitelbaum, S.L.; Ghiselli, R.; Gluck, S. Osteoclastic bone resorption by a polarized vacuolar proton pump. Science 1989, 245, 855–857. [Google Scholar] [CrossRef]
- Futai, M.; Sun-Wada, G.-H.; Wada, Y.; Matsumoto, N.; Nakanishi-Matsui, M. Vacuolar-type ATPase: A proton pump to lysosomal trafficking. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2019, 95, 261–277. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Yang, S.; Zhang, L.; Yang, T. V-ATPases and osteoclasts: Ambiguous future of V-ATPases inhibitors in osteoporosis. Theranostics 2018, 8, 5379–5399. [Google Scholar] [CrossRef]
- Wang, R.; Long, T.; Hassan, A.; Wang, J.; Sun, Y.; Xie, X.S.; Li, X. Cryo-EM structures of intact V-ATPase from bovine brain. Nat. Commun. 2020, 11, 3921. [Google Scholar] [CrossRef]
- Abbas, Y.M.; Wu, D.; Bueler, S.A.; Robinson, C.V.; Rubinstein, J.L. Structure of V-ATPase from the mammalian brain. Science 2020, 367, 1240–1246. [Google Scholar] [CrossRef]
- Qin, A.; Cheng, T.S.; Pavlos, N.J.; Lin, Z.; Dai, K.R.; Zheng, M.H. V-ATPases in osteoclasts: Structure, function and potential inhibitors of bone resorption. Int. J. Biochem. Cell Biol. 2012, 44, 1422–1435. [Google Scholar] [CrossRef]
- Gao, Y.; Patil, S.; Qian, A. The Role of MicroRNAs in Bone Metabolism and Disease. Int. J. Mol. Sci. 2020, 21, 6081. [Google Scholar] [CrossRef]
- Ghildiyal, M.; Zamore, P.D. Small silencing RNAs: An expanding universe. Nat. Rev. Genet. 2009, 10, 94–108. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Lian, J.X.; Meng, S. MiR-125a-5p promotes osteoclastogenesis by targeting TNFRSF1B. Cell. Mol. Biol. Lett. 2019, 24, 23. [Google Scholar] [CrossRef] [Green Version]
- Cong, F.; Wu, N.; Tian, X.; Fan, J.; Liu, J.; Song, T.; Fu, H. MicroRNA-34c promotes osteoclast differentiation through targeting LGR4. Gene 2017, 610, 1–8. [Google Scholar] [CrossRef]
- Sul, O.J.; Rajasekaran, M.; Park, H.J.; Suh, J.H.; Choi, H.S. MicroRNA-29b Enhances Osteoclast Survival by Targeting BCL-2-Modifying Factor after Lipopolysaccharide Stimulation. Oxid. Med. Cell Longev. 2019, 2019, 6018180. [Google Scholar] [CrossRef]
- Li, M.; Luo, R.; Yang, W.; Zhou, Z.; Li, C. miR-363-3p is activated by MYB and regulates osteoporosis pathogenesis via PTEN/PI3K/AKT signaling pathway. In Vitro Cell Dev. Biol. Anim. 2019, 55, 376–386. [Google Scholar] [CrossRef]
- Yin, R.; Jiang, J.; Deng, H.; Wang, Z.; Gu, R.; Wang, F. miR-140-3p aggregates osteoporosis by targeting PTEN and activating PTEN/PI3K/AKT signaling pathway. Hum. Cell 2020, 33, 569–581. [Google Scholar] [CrossRef]
- Zhao, C.; Sun, W.; Zhang, P.; Ling, S.; Li, Y.; Zhao, D.; Peng, J.; Wang, A.; Li, Q.; Song, J.; et al. miR-214 promotes osteoclastogenesis by targeting Pten/PI3k/Akt pathway. RNA Biol. 2015, 12, 343–353. [Google Scholar] [CrossRef]
- Lou, Z.; Peng, Z.; Wang, B.; Li, X.; Li, X.; Zhang, X. miR-142-5p promotes the osteoclast differentiation of bone marrow-derived macrophages via PTEN/PI3K/AKT/FoxO1 pathway. J. Bone Miner. Metab. 2019, 37, 815–824. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, J.; Shao, H.; Liu, J.; Jin, M.; Chen, J.; Huang, Y. miRNA-340 inhibits osteoclast differentiation via repression of MITF. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef]
- Mizoguchi, F.; Murakami, Y.; Saito, T.; Miyasaka, N.; Kohsaka, H. miR-31 controls osteoclast formation and bone resorption by targeting RhoA. Arthritis Res. Ther. 2013, 15, R102. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Liu, J.; Guo, B.; Liang, C.; Dang, L.; Lu, C.; He, X.; Cheung, H.Y.; Xu, L.; Lu, C.; et al. Osteoclast-derived exosomal miR-214-3p inhibits osteoblastic bone formation. Nat. Commun. 2016, 7, 10872. [Google Scholar] [CrossRef]
- Patil, S.; Dang, K.; Zhao, X.; Gao, Y.; Qian, A. Role of LncRNAs and CircRNAs in Bone Metabolism and Osteoporosis. Front. Genet. 2020, 11. [Google Scholar] [CrossRef]
- Liu, C.; Cao, Z.; Bai, Y.; Dou, C.; Gong, X.; Liang, M.; Dong, R.; Quan, H.; Li, J.; Dai, J. LncRNA AK077216 promotes RANKL-induced osteoclastogenesis and bone resorption via NFATc1 by inhibition of NIP45. J. Cell Physiol. 2019, 234, 1606–1617. [Google Scholar] [CrossRef]
- Ling, L.; Hu, H.L.; Liu, K.Y.; Ram, Y.I.; Gao, J.L.; Cao, Y.M. Long noncoding RNA MIRG induces osteoclastogenesis and bone resorption in osteoporosis through negative regulation of miR-1897. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10195–10203. [Google Scholar] [CrossRef]
- Lee, C.P.; Huang, Y.N.; Nithiyanantham, S.; Huang, C.M.; Ko, Y.C. LncRNA-Jak3:Jak3 coexpressed pattern regulates monosodium urate crystal-induced osteoclast differentiation through Nfatc1/Ctsk expression. Environ. Toxicol. 2019, 34, 179–187. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, T.B.; Liu, L.; Cui, Z.Q. LncRNA LINC00311 Promotes the Proliferation and Differentiation of Osteoclasts in Osteoporotic Rats Through the Notch Signaling Pathway by Targeting DLL3. Cell Physiol. Biochem. 2018, 47, 2291–2306. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhu, H.M.; Xu, H.D.; Zhang, B.; Huang, S.M. CRNDE impacts the proliferation of osteoclast by estrogen deficiency in postmenopausal osteoporosis. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5815–5821. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Fu, S.; Sun, D.; Xing, J.; Hou, T.; Wu, X. EPC-derived exosomes promote osteoclastogenesis through Lnc RNA-MALAT 1. J. Cell. Mol. Med. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, N.; Lu, Y.; Wang, Y.; Ren, X.; Han, J. Circular RNAs and hereditary bone diseases. Intract. Rare Dis. Res. 2018, 7, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Ouyang, Z.; Shen, Y.; Liu, B.; Zhang, Q.; Wan, L.; Yin, Z.; Zhu, W.; Li, S.; Peng, D. CircRNA_28313/miR-195a/CSF1 axis modulates osteoclast differentiation to affect OVX-induced bone absorption in mice. RNA Biol. 2019, 16, 1249–1262. [Google Scholar] [CrossRef]
- Miao, F.; Yin, B.H.; Zhang, X.; Xue, D.D.; Ma, C. CircRNA_009934 induces osteoclast bone resorption via silencing miR-5107. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 7580–7588. [Google Scholar] [CrossRef]
- Liu, S.; Wang, C.; Bai, J.; Li, X.; Yuan, J.; Shi, Z.; Mao, N. Involvement of circRNA_0007059 in the regulation of postmenopausal osteoporosis by promoting the microRNA-378/BMP-2 axis. Cell Biol. Int. 2021, 45, 447–455. [Google Scholar] [CrossRef]
- Liu, Z.; Li, C.; Huang, P.; Hu, F.; Jiang, M.; Xu, X.; Li, B.; Deng, L.; Ye, T.; Guo, L. CircHmbox1 Targeting miRNA-1247-5p Is Involved in the Regulation of Bone Metabolism by TNF-α in Postmenopausal Osteoporosis. Front. Cell. Dev. Biol. 2020, 8, 594785. [Google Scholar] [CrossRef]
- Soysa, N.S.; Alles, N. Osteoclast function and bone-resorbing activity: An overview. Biochem. Biophys. Res. Commun. 2016, 476, 115–120. [Google Scholar] [CrossRef]
- Vaananen, H.K.; Zhao, H.; Mulari, M.; Halleen, J.M. The cell biology of osteoclast function. J. Cell Sci. 2000, 113, 377. [Google Scholar] [CrossRef]
- Zhao, H.; Ito, Y.; Chappel, J.; Andrews, N.W.; Teitelbaum, S.L.; Ross, F.P. Synaptotagmin VII Regulates Bone Remodeling by Modulating Osteoclast and Osteoblast Secretion. Dev. Cell 2008, 14, 914–925. [Google Scholar] [CrossRef] [Green Version]
- Chellaiah, M.A. Regulation of Actin Ring Formation by Rho GTPases in Osteoclasts*. J. Biol. Chem. 2005, 280, 32930–32943. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, M.; Blair, H.C.; Moonga, B.S.; Abe, E.; Huang, C.L.H. Osteoclastogenesis, Bone Resorption, and Osteoclast-Based Therapeutics. J. Bone Miner. Res. 2003, 18, 599–609. [Google Scholar] [CrossRef]
- Shalev, M.; Elson, A. The roles of protein tyrosine phosphatases in bone-resorbing osteoclasts. Biochim. Biophys. Acta Mol. Cell. Res. 2019, 1866, 114–123. [Google Scholar] [CrossRef]
- Sims, N.A.; Martin, T.J. Coupling the activities of bone formation and resorption: A multitude of signals within the basic multicellular unit. Bonekey Rep. 2014, 3, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sims, N.A.; Martin, T.J. Osteoclasts Provide Coupling Signals to Osteoblast Lineage Cells Through Multiple Mechanisms. Annu. Rev. Physiol. 2020, 82, 507–529. [Google Scholar] [CrossRef] [PubMed]
- McNamara, L.M. Perspective on post-menopausal osteoporosis: Establishing an interdisciplinary understanding of the sequence of events from the molecular level to whole bone fractures. J. R. Soc. Interface 2010, 7, 353–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, U.H. Bone remodeling in post-menopausal osteoporosis. J. Dent. Res. 2006, 85, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Stein, E.; Shane, E. Secondary osteoporosis. Endocrinol. Metab. Clin. N. Am. 2003, 32, 115–134. [Google Scholar] [CrossRef]
- Armas, L.A.; Recker, R.R. Pathophysiology of osteoporosis: New mechanistic insights. Endocrinol. Metab. Clin. N. Am. 2012, 41, 475–486. [Google Scholar] [CrossRef]
- Johannesdottir, F.; Thrall, E.; Muller, J.; Keaveny, T.M.; Kopperdahl, D.L.; Bouxsein, M.L. Comparison of non-invasive assessments of strength of the proximal femur. Bone 2017, 105, 93–102. [Google Scholar] [CrossRef]
- Kanis, J.A.; Cooper, C.; Rizzoli, R.; Reginster, J.Y. European guidance for the diagnosis and management of osteoporosis in postmenopausal women. Osteoporos. Int. 2019, 30, 3–44. [Google Scholar] [CrossRef] [Green Version]
- Stapleton, M.; Sawamoto, K.; Alméciga-Díaz, C.J.; Mackenzie, W.G.; Mason, R.W.; Orii, T.; Tomatsu, S. Development of Bone Targeting Drugs. Int. J. Mol. Sci. 2017, 18, 1345. [Google Scholar] [CrossRef]
- Anagnostis, P.; Paschou, S.A.; Mintziori, G.; Ceausu, I.; Depypere, H.; Lambrinoudaki, I.; Mueck, A.; Perez-Lopez, F.R.; Rees, M.; Senturk, L.M.; et al. Drug holidays from bisphosphonates and denosumab in postmenopausal osteoporosis: EMAS position statement. Maturitas 2017, 101, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Stone, J.A.; McCrea, J.B.; Witter, R.; Zajic, S.; Stoch, S.A. Clinical and translational pharmacology of the cathepsin K inhibitor odanacatib studied for osteoporosis. Br. J. Clin. Pharmacol. 2019, 85, 1072–1083. [Google Scholar] [CrossRef]
- Boggild, M.K.; Gajic-Veljanoski, O.; McDonald-Blumer, H.; Ridout, R.; Tile, L.; Josse, R.; Cheung, A.M. Odanacatib for the treatment of osteoporosis. Expert Opin. Pharmacother. 2015, 16, 1717–1726. [Google Scholar] [CrossRef]
- McClung, M.R.; O’Donoghue, M.L.; Papapoulos, S.E.; Bone, H.; Langdahl, B.; Saag, K.G.; Reid, I.R.; Kiel, D.P.; Cavallari, I.; Bonaca, M.P.; et al. Odanacatib for the treatment of postmenopausal osteoporosis: Results of the LOFT multicentre, randomised, double-blind, placebo-controlled trial and LOFT Extension study. Lancet Diabetes Endocrinol. 2019, 7, 899–911. [Google Scholar] [CrossRef]
- Deeks, E.D. Denosumab: A Review in Postmenopausal Osteoporosis. Drugs Aging 2018, 35, 163–173. [Google Scholar] [CrossRef]
- Ran, S.Y.; Yu, Q.; Chen, Y.; Lin, S.Q. Prevention of postmenopausal osteoporosis in Chinese women: A 5-year, double-blind, randomized, parallel placebo-controlled study. Climact. J. Int. Menopause Soc. 2017, 20, 391–396. [Google Scholar] [CrossRef]
- Lufkin, E.G.; Wahner, H.W.; O’Fallon, W.M.; Hodgson, S.F.; Kotowicz, M.A.; Lane, A.W.; Judd, H.L.; Caplan, R.H.; Riggs, B.L. Treatment of postmenopausal osteoporosis with transdermal estrogen. Ann. Intern. Med. 1992, 117, 1–9. [Google Scholar] [CrossRef]
- Gambacciani, M. Selective estrogen modulators in menopause. Minerva Ginecol. 2013, 65, 621–630. [Google Scholar]
- Gambacciani, M.; Levancini, M. Management of postmenopausal osteoporosis and the prevention of fractures. Panminerva Med. 2014, 56, 115–131. [Google Scholar]
- Yamamoto, Y.; Yamamoto, Y.; Udagawa, N.; Okumura, S.; Mizoguchi, T.; Take, I.; Yamauchi, H.; Yamauchi, H.; Noguchi, T.; Takahashi, N. Effects of calcitonin on the function of human osteoclast-like cells formed from CD14-positive monocytes. Cell. Mol. Biol. 2006, 52, 25–31. [Google Scholar]
- Zhou, Z.; Wang, N.; Ding, C.; Zhou, X.; Zhou, J. Postmenopausal osteoporosis treated with acupoint injection of salmon calcitonin:a randomized controlled trial. Zhongguo Zhen Jiu Chin. Acupunct. Moxib. 2016, 36, 705–708. [Google Scholar] [CrossRef]
- Sugimoto, T.; Shiraki, M.; Nakano, T.; Kishimoto, H.; Ito, M.; Fukunaga, M.; Sone, T.; Hagino, H.; Miki, T.; Nishizawa, Y.; et al. A randomized, double-blind, placebo-controlled study of once weekly elcatonin in primary postmenopausal osteoporosis. Curr. Med. Res. Opin. 2019, 35, 447–454. [Google Scholar] [CrossRef]
- Przedlacki, J. Strontium ranelate in post-menopausal osteoporosis. Endokrynol. Polska 2011, 62, 65–72. [Google Scholar]
- Hamdy, N.A.T. Strontium ranelate improves bone microarchitecture in osteoporosis. Rheumatology 2009, 48, iv9–iv13. [Google Scholar] [CrossRef] [Green Version]
- Visentin, L.; Dodds, R.A.; Valente, M.; Misiano, P.; Bradbeer, J.N.; Oneta, S.; Liang, X.; Gowen, M.; Farina, C. A selective inhibitor of the osteoclastic V-H(+)-ATPase prevents bone loss in both thyroparathyroidectomized and ovariectomized rats. J. Clin. Investig. 2000, 106, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Beth-Tasdogan, N.H.; Mayer, B.; Hussein, H.; Zolk, O. Interventions for managing medication-related osteonecrosis of the jaw. Cochrane Database Syst. Rev. 2017, 10. [Google Scholar] [CrossRef]
- Zhao, X.; Patil, S.; Qian, A.; Zhao, C. Bioactive Compounds of Polygonatum Sibiricum - Therapeutic Effect and Biological Activity. Endocr. Metab. Immune Disord. Drug Targets 2021. [Google Scholar] [CrossRef]
- Cao, H.; Liu, J.; Lewith, G.T. Traditional Chinese Medicine for treatment of fibromyalgia: A systematic review of randomized controlled trials. J. Altern. Complement. Med. 2010, 16, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Xiao, E.; Luo, L. Alternative Therapies for Diabetes: A Comparison of Western and Traditional Chinese Medicine (TCM) Approaches. Curr. Diabetes Rev. 2018, 14, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Zhu, J.; Wang, Y.; Zhang, N.; Gober, H.J.; Qiu, X.; Li, D.; Wang, L. Chinese single herbs and active ingredients for postmenopausal osteoporosis: From preclinical evidence to action mechanism. Biosci. Trends 2017, 11, 496–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Dang, L.; Li, D.; Liang, C.; He, X.; Wu, H.; Qian, A.; Yang, Z.; Au, D.W.; Chiang, M.W.; et al. A delivery system specifically approaching bone resorption surfaces to facilitate therapeutic modulation of microRNAs in osteoclasts. Biomaterials 2015, 52, 148–160. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Yang, L.; Zhang, S.; Liu, J.; Sun, Y.; Wang, X. A bone-resorption surface-targeting nanoparticle to deliver anti-miR214 for osteoporosis therapy. Int. J. Nanomed. 2017, 12, 7469–7482. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.S.; Xie, J.; Chaugule, S.; Wang, D.; Kim, J.M.; Kim, J.; Tai, P.W.L.; Seo, S.K.; Gravallese, E.; Gao, G.; et al. Bone-Targeting AAV-Mediated Gene Silencing in Osteoclasts for Osteoporosis Therapy. Mol. Ther. Methods Clin. Dev. 2020, 17, 922–935. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| MiRNA | Role | Target | Reference |
|---|---|---|---|
| MiR-125a-5p | Promotes osteoclastogenesis | TNFRSF1B | [83] |
| MiR-34c | Promotes osteoclast differentiation | LGR4 | [84] |
| MiR-29b | Increases osteoclasts number | BMF | [85] |
| MiR-363-3p | Promote osteoclastogenesis | PTEN | [86] |
| MiR-140-3p | [87] | ||
| MiR-214 | [88] | ||
| MiR-142-5p | [89] | ||
| MiR-340 | Inhibits osteoclast differentiation | MITF | [90] |
| MiR-31 | Regulates osteoclast cytoskeleton organization | RhoA | [91] |
| MiR-214-3p | Reduces bone formation | ND | [92] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Patil, S.; Xu, F.; Lin, X.; Qian, A. Role of Biomolecules in Osteoclasts and Their Therapeutic Potential for Osteoporosis. Biomolecules 2021, 11, 747. https://doi.org/10.3390/biom11050747
Zhao X, Patil S, Xu F, Lin X, Qian A. Role of Biomolecules in Osteoclasts and Their Therapeutic Potential for Osteoporosis. Biomolecules. 2021; 11(5):747. https://doi.org/10.3390/biom11050747
Chicago/Turabian StyleZhao, Xin, Suryaji Patil, Fang Xu, Xiao Lin, and Airong Qian. 2021. "Role of Biomolecules in Osteoclasts and Their Therapeutic Potential for Osteoporosis" Biomolecules 11, no. 5: 747. https://doi.org/10.3390/biom11050747
APA StyleZhao, X., Patil, S., Xu, F., Lin, X., & Qian, A. (2021). Role of Biomolecules in Osteoclasts and Their Therapeutic Potential for Osteoporosis. Biomolecules, 11(5), 747. https://doi.org/10.3390/biom11050747