
Investigation of Active Anti-Inflammatory Constituents of Essential Oil from Pinus koraiensis (Sieb. et Zucc.) Wood in LPS-Stimulated RBL-2H3 Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material
2.3. Extraction of Essential Oil and Its Fractioanl Distillation
2.4. GC–MS Analysis
2.5. Cell Culture
2.6. Cell Cytotoxicity
2.7. Determination of the Levels of Cytokines IL-4 and IL-13 via Quantitative Real-Time PCR
2.8. β-Hexosaminidase Secretion Assay
2.9. Statistical Analysis
3. Results
3.1. Yield and Chemical Composition
3.2. Cell Cytotoxicity
3.3. Effects of the Oil Fractions on the Expression of IL-4 and IL-13
3.4. Effects of the Oil Fractions on β-Hexosaminidase Release
3.5. Effects of Single Compounds on Cell Viability
3.6. Effects of Single Compounds on the Expression of IL-4 and IL-13
3.7. Effects of Active Compounds on β-Hexosaminidase Release
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vidakovic, M. Conifers: Morphology and variation. Tree Physiol. 1993, 12, 319. [Google Scholar] [CrossRef]
- Wolff, R.L.; Bayard, C.C. Fatty acid composition of some pine seed oils. J. Am. Oil Chem. Soc. 1995, 72, 1043–1046. [Google Scholar] [CrossRef]
- Asset, G.; Staels, B.; Wolff, R.L.; Baugé, E.; Madj, Z.; Fruchart, J.-C.; Dallongeville, J. Effects of Pinus pinaster and Pinus koraiensis seed oil supplementation on lipoprotein metabolism in the rat. Lipids 1999, 34, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Li, Q.; Li, J.; Zhang, T.; Han, Z.; Gao, D.; Zheng, F. Antitumor Activity of the Procyanidins from Pinus koraiensis Bark on mice bearing U14 cervical cancer. Yakugaku Zasshi 2007, 127, 1145–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-K.; Im, S.-B.; Choi, S.-H.; Lee, J.-S.; Roh Mark, S.; Lim, Y.-P. Construction of a full-length cDNA library from Pinus koraiensis and analysis of EST dataset. Korean, J. Agric. Sci. 2011, 38, 11–16. [Google Scholar] [CrossRef]
- Pérez, G.S.; Zavala, S.M.; Arias, G.L.; Ramos, L.M. Anti-inflammatory activity of some essential oils. J. Essent. Oil Res. 2011, 23, 38–44. [Google Scholar] [CrossRef]
- Dinarello, C.A. Proinflammatory and anti-inflammatory cytokines as mediators in the pathogenesis of septic shock. Chest 1997, 112, 321S–329S. [Google Scholar] [CrossRef] [Green Version]
- Marie, C.; Pitton, C.; Fitting, C.; Cavaillon, J.M. Regulation by anti-inflammatory cytokines (IL-4, IL-10, IL-13, TGFβ)of interleukin-8 production by LPS- and/ or TNFα-activated human polymorphonuclear cells. Mediat. Inflamm. 1996, 5, 334–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soyer, O.U.; Akdis, M.; Akdis, C.A. Mechanisms of subcutaneous allergen immunotherapy. Immunol. Allergy Clin. N. Am. 2011, 31, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Catley, M.C.; Coote, J.; Bari, M.; Tomlinson, K.L. Monoclonal antibodies for the treatment of asthma. Pharmacol. Ther. 2011, 132, 333–351. [Google Scholar] [CrossRef] [PubMed]
- Grünig, G.; Warnock, M.; Wakil, A.E.; Venkayya, R.; Brombacher, F.; Rennick, D.M.; Sheppard, D.; Mohrs, M.; Donaldson, D.D.; Locksley, R.M.; et al. Requirement for IL-13 Independently of IL-4 in Experimental Asthma. Science 1998, 282, 2261–2263. [Google Scholar] [CrossRef] [Green Version]
- Wills-Karp, M.; Luyimbazi, J.; Xu, X.; Schofield, B.; Neben, T.Y.; Karp, C.L.; Donaldson, D.D. Interleukin-13: Central mediator of allergic asthma. Science 1998, 282, 2258–2261. [Google Scholar] [CrossRef] [Green Version]
- Ogirala, R.G.; Aldrich, T.K.; Prezant, D.J.; Sinnett, M.J.; Enden, J.B.; Williams, M.H. High-Dose intramuscular triamcinolone in severe, chronic, life-threatening asthma. N. Engl. J. Med. 1991, 324, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Ballabeni, V.; Tognolini, M.; Chiavarini, M.; Impicciatore, M.; Bruni, R.; Bianchi, A.; Barocelli, E. Novel antiplatelet and antithrombotic activities of essential oil from Lavandula hybrida Reverchon “grosso”. Phytomedicine 2004, 11, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Tu, P.T.B.; Tawata, S. Anti-Oxidant, Anti-Aging, and Anti-Melanogenic Properties of the Essential Oils from Two Varieties of Alpinia zerumbet. Molecules 2015, 20, 16723–16740. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Choi, W.S.; Kim, J.W.; Lee, S.S.; Park, M.J. Anti-Inflammatory Effect of Essential Oils Extracted from Wood of Four Coniferous Tree Species. J. Korean Wood Sci. Technol. 2019, 47, 674–691. [Google Scholar] [CrossRef]
- Nam, H.; Choi, J.; Capareda, S. Comparative study of vacuum and fractional distillation using pyrolytic microalgae (Nannochloropsis oculata) bio-oil. Algal Res. 2016, 17, 87–96. [Google Scholar] [CrossRef]
- Ham, Y.; Yang, J.; Choi, W.; Ahn, B.; Park, M. Antibacterial activity of essential oils from Pinaceae leaves against Fish pathogens. J. Korean Wood Sci. Technol. 2020, 48, 527–547. [Google Scholar] [CrossRef]
- Yang, J.; Ahn, C.; Jeung, E.; Choi, W.; Kim, J.; Park, M. Anti-inflammatory activity of essential oil extracted from Chamaecyparis obtuse (Sieb. et Zucc.) wood. Kor. J. Pharmacogn. 2017, 48, 18–24. [Google Scholar]
- Jung, E.-M.; Kim, J.-W.; Park, M.-J.; Lee, S.-S.; Choi, D.-H.; Lee, J.; Jeung, E.-B. Inhibitory Effect of Extracts from Rhododendron Brachycarpum and Abies Koreana, E.H. Wilson on Degranulation and Cytokine Expression in RBL-2H3 Cells. J. Korean Wood Sci. Technol. 2013, 41, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Niss, F.; Rosenmai, A.K.; Mandava, G.; Örn, S.; Oskarsson, A.; Lundqvist, J. Toxicity bioassays with concentrated cell culture media—a methodology to overcome the chemical loss by conventional preparation of water samples. Environ. Sci. Pollut. Res. 2018, 25, 12183–12188. [Google Scholar] [CrossRef] [Green Version]
- Pawankar, R.; Mori, S.; Ozu, C.; Kimura, S. Overview on the pathomechanisms of allergic rhinitis. Asia Pac. Allergy 2011, 1, 157–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juckmeta, T.; Thongdeeying, P.; Itharat, A. Inhibitory Effect onβ-Hexosaminidase Release from RBL-2H3 Cells of Extracts and Some Pure Constituents of Benchalokawichian, a Thai Herbal Remedy, Used for Allergic Disorders. Evid.-Based Complement. Altern. Med. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jüni, P.; Reichenbach, S.; Egger, M. COX 2 inhibitors, traditional NSAIDs, and the heart. BMJ 2005, 330, 1342–1343. [Google Scholar] [CrossRef]
- Opal, S.M.; DePalo, V.A. Anti-Inflammatory Cytokines. Chest 2000, 117, 1162–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callard, R.E.; Matthews, D.J.; Hibbert, L. IL-4 and IL-13 receptors: Are they one and the same? Immunol. Today 1996, 17, 108–110. [Google Scholar] [CrossRef]
- Kim, S.H.; Lee, S.Y.; Cho, S.M.; Hong, C.Y.; Park, M.J.; Choi, I.G. Evaluation on anti-fungal activity and synergy effects of essential oil and their constituents from abies holophylla. J. Korean Wood Sci. Technol. 2016, 44, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Kim, Y.Y.; Choi, Y.A.; Baek, M.C.; Lee, B.; Park, P.H.; Shin, T.Y.; Kwon, T.K.; Khang, D.; Kim, S.H. Elaeocarpusin inhibits mast cell-mediated allergic inflammation. Front. Pharmacol. 2018, 9, 591. [Google Scholar] [CrossRef]
- Fang, H.; Pengal, R.A.; Cao, X.; Ganesan, L.P.; Wewers, M.D.; Marsh, C.B.; Tridandapani, S. Lipopolysaccharide-induced macrophage inflammatory response is regulated by SHIP. J. Immunol. 2004, 173, 360–366. [Google Scholar] [CrossRef] [Green Version]
- Seemann, S.; Zohles, F.; Lupp, A. Comprehensive comparison of three different animal models for systemic inflammation. J. Biomed. Sci. 2017, 24, 1–17. [Google Scholar] [CrossRef]
- Khaleel, C.; Tabanca, N.; Buchbauer, G. α-Terpineol, a natural monoterpene: A review of its biological properties. Open Chem. 2018, 16, 349–361. [Google Scholar] [CrossRef]
- Drosopoulou, E.; Vlastos, D.; Efthimiou, I.; Kyrizaki, P.; Tsamadou, S.; Anagnostopoulou, M.; Kofidou, D.; Gavriilidis, M.; Mademtzoglou, D.; Mavragani-Tsipidou, P. In vitro and in vivo evaluation of the genotoxic and antigenotoxic potential of the major Chios mastic water constituents. Sci. Rep. 2018, 8, 12200. [Google Scholar] [CrossRef] [PubMed]
- Sabino, C.K.B.; Ferreira-Filho, E.S.; Mendes, M.B.; Da Silva-Filho, J.C.; Ponte, M.P.T.R.; Moura, L.H.P.; Oliveira, E.C.A.; Quintans-Junior, L.J.; Dos Santos, M.R.V.; Oliveira, R.D.C.M.; et al. Cardiovascular effects induced by α-terpineol in hypertensive rats. Flavour Fragr. J. 2013, 28, 333–339. [Google Scholar] [CrossRef]
- Salehi, B.; Upadhyay, S.; Orhan, I.E.; Jugran, A.K.; Jayaweera, S.L.; Dias, D.A.; Sharopov, F.; Taheri, Y.; Martins, N.; Baghalpour, N.; et al. Therapeutic Potential of α- and β-Pinene: A Miracle Gift of Nature. Biomolecules 2019, 9, 738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conathy, J.M.; Owens, M.J. Stereochemistry in drug action. Prim. Care Companion J. Clin. Psychiatry 2003, 5, 70–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, K.D.; Birt, D.F. Evidence for contributions of interactions of constituents to the anti-inflammatory activity ofHypericum perforatum. Crit. Rev. Food Sci. Nutr. 2013, 54, 781–789. [Google Scholar] [CrossRef] [PubMed]
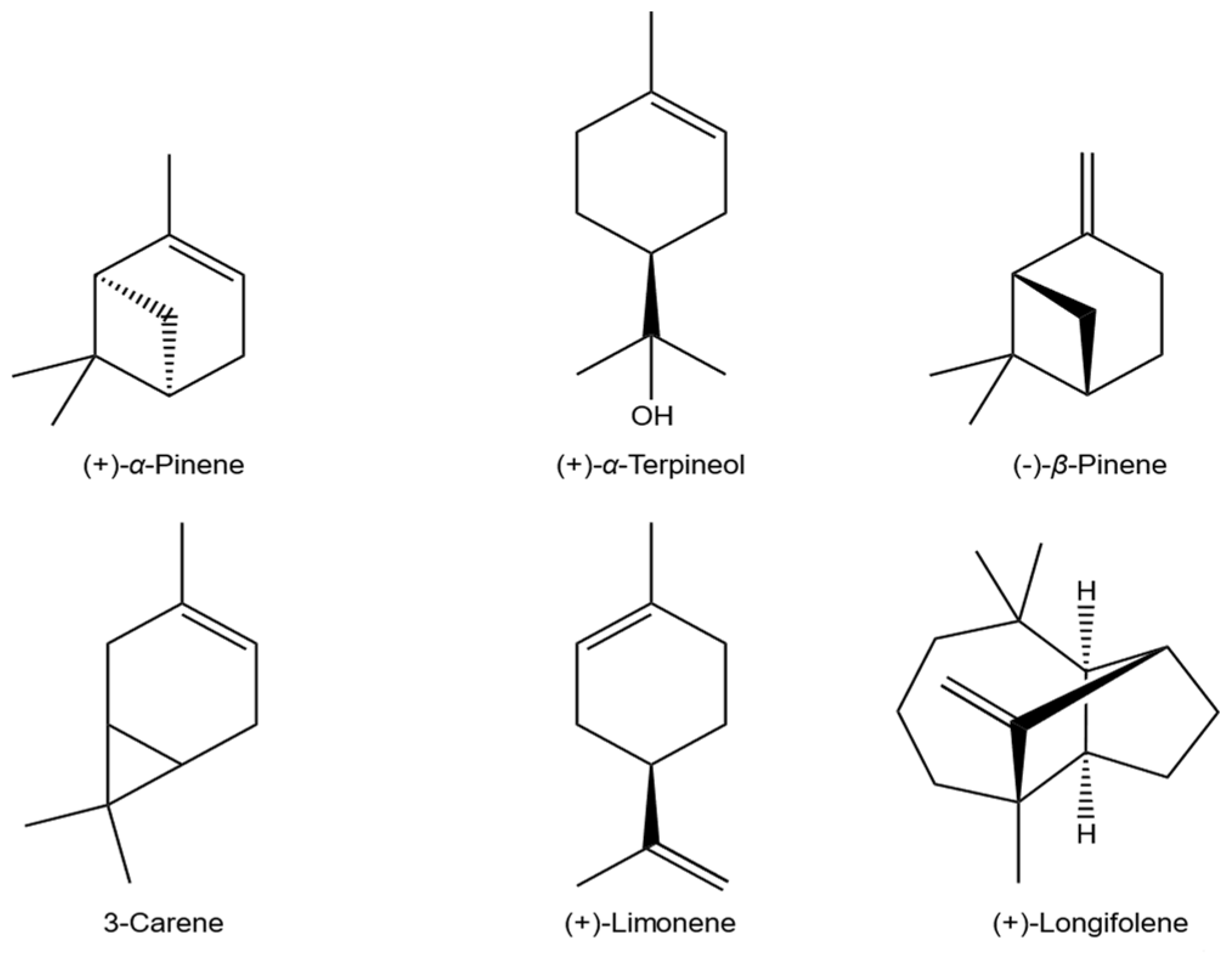







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oil Fraction | Temperature Range (°C) |
|---|---|
| A | 50–65 |
| B | 65–75 |
| C | 75–81 |
| D | 81–82 |
| E | 82–94 |
| F | Residues |
| Column | DB-5MS capillary column | ||
| (30 m × 0.25 mm; 0.25 μm; Thermo Scientific, USA) | |||
| GC oven conditions | 50 °C, hold 5 min | ||
| 10 °C/min to 65 °C, hold 30 min | |||
| 5 °C/min to 120 °C, hold 10 min | |||
| 5 °C/min to 180 °C | |||
| 5 °C/min to 210 °C, hold 10 min | |||
| 20 °C/min to 325 °C, hold 10 min | |||
| Carrier gas | He (1 mL/min, 25 psi) | ||
| Linear velocity | 19.8 cm/s | ||
| Injection mode | Split 1:20 | ||
| Injection temperature | 250 °C | ||
| MS parameters | FID parameters | ||
| MS ionization mode | EI | FID temperature | 300 °C |
| Scan time | 0.2 s | Hydrogen flow | 35.0 mL/min |
| Mass range | 35–550 amu | Air flow | 350.0 mL/min |
| Ion source temperature | 270 °C | Make up flow | 40.0 mL/min |
| Interface temperature | 250 °C | ||
| Gene | Primer Sequence (5′–3′) | Accession No. |
|---|---|---|
| IL-4 | F: TGA TGT ACC TCC GTG CTT GA R: AGG ACA TGG AAG TGC AGG AC | X16058 |
| IL-13 | F: CTG GAA TCC CTG ACC AAC AT R: CCA TAG CGG AAA AGT TGC TT | L26913 |
| GAPDH | F: CCA CAG TCC ATG CCA TCA C R: TCC ACC ACC CTG TTG CTG TA | NM_017008.4 |
| Oil | Weight of Fraction (g) | Yield (%) |
|---|---|---|
| Fraction A | 4.0 | 15.2 |
| Fraction B | 0.7 | 2.6 |
| Fraction C | 6.1 | 23.4 |
| Fraction D | 1.4 | 5.5 |
| Fraction E | 1.7 | 6.4 |
| Fraction F | 12.3 | 46.9 |
| KI a | Compound Name b | Area % | ||||||
|---|---|---|---|---|---|---|---|---|
| Wood Oil | Fraction A | Fraction B | Fraction C | Fraction D | Fraction E | Fraction F | ||
| Monoterpene hydrocarbons | ||||||||
| 913 | β-Ocimene | - | - | - | - | 0.01 | - | - |
| 919 | Tricyclene | 0.08 | 0.23 | 0.24 | 0.17 | 0.05 | - | - |
| 921 | α-Thujene | - | 0.11 | 0.12 | 0.09 | - | - | - |
| 928 | (+)-α-Pinene | 27.00 | 58.26 | 62.99 | 54.66 | 22.78 | 3.34 | - |
| 935 | (−)-β-Citronellene | - | 0.08 | 0.09 | 0.08 | 0.04 | - | - |
| 941 | (+)-Camphene | 0.14 | 0.25 | 0.29 | 0.29 | 0.20 | 0.05 | - |
| 943 | (−)-Camphene | 1.10 | 1.93 | 2.27 | 2.40 | 1.80 | 0.45 | - |
| 946 | 2,4(10)-Thujadien | 0.13 | 0.17 | 0.20 | 0.18 | 0.10 | 0.07 | - |
| 973 | (−)-β-Pinene | 11.15 | 13.19 | 16.54 | 21.95 | 29.67 | 6.65 | - |
| 986 | β-Myrcene | 0.23 | 0.15 | 0.21 | 0.26 | 0.27 | 0.06 | - |
| 1013 | α-Phellandrene | - | 0.03 | 0.05 | 0.08 | 0.16 | 0.11 | - |
| 1017 | 3-Carene | 3.05 | 2.49 | 3.32 | 5.44 | 11.67 | 4.30 | - |
| 1030 | α-Terpinene | 0.33 | 0.17 | 0.27 | 0.48 | 1.05 | 0.68 | - |
| 1040 | Cymene | 1.32 | 0.77 | 1.09 | 1.91 | 5.71 | 6.66 | - |
| 1045 | (+)-Limonene | 3.47 | 1.93 | 2.68 | 4.92 | 14.54 | 13.32 | - |
| 1047 | Sabinene | 0.16 | 0.05 | 0.08 | 0.16 | 0.40 | 0.32 | - |
| 1063 | (1S,3S)-(E)-4-Carene | - | - | - | - | 0.03 | - | - |
| 1072 | γ-Terpinene | 0.12 | 0.04 | 0.06 | 0.11 | 0.32 | 0.38 | - |
| 1092 | Terpinolene | 0.65 | 0.14 | 0.21 | 0.44 | 1.41 | 3.70 | - |
| 1109 | (+)-Verbenone | - | - | - | - | - | 0.04 | - |
| 1114 | 1,5,8-p-Menthatriene | - | - | - | - | - | 0.15 | - |
| 1158 | Camphene hydrate | 0.15 | 0.01 | 0.02 | 0.03 | 0.06 | 0.79 | 0.22 |
| 1213 | 3,6,6-Trimethylnorpinan-2-one | - | - | - | - | - | 0.08 | - |
| 1250 | Carvotanacetone | - | - | - | - | - | 0.06 | - |
| Oxygenated monoterpenes | ||||||||
| 982 | (1S,2S,3R,5S)-(+)-2,3-Pinanediol | - | - | - | - | 0.04 | - | - |
| 1027 | 1,4-Cineole | - | 0.02 | 0.03 | 0.06 | 0.14 | 0.09 | - |
| 1049 | 1,8-Cineole | - | 0.01 | 0.02 | 0.04 | 0.09 | 0.10 | - |
| 1083 | ρ-Menth-1-ene-3β,7-diol | 0.15 | 0.04 | 0.06 | 0.12 | 0.36 | 1.10 | - |
| 1101 | α-Pinene oxide | - | 0.03 | 0.02 | - | - | - | - |
| 1121 | Fenchol | 0.77 | 0.05 | 0.12 | 0.25 | 0.61 | 5.36 | 0.72 |
| 1128 | α-Campholenic aldehyde | 0.23 | 0.04 | 0.05 | 0.09 | 0.18 | 1.28 | - |
| 1142 | (−)-(E)-Pinocarveol | 0.71 | 0.03 | 0.08 | 0.15 | 0.28 | 1.92 | 0.18 |
| 1149 | (+)-Camphor | 0.25 | 0.03 | 0.04 | 0.06 | 0.12 | 1.33 | 0.54 |
| 1152 | α-Phellandren-8-ol | 0.25 | 0.01 | 0.01 | 0.02 | 0.02 | 0.06 | - |
| 1163 | (E)-Pinocamphone | 0.10 | 0.02 | 0.04 | 0.07 | 0.12 | 1.26 | 0.50 |
| 1165 | Pinocarvone | 0.13 | - | - | 0.01 | 0.01 | - | - |
| 1166 | Isoborneol | - | - | - | - | - | 0.14 | - |
| 1180 | Isopinocamphone | 0.31 | 0.01 | 0.01 | 0.02 | 0.02 | 0.30 | - |
| 1183 | Terpinen-4-ol | 1.55 | 0.07 | 0.12 | 0.20 | 0.32 | 5.51 | 3.17 |
| 1190 | ρ-Cymen-8-ol | 0.28 | - | 0.02 | 0.02 | 0.04 | 0.74 | 0.60 |
| 1200 | (+)-α-Terpineol | 7.07 | 0.23 | 0.40 | 0.66 | 0.84 | 16.69 | 16.98 |
| 1202 | (−)-Myrtenol | 0.81 | 0.02 | 0.03 | 0.06 | 0.10 | - | - |
| 1202 | Estragole | - | - | - | - | - | 1.04 | 0.47 |
| 1212 | (−)-Verbenone | 0.30 | 0.01 | 0.02 | 0.02 | 0.02 | - | - |
| 1218 | (−)-β-Fenchyl acetate | - | - | - | - | - | 0.05 | - |
| 1220 | (R)-Carveol | 0.08 | - | - | - | - | 0.19 | 0.27 |
| 1228 | Methyl thymol | - | - | - | - | - | 0.14 | - |
| 1244 | (+)-Carvone | - | - | - | - | - | 0.06 | 0.16 |
| 1286 | Bornyl acetate | 1.26 | 0.03 | 0.04 | 0.06 | 0.07 | 1.63 | 3.51 |
| 1304 | Perillyl alcohol | - | - | - | - | - | - | 0.21 |
| 1343 | Piperitenone | - | - | - | - | - | - | 0.31 |
| Sesquiterpene hydrocarbons | ||||||||
| 1349 | (−)-α-Cubebene | - | - | - | - | - | 0.06 | - |
| 1354 | α-Longipinene | 1.09 | 0.01 | 0.01 | 0.02 | 0.02 | 0.60 | 2.24 |
| 1373 | (+)-Cyclosativene | 0.18 | - | - | - | - | 0.10 | 0.64 |
| 1379 | α-Copaene | 1.34 | 0.01 | 0.01 | 0.02 | 0.02 | 0.55 | 2.35 |
| 1396 | (+)-Sativene | 0.22 | - | - | - | - | 0.08 | 0.73 |
| 1405 | (+)-Aromadendrene | - | - | - | - | - | - | 0.15 |
| 1406 | (−)-α-Gurjunene | - | - | - | - | - | - | 0.24 |
| 1408 | γ-Elemene | - | - | - | - | - | - | 0.68 |
| 1415 | Longifolene | 8.77 | 0.04 | 0.05 | 0.07 | 0.06 | 2.61 | 27.87 |
| 1423 | β-Caryophyllene | 1.08 | - | - | - | - | 0.19 | 1.82 |
| 1436 | α-Guaiene | - | - | - | - | - | - | 0.29 |
| 1452 | (E)-β-Famesene | 0.24 | - | - | - | - | - | - |
| 1457 | α-Humulene | 0.26 | - | - | - | - | - | 0.78 |
| 1475 | γ-Muurolene | - | - | - | - | - | - | 0.21 |
| 1489 | α-Ylangene | - | - | - | - | - | - | 0.38 |
| 1491 | β-Selinene | 0.07 | - | - | - | - | - | 0.20 |
| 1495 | (−)-β-Chamigrene | - | - | - | - | - | - | 0.32 |
| 1498 | α-Selinene | - | - | - | - | - | - | 0.16 |
| 1501 | α-Muurolene | 0.51 | - | - | - | - | 0.05 | 1.77 |
| 1504 | β-Himachalene | 0.07 | - | - | - | - | - | 0.30 |
| 1512 | β-Bisabolene | 0.19 | - | - | - | - | - | 0.62 |
| 1514 | (+)-Cuparene | - | - | - | - | - | - | 0.16 |
| 1526 | (+)-δ-Cadinene | 0.47 | - | - | - | - | - | 1.46 |
| 1531 | Calamenene | 0.09 | - | - | - | - | - | 0.64 |
| 1559 | α-Calacorene | - | - | - | - | - | - | 0.14 |
| 1984 | β-Elemene | - | - | - | - | - | - | 1.10 |
| Oxygenated sesquiterpenes | ||||||||
| 1578 | (E)-Nerolidol | 0.24 | - | - | - | - | - | 0.30 |
| 1593 | (−)-Caryophyllene oxide | 0.24 | - | - | - | - | - | - |
| 1598 | Viridiflorol | - | - | - | - | - | - | 0.29 |
| 1609 | Longiborneol | 0.27 | - | - | - | - | - | 1.19 |
| 1639 | Cubenol | 0.16 | - | - | - | - | - | 0.63 |
| 1659 | δ-Cadinol | 0.08 | - | - | - | - | - | 0.31 |
| 1668 | α-Cadinol | 0.08 | - | - | - | - | - | - |
| 1694 | α-Bisabolol | - | - | - | - | - | - | 0.22 |
| Diterpene hydrocarbons | ||||||||
| 1951 | Cembrene | 8.36 | - | - | - | - | - | 0.28 |
| 2036 | Manoyl oxide | 0.07 | - | - | - | - | - | - |
| 2043 | Pimara-8(14),15-diene | - | - | - | - | - | - | 0.28 |
| 2151 | Sclarene | - | - | - | - | - | - | 0.15 |
| 2293 | Isopimara-7,15-dien-3-one | 0.12 | - | - | - | - | - | 0.22 |
| Oxygenated diterpenes | ||||||||
| 2083 | Thunbergol | 2.38 | - | - | - | - | - | - |
| 2092 | Verticiol | - | - | - | - | - | - | 0.32 |
| 2199 | Sclareol | - | - | - | - | - | - | 0.32 |
| Triterpene hydrocarbons | ||||||||
| 1855 | Squalene | - | - | - | - | - | - | 0.18 |
| Monoterpene hydrocarbons | 49.09 | 80.00 | 90.74 | 93.66 | 90.30 | 41.22 | 0.22 | |
| Oxygenated monoterpenes | 14.27 | 0.65 | 1.11 | 1.91 | 3.39 | 38.99 | 27.62 | |
| Sesquiterpene hydrocarbons | 14.58 | 0.06 | 0.07 | 0.11 | 0.10 | 4.25 | 45.27 | |
| Oxygenated sesquiterpenes | 1.08 | - | - | - | - | - | 2.93 | |
| Diterpene hydrocarbons | 8.56 | - | - | - | - | - | 0.93 | |
| Oxygenated diterpenes | 2.38 | - | - | - | - | - | 0.63 | |
| Triterpene hydrocarbons | - | - | - | - | - | - | 0.18 | |
| Others | 10.03 | 19.29 | 8.08 | 4.32 | 6.21 | 15.54 | 22.20 | |
| Total identified | 99.99 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 99.98 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Choi, W.-S.; Kim, K.-J.; Eom, C.-D.; Park, M.-J. Investigation of Active Anti-Inflammatory Constituents of Essential Oil from Pinus koraiensis (Sieb. et Zucc.) Wood in LPS-Stimulated RBL-2H3 Cells. Biomolecules 2021, 11, 817. https://doi.org/10.3390/biom11060817
Yang J, Choi W-S, Kim K-J, Eom C-D, Park M-J. Investigation of Active Anti-Inflammatory Constituents of Essential Oil from Pinus koraiensis (Sieb. et Zucc.) Wood in LPS-Stimulated RBL-2H3 Cells. Biomolecules. 2021; 11(6):817. https://doi.org/10.3390/biom11060817
Chicago/Turabian StyleYang, Jiyoon, Won-Sil Choi, Ki-Joong Kim, Chang-Deuk Eom, and Mi-Jin Park. 2021. "Investigation of Active Anti-Inflammatory Constituents of Essential Oil from Pinus koraiensis (Sieb. et Zucc.) Wood in LPS-Stimulated RBL-2H3 Cells" Biomolecules 11, no. 6: 817. https://doi.org/10.3390/biom11060817