
Circulating MicroRNAs Highly Correlate to Expression of Cartilage Genes Potentially Reflecting OA Susceptibility—Towards Identification of Applicable Early OA Biomarkers

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Description
2.2. Small RNA-Sequencing
2.3. mRNA-Sequencing Dataset
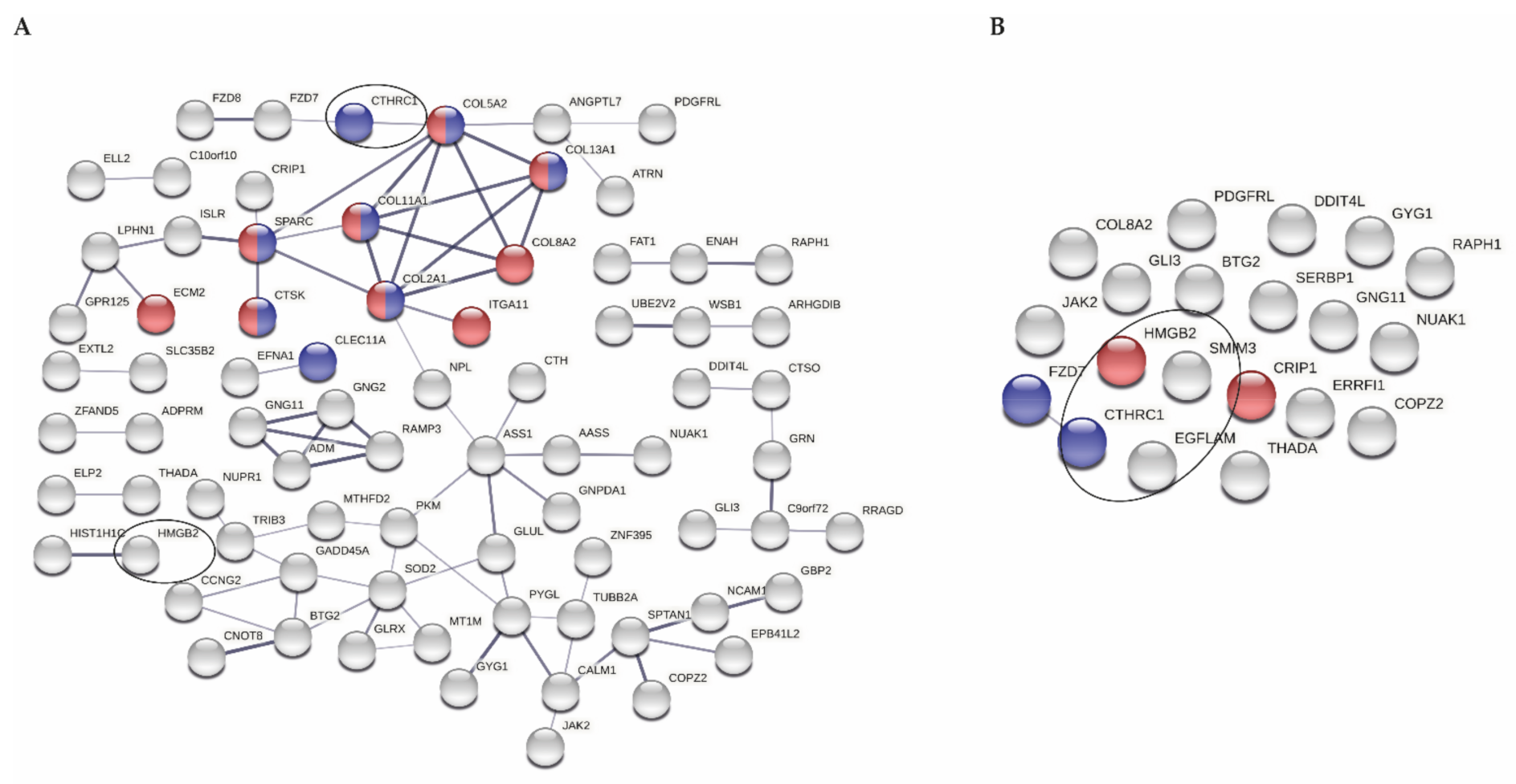
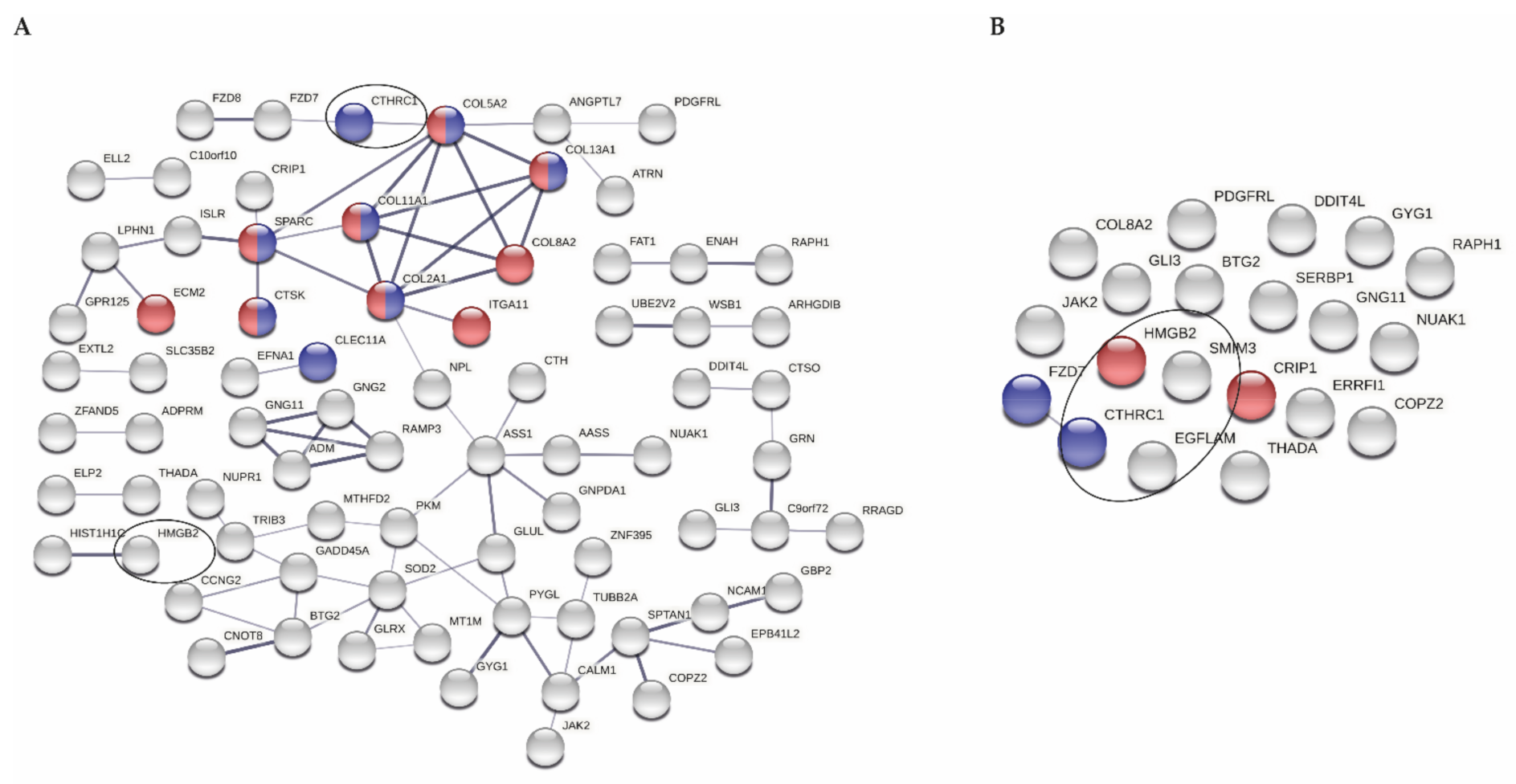
2.4. Analysis of Protein Interaction Networks
2.5. MicroRNA-mRNA Target Identification
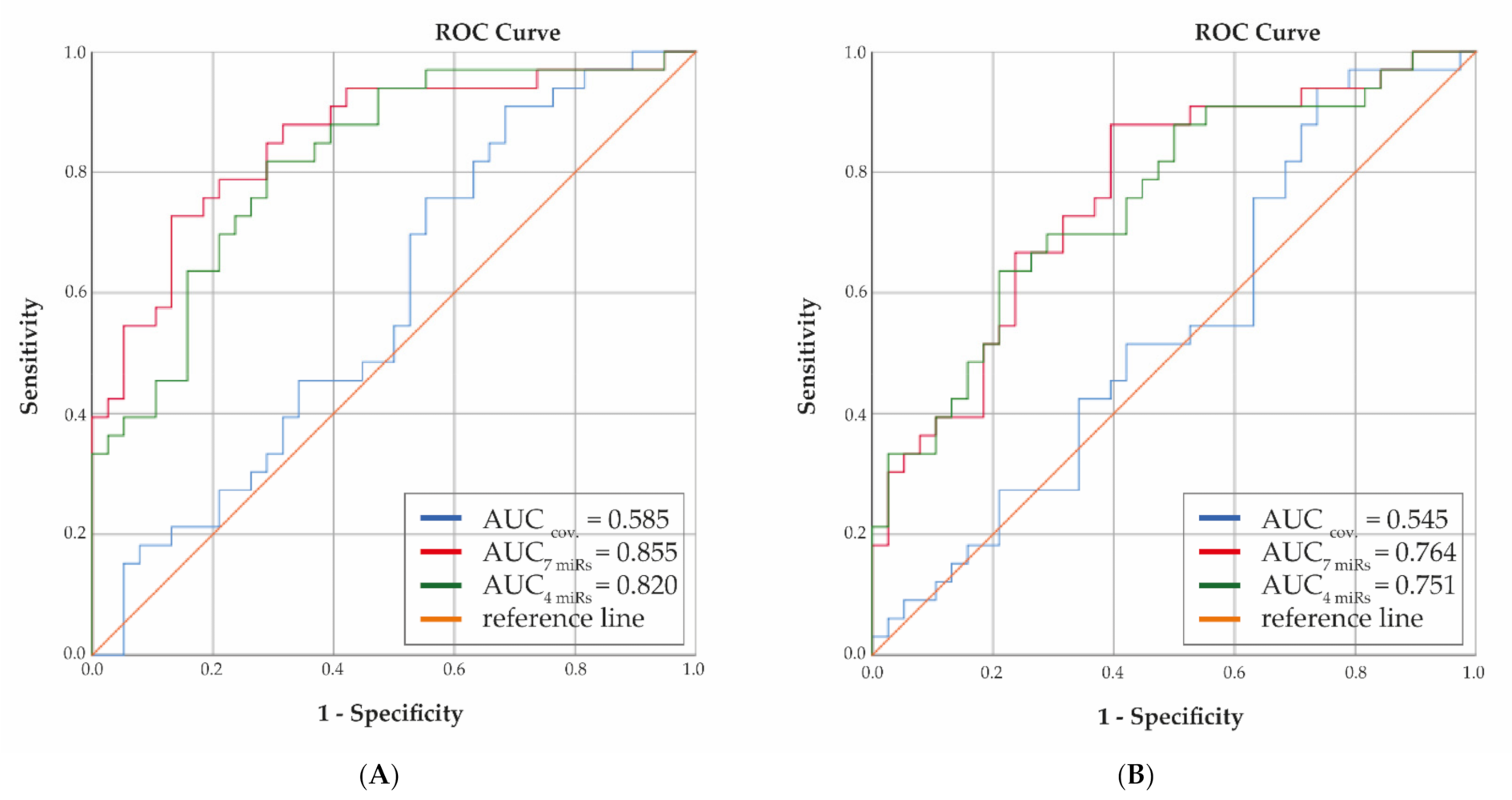
2.6. Calculation Area under Receiver Operating Curves (AUC)
2.7. Statistical Methods and Analyses
3. Results
3.1. Study Characteristics
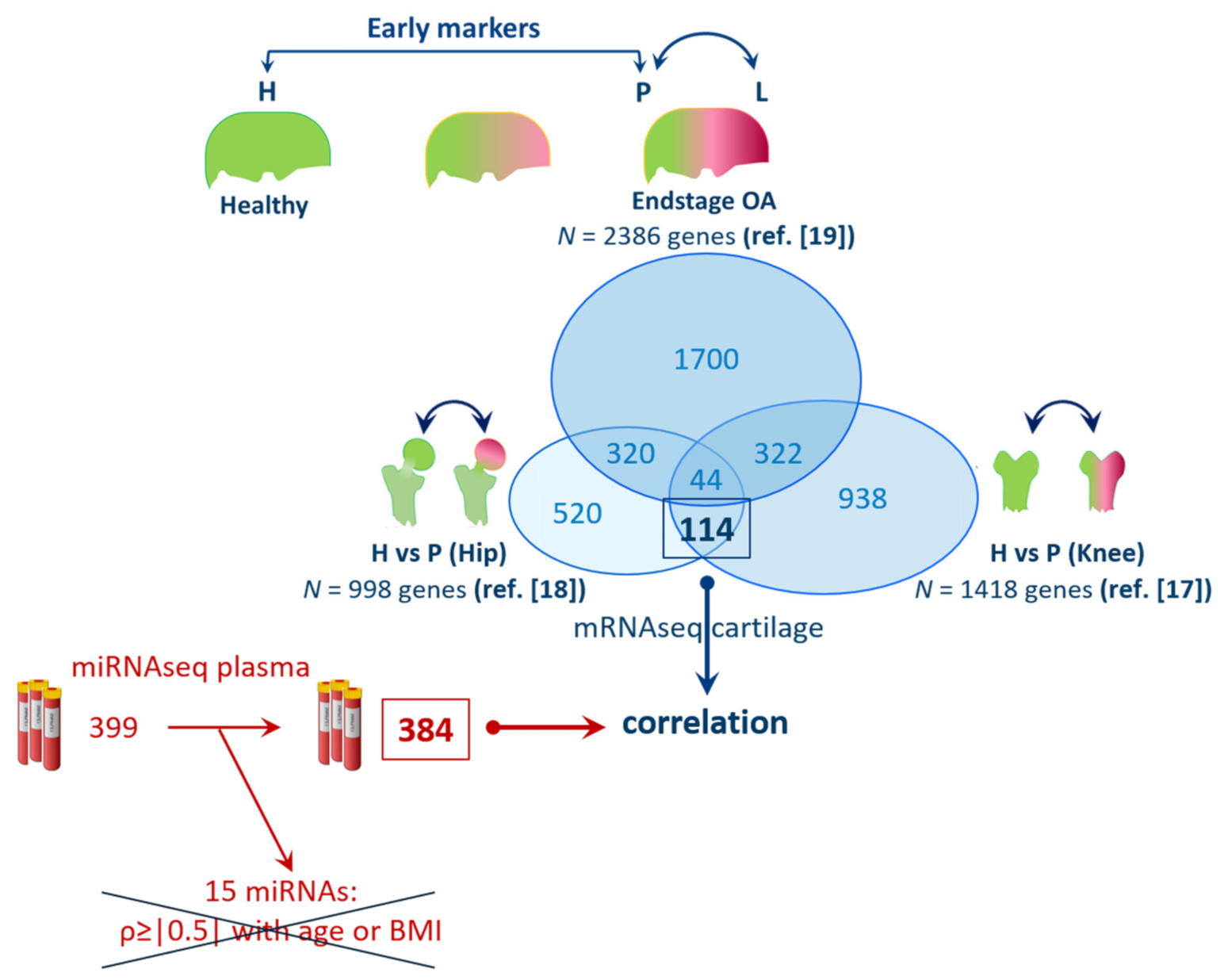
3.2. Selection of Genes Marking Early OA in Articular Cartilage
3.3. miRNA Expression Levels in Correlation with Expression of Genes Marking Early OA
3.4. Receiver Operator Curves with Selected Plasma miRNAs as Determinants of OA Progression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peat, G.; Thomas, M.J. Osteoarthritis year in review 2020: Epidemiology & therapy. Osteoarthr. Cartil. 2021, 29, 180–189. [Google Scholar]
- Safiri, S.; Kolahi, A.A.; Smith, E.; Hill, C.; Bettampadi, D.; Mansournia, M.A.; Hoy, D.; Ashrafi-Asgarabad, A.; Sepidarkish, M.; Almasi-Hashiani, A.; et al. Global, regional and national burden of osteoarthritis 1990–2017: A systematic analysis of the Global Burden of Disease Study 2017. Ann. Rheum. Dis. 2020, 79, 819–828. [Google Scholar] [CrossRef]
- Kolasinski, S.L.; Neogi, T.; Hochberg, M.C.; Oatis, C.; Guyatt, G.; Block, J.; Callahan, L.; Copenhaver, C.; Dodge, C.; Felson, D.; et al. 2019 American College of Rheumatology/Arthritis Foundation Guideline for the Management of Osteoarthritis of the Hand, Hip, and Knee. Arthritis Rheumatol. 2020, 72, 220–233. [Google Scholar] [CrossRef]
- Bernotiene, E.; Bagdonas, E.; Kirdaite, G.; Bernotas, P.; Kalvaityte, U.; Uzieliene, I.; Thudium, C.S.; Hannula, H.; Lorite, G.S.; Dvir-Ginzberg, M.; et al. Emerging Technologies and Platforms for the Immunodetection of Multiple Biochemical Markers in Osteoarthritis Research and Therapy. Front. Med. 2020, 7, 572977. [Google Scholar] [CrossRef]
- Thudium, C.S.; Nielsen, S.H.; Sardar, S.; Mobasheri, A.; van Spil, W.E.; Lories, R.; Henriksen, K.; Bay-Jensen, A.C.; Karsdal, M.A. Bone phenotypes in rheumatology—There is more to bone than just bone. BMC Musculoskelet. Disord. 2020, 21, 789. [Google Scholar] [CrossRef]
- Kraus, V.B.; Karsdal, M.A. Osteoarthritis: Current Molecular Biomarkers and the Way Forward. Calcif. Tissue Int. 2020, 109, 329–338. [Google Scholar] [CrossRef]
- Valdes, A.M.; Meulenbelt, I.; Chassaing, E.; Arden, N.K.; Bierma-Zeinstra, S.; Hart, D.; Hofman, A.; Karsdal, M.; Kloppenburg, M.; Kroon, H.M.; et al. Large scale meta-analysis of urinary C-terminal telopeptide, serum cartilage oligomeric protein and matrix metalloprotease degraded type II collagen and their role in prevalence, incidence and progression of osteoarthritis. Osteoarthr. Cartil. 2014, 22, 683–689. [Google Scholar] [CrossRef] [Green Version]
- Steyerberg, E.W. Clinical Prediction Models: A Practical Approach to Development, Validation, and Updating; Springer: New York, NY, USA, 2009; Volume 1, p. 500. [Google Scholar]
- Schummers, L.; Himes, K.P.; Bodnar, L.M.; Hutcheon, J.A. Predictor characteristics necessary for building a clinically useful risk prediction model: A simulation study. BMC Med. Res. Methodol. 2016, 16, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budd, E.; Nalesso, G.; Mobasheri, A. Extracellular genomic biomarkers of osteoarthritis. Expert Rev. Mol. Diagn. 2018, 18, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Swingler, T.E.; Niu, L.; Smith, P.; Paddy, P.; Le, L.; Barter, M.J.; Young, D.A.; Clark, I.M. The function of microRNAs in cartilage and osteoarthritis. Clin. Exp. Rheumatol. 2019, 37 (Suppl. S120), 40–47. [Google Scholar] [PubMed]
- Grillari, J.; Makitie, R.E.; Kocijan, R.; Haschka, J.; Vazquez, D.C.; Semmelrock, E.; Hackl, M. Circulating miRNAs in bone health and disease. Bone 2020, 145, 115787. [Google Scholar] [CrossRef]
- Szelenberger, R.; Kacprzak, M.; Saluk-Bijak, J.; Zielinska, M.; Bijak, M. Plasma MicroRNA as a novel diagnostic. Clin. Chim. Acta 2019, 499, 98–107. [Google Scholar] [CrossRef]
- Beyer, C.; Zampetaki, A.; Lin, N.Y.; Kleyer, A.; Perricone, C.; Iagnocco, A.; Distler, A.; Langley, S.R.; Gelse, K.; Sesselmann, S.; et al. Signature of circulating microRNAs in osteoarthritis. Ann. Rheum. Dis. 2014, 74, e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ntoumou, E.; Tzetis, M.; Braoudaki, M.; Lambrou, G.; Poulou, M.; Malizos, K.; Stefanou, N.; Anastasopoulou, L.; Tsezou, A. Serum microRNA array analysis identifies miR-140-3p, miR-33b-3p and miR-671-3p as potential osteoarthritis biomarkers involved in metabolic processes. Clin. Epigenet. 2017, 9, 127. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.A.; Gandhi, R.; Potla, P.; Keshavarzi, S.; Espin-Garcia, O.; Shestopaloff, K.; Pastrello, C.; Bethune-Waddell, D.; Lively, S.; Perruccio, A.V.; et al. Sequencing identifies a distinct signature of circulating microRNAs in early radiographic knee osteoarthritis. Osteoarthr. Cartil. 2020, 28, 1471–1481. [Google Scholar] [CrossRef]
- Karlsson, C.; Dehne, T.; Lindahl, A.; Brittberg, M.; Pruss, A.; Sittinger, M.; Ringe, J. Genome-wide expression profiling reveals new candidate genes associated with osteoarthritis. Osteoarthr. Cartil. 2010, 18, 581–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Barter, M.J.; Swan, D.C.; Rankin, K.S.; Rowan, A.D.; Santibanez-Koref, M.; Loughlin, J.; Young, D.A. Identification of the pathogenic pathways in osteoarthritic hip cartilage: Commonality and discord between hip and knee OA. Osteoarthr. Cartil. 2012, 20, 1029–1038. [Google Scholar] [CrossRef] [Green Version]
- Coutinho de Almeida, R.; Ramos, Y.F.M.; Mahfouz, A.; den Hollander, W.; Lakenberg, N.; Houtman, E.; van Hoolwerff, M.; Suchiman, H.E.D.; Rodriguez Ruiz, A.; Slagboom, P.E.; et al. RNA sequencing data integration reveals an miRNA interactome of osteoarthritis cartilage. Ann. Rheum. Dis. 2019, 78, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Ramos, Y.F.; den Hollander, W.; Bovee, J.V.; Bomer, N.; van der Breggen, R.; Lakenberg, N.; Keurentjes, J.C.; Goeman, J.J.; Slagboom, P.E.; Nelissen, R.G.; et al. Genes Involved in the Osteoarthritis Process Identified through Genome Wide Expression Analysis in Articular Cartilage; the RAAK Study. PLoS ONE 2014, 9, e103056. [Google Scholar]
- Riyazi, N.; Rosendaal, F.R.; Slagboom, E.; Kroon, H.M.; Breedveld, F.C.; Kloppenburg, M. Risk factors in familial osteoarthritis: The GARP sibling study. Osteoarthr. Cartil. 2008, 16, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Altman, R.D.; Hochberg, M.; Murphy, W.A.; Wolfe, F.; Lequesne, M. Atlas of Individual Radiographic Features in Osteoarthritis. Osteoarthr. Cartil. 1995, 3, 3–70. [Google Scholar] [CrossRef] [Green Version]
- Bijsterbosch, J.; Meulenbelt, I.; Watt, I.; Rosendaal, F.R.; Huizinga, T.W.; Kloppenburg, M. Clustering of hand osteoarthritis progression and its relationship to progression of osteoarthritis at the knee. Ann. Rheum. Dis. 2014, 73, 567–572. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Wu, T.D.; Watanabe, C.K. GMAP: A genomic mapping and alignment program for mRNA and EST sequences. Bioinformatics 2005, 21, 1859–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Franceschini, A.; Kuhn, M.; Simonovic, M.; Roth, A.; Minguez, P.; Doerks, T.; Stark, M.; Muller, J.; Bork, P.; et al. The STRING database in 2011: Functional interaction networks of proteins, globally integrated and scored. Nucleic Acids Res. 2011, 39, D561–D568. [Google Scholar] [CrossRef] [PubMed]
- Paraskevopoulou, M.D.; Georgakilas, G.; Kostoulas, N.; Vlachos, I.S.; Vergoulis, T.; Reczko, M.; Filippidis, C.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-microT web server v5.0: Service integration into miRNA functional analysis workflows. Nucleic Acids Res. 2013, 41, W169–W173. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, X. miRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.H.; Shrestha, S.; Yang, C.D.; Chang, N.W.; Lin, Y.L.; Liao, K.W.; Huang, W.C.; Sun, T.H.; Tu, S.J.; Lee, W.H.; et al. miRTarBase update 2018: A resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2018, 46, D296–D302. [Google Scholar] [CrossRef]
- Karagkouni, D.; Paraskevopoulou, M.D.; Chatzopoulos, S.; Vlachos, I.S.; Tastsoglou, S.; Kanellos, I.; Papadimitriou, D.; Kavakiotis, I.; Maniou, S.; Skoufos, G.; et al. DIANA-TarBase v8: A decade-long collection of experimentally supported miRNA-gene interactions. Nucleic Acids Res. 2018, 46, D239–D245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, Y.F.; Bos, S.D.; Lakenberg, N.; Bohringer, S.; den Hollander, W.J.; Kloppenburg, M.; Slagboom, P.E.; Meulenbelt, I. Genes expressed in blood link osteoarthritis with apoptotic pathways. Ann. Rheum. Dis. 2013, 73, 1844–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukaka, M.M. Statistics corner: A guide to appropriate use of correlation coefficient in medical research. Malawi Med. J. 2012, 24, 69–71. [Google Scholar]
- Zirkel, A.; Nikolic, M.; Sofiadis, K.; Mallm, J.P.; Brackley, C.A.; Gothe, H.; Drechsel, O.; Becker, C.; Altmuller, J.; Josipovic, N.; et al. HMGB2 Loss upon Senescence Entry Disrupts Genomic Organization and Induces CTCF Clustering across Cell Types. Mol. Cell 2018, 70, 730–744.e6. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, N.; Kawakami, Y.; Maruyama, I.; Lotz, M. HMGB proteins and arthritis. Hum. Cell 2018, 31, 1–9. [Google Scholar] [CrossRef]
- Castanheira, C.; Balaskas, P.; Falls, C.; Ashraf-Kharaz, Y.; Clegg, P.; Burke, K.; Fang, Y.; Dyer, P.; Welting, T.J.M.; Peffers, M.J. Equine synovial fluid small non-coding RNA signatures in early osteoarthritis. BMC Vet. Res. 2021, 17, 26. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Abak, A.; Tavakkoli Avval, S.; Shoorei, H.; Taheri, M.; Samadian, M. The impact of non-coding RNAs on macrophage polarization. Biomed. Pharm. 2021, 142, 112112. [Google Scholar] [CrossRef]
- Qiu, Z.; Ma, X.; Xie, J.; Liu, Z.; Zhang, Y.; Xia, C. miR-1307-5p regulates proliferation and apoptosis of chondrocytes in osteoarthritis by specifically inhibiting transforming growth factor beta-induced gene. Am. J. Transl. Res. 2021, 13, 7756–7766. [Google Scholar]
- Nakamura, A.; Rampersaud, Y.R.; Nakamura, S.; Sharma, A.; Zeng, F.; Rossomacha, E.; Ali, S.A.; Krawetz, R.; Haroon, N.; Perruccio, A.V.; et al. microRNA-181a-5p antisense oligonucleotides attenuate osteoarthritis in facet and knee joints. Ann. Rheum. Dis. 2019, 78, 111–121. [Google Scholar] [CrossRef]
- Stadnik, P.S.; Gilbert, S.J.; Tarn, J.; Charlton, S.; Skelton, A.J.; Barter, M.J.; Duance, V.C.; Young, D.A.; Blain, E.J. Regulation of microRNA-221, -222, -21 and -27 in articular cartilage subjected to abnormal compressive forces. J. Physiol. 2021, 599, 143–155. [Google Scholar] [CrossRef]
- Bottani, M.; Banfi, G.; Lombardi, G. The Clinical Potential of Circulating miRNAs as Biomarkers: Present and Future Applications for Diagnosis and Prognosis of Age-Associated Bone Diseases. Biomolecules 2020, 10, 589. [Google Scholar] [CrossRef]
- Brzeszczynska, J.; Brzeszczynski, F.; Hamilton, D.F.; McGregor, R.; Simpson, A. Role of microRNA in muscle regeneration and diseases related to muscle dysfunction in atrophy, cachexia, osteoporosis, and osteoarthritis. Bone Jt. Res. 2020, 9, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Coutinho de Almeida, R.; Mahfouz, A.; Mei, H.; Houtman, E.; den Hollander, W.; Soul, J.; Suchiman, E.; Lakenberg, N.; Meessen, J.; Huetink, K.; et al. Identification and characterization of two consistent osteoarthritis subtypes by transcriptome and clinical data integration. Rheumatology 2021, 60, 1166–1175. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | ||
| Sex | 18/22 Female | |
| Age | 55–81 (avg. 71.1) | |
| BMI | 21–33 (avg. 27.9) | |
| Joint | 16/22 Knee | |
| B | ||
| Non-Prog. | Prog. | |
| Sex | 33/38 Female | 29/33 Female |
| Age | 47–75 (mean: 61.4) | 50–69 (mean: 59.4) |
| BMI | 20–34 (mean: 26.1) | 20–40 (mean: 26.2) |
| Joint | 18/38 Knee | 17/33 Knee |
| A | ||||||
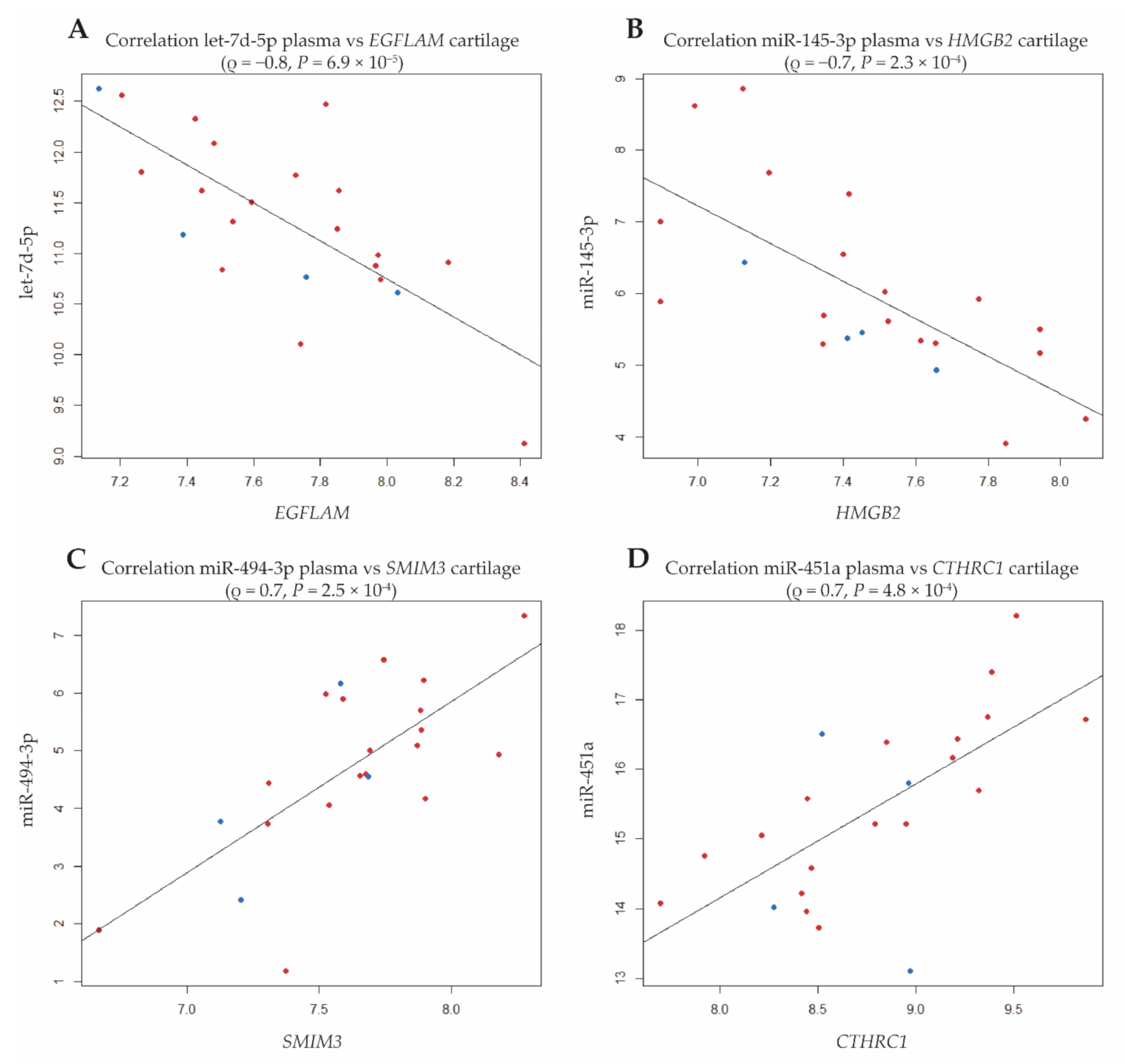
| miRNA | Gene | Corr. | P | Additional miRNAs (ρ ≥ |0.6|) | CartilageDEG (P vs. H) | |
| let-7d-5p | EGFLAM | −0.75 | 6.8 × 10−5 | let-7f-5p; let-7a-5p; miR-4443; miR-221-5p; miR-3615; miR-200b-3p; let-7e-5p; miR-1180-3p; let-7g-5p | dn | |
| miR-3928-3p | PDGFRL | −0.74 | 9.0 × 10−5 | miR-1260a; miR-106b-5p; 6852-5p; miR-23b-5p | up | |
| let-7a-5p | THADA | −0.73 | 1.0 × 10−4 | miR-339-3p; miR-22-3p | dn | |
| miR-145-3p; miR-23b-3p | HMGB2 | −0.71 | 2.3 × 10−4 | miR-181a-3p; miR-425-3p; miR-7849-3p; miR-326; miR-339-5p; miR-133a-3p; miR-3613-5p; miR-335-5p; miR-421 | dn | |
| miR-19b-3p | GLI3 | −0.71 | 2.4 × 10−4 | miR-3909; miR-23b-5p; miR-181a-5p; miR-4755-5p | up | |
| miR-494-3p | SMIM3 | 0.71 | 2.5 × 10−4 | miR-889-3p; miR-411-3p; miR-224-5p; miR-379-5p; miR-4326; miR-222-3p; miR-431-5p; miR-12136; miR-382-5p; miR-329-3p; miR-495-3p; miR-505-3p; miR-221-5p; miR-7849-3p; miR-30b-3p; miR-6772-3p; miR-493-5p; miR-381-3p | dn | |
| miR-106b-5p | COPZ2 | 0.70 | 2.7 × 10−4 | miR-939-3p; miR-210-3p | up | |
| B | ||||||
| Gene | Gene | Corr. | P | |||
| HMGB2 | SMIM3 | −0.39 | 7.0 × 10−2 | |||
| CTHRC1 | SMIM3 | −0.31 | 1.7 × 10−1 | |||
| CTHRC1 | EGFLAM | 0.22 | 3.2 × 10−1 | |||
| CTHRC1 | HMGB2 | 0.22 | 3.3 × 10−1 | |||
| EGFLAM | HMGB2 | 0.19 | 4.0 × 10−1 | |||
| EGFLAM | SMIM3 | −0.10 | 6.5 × 10−1 | |||
| CTHRC1 | EGFLAM | HMGB2 | SMIM3 | |||
| CTHRC1 | 1.00 | |||||
| EGFLAM | 0.22 | 1.00 | ||||
| HMGB2 | 0.22 | 0.19 | 1.00 | |||
| SMIM3 | −0.31 | −0.10 | −0.39 | 1.00 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos, Y.F.M.; Coutinho de Almeida, R.; Lakenberg, N.; Suchiman, E.; Mei, H.; Kloppenburg, M.; Nelissen, R.G.H.H.; Meulenbelt, I. Circulating MicroRNAs Highly Correlate to Expression of Cartilage Genes Potentially Reflecting OA Susceptibility—Towards Identification of Applicable Early OA Biomarkers. Biomolecules 2021, 11, 1356. https://doi.org/10.3390/biom11091356
Ramos YFM, Coutinho de Almeida R, Lakenberg N, Suchiman E, Mei H, Kloppenburg M, Nelissen RGHH, Meulenbelt I. Circulating MicroRNAs Highly Correlate to Expression of Cartilage Genes Potentially Reflecting OA Susceptibility—Towards Identification of Applicable Early OA Biomarkers. Biomolecules. 2021; 11(9):1356. https://doi.org/10.3390/biom11091356
Chicago/Turabian StyleRamos, Yolande F. M., Rodrigo Coutinho de Almeida, Nico Lakenberg, Eka Suchiman, Hailiang Mei, Margreet Kloppenburg, Rob G. H. H. Nelissen, and Ingrid Meulenbelt. 2021. "Circulating MicroRNAs Highly Correlate to Expression of Cartilage Genes Potentially Reflecting OA Susceptibility—Towards Identification of Applicable Early OA Biomarkers" Biomolecules 11, no. 9: 1356. https://doi.org/10.3390/biom11091356
APA StyleRamos, Y. F. M., Coutinho de Almeida, R., Lakenberg, N., Suchiman, E., Mei, H., Kloppenburg, M., Nelissen, R. G. H. H., & Meulenbelt, I. (2021). Circulating MicroRNAs Highly Correlate to Expression of Cartilage Genes Potentially Reflecting OA Susceptibility—Towards Identification of Applicable Early OA Biomarkers. Biomolecules, 11(9), 1356. https://doi.org/10.3390/biom11091356