
Multi-Layered Regulations on the Chromatin Architectures: Establishing the Tight and Specific Responses of Fission Yeast fbp1 Gene Transcription


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
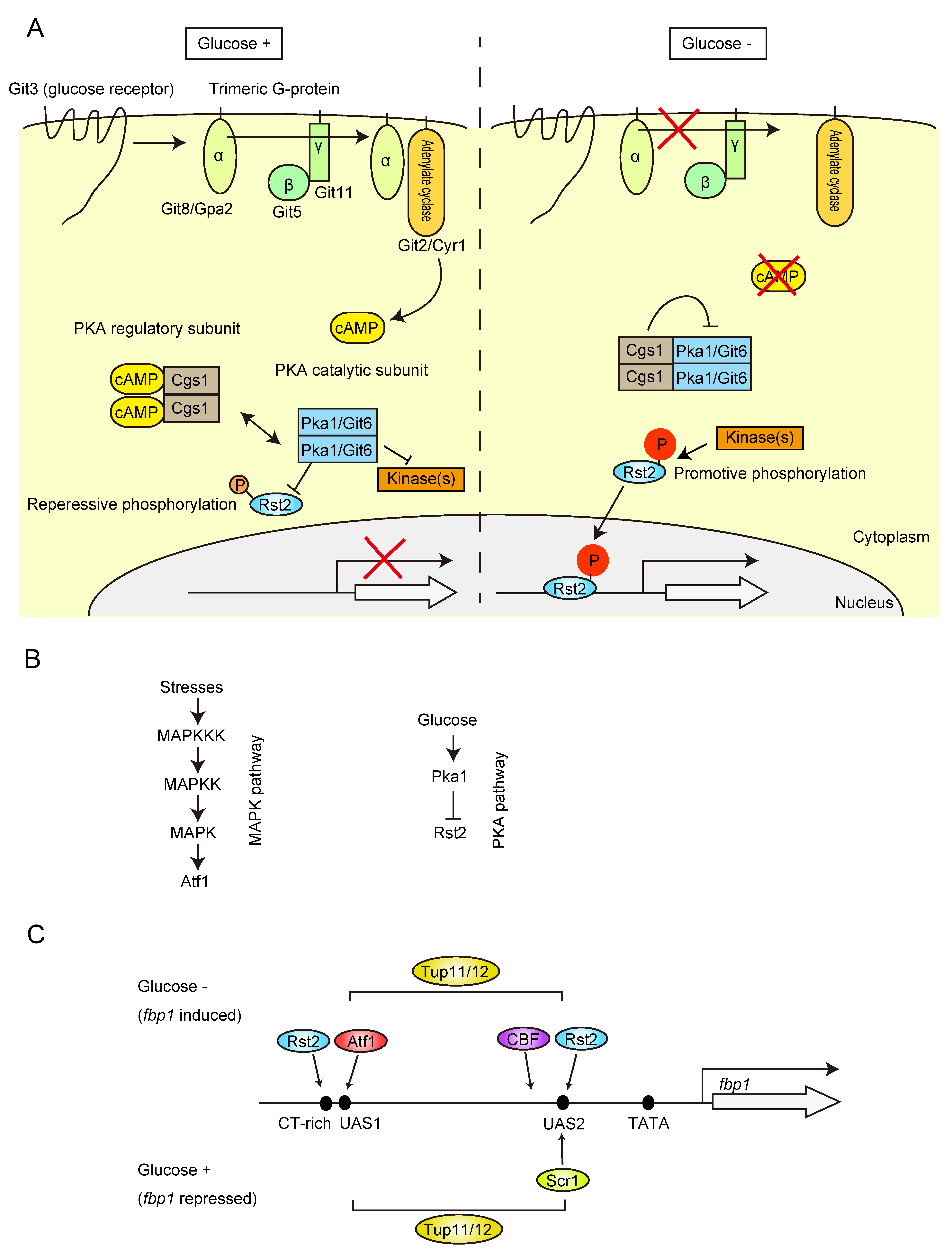
2. Signal Pathways and Transcription Factors Required for the Regulation of fbp1 Gene
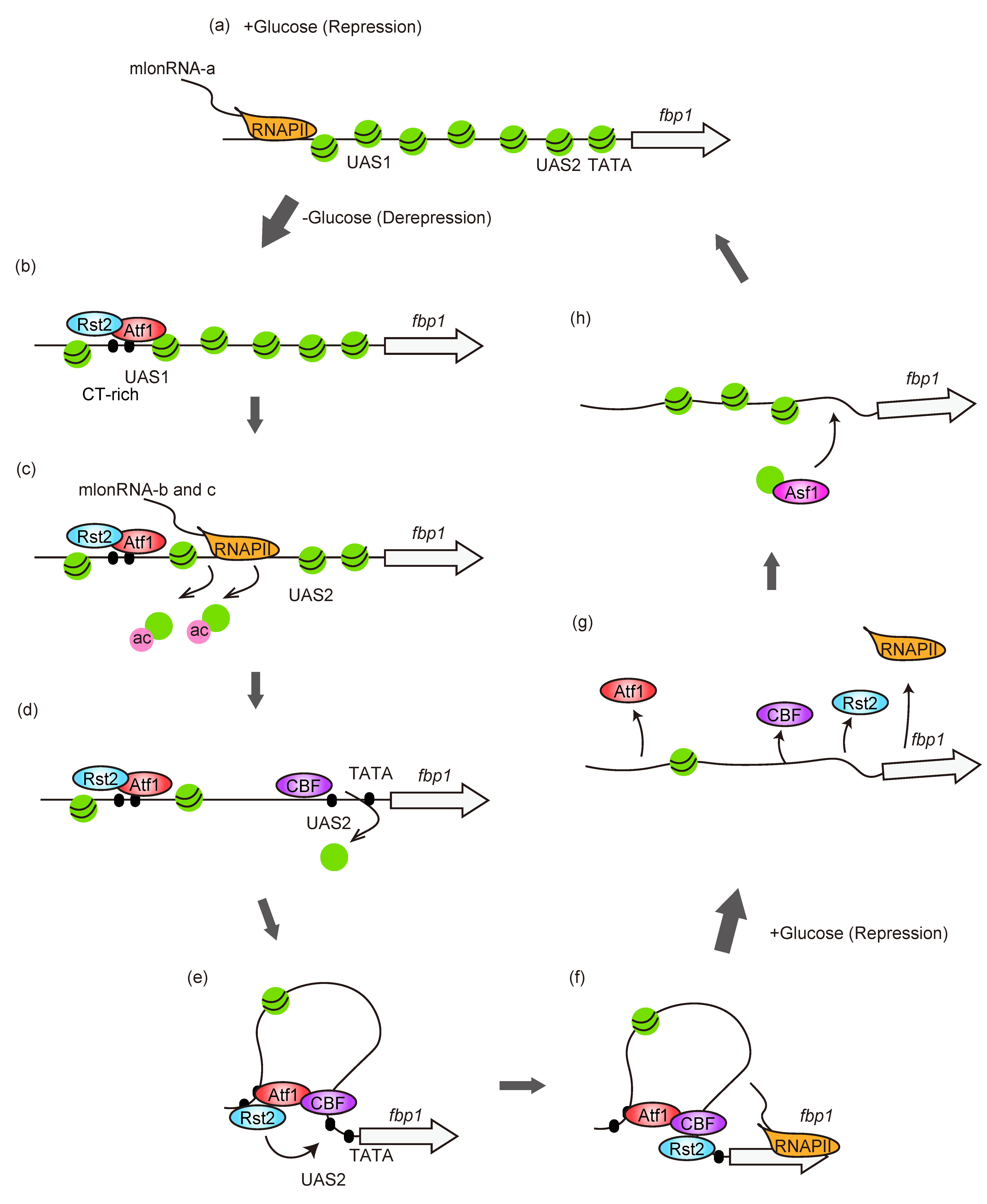
3. Chromatin Dynamics at fbp1 Promoter Region
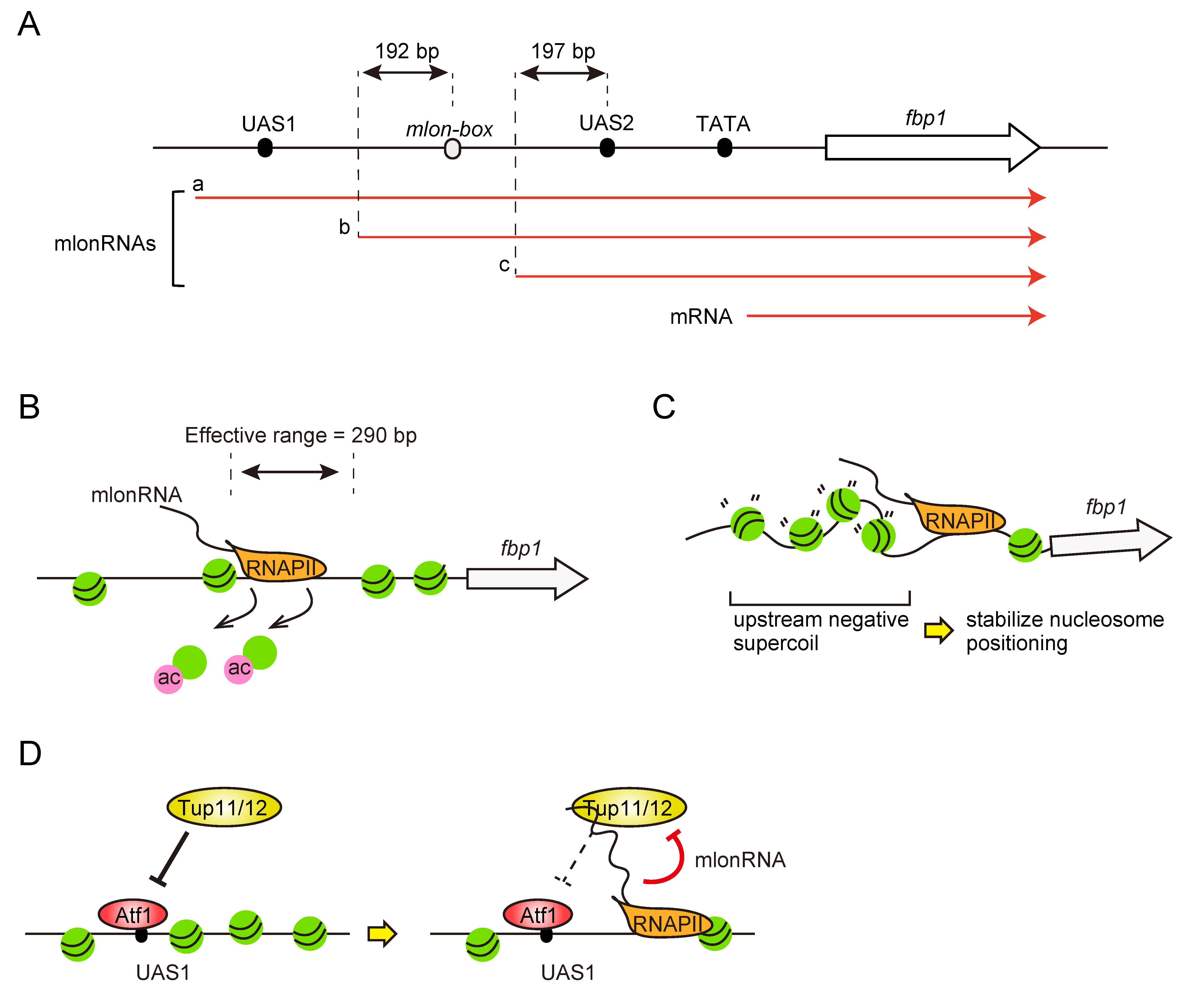
4. mlonRNAs-Transcription-Mediated Regulation of Chromatin and Transcription Factor Binding for fbp1 Transcriptional Regulation
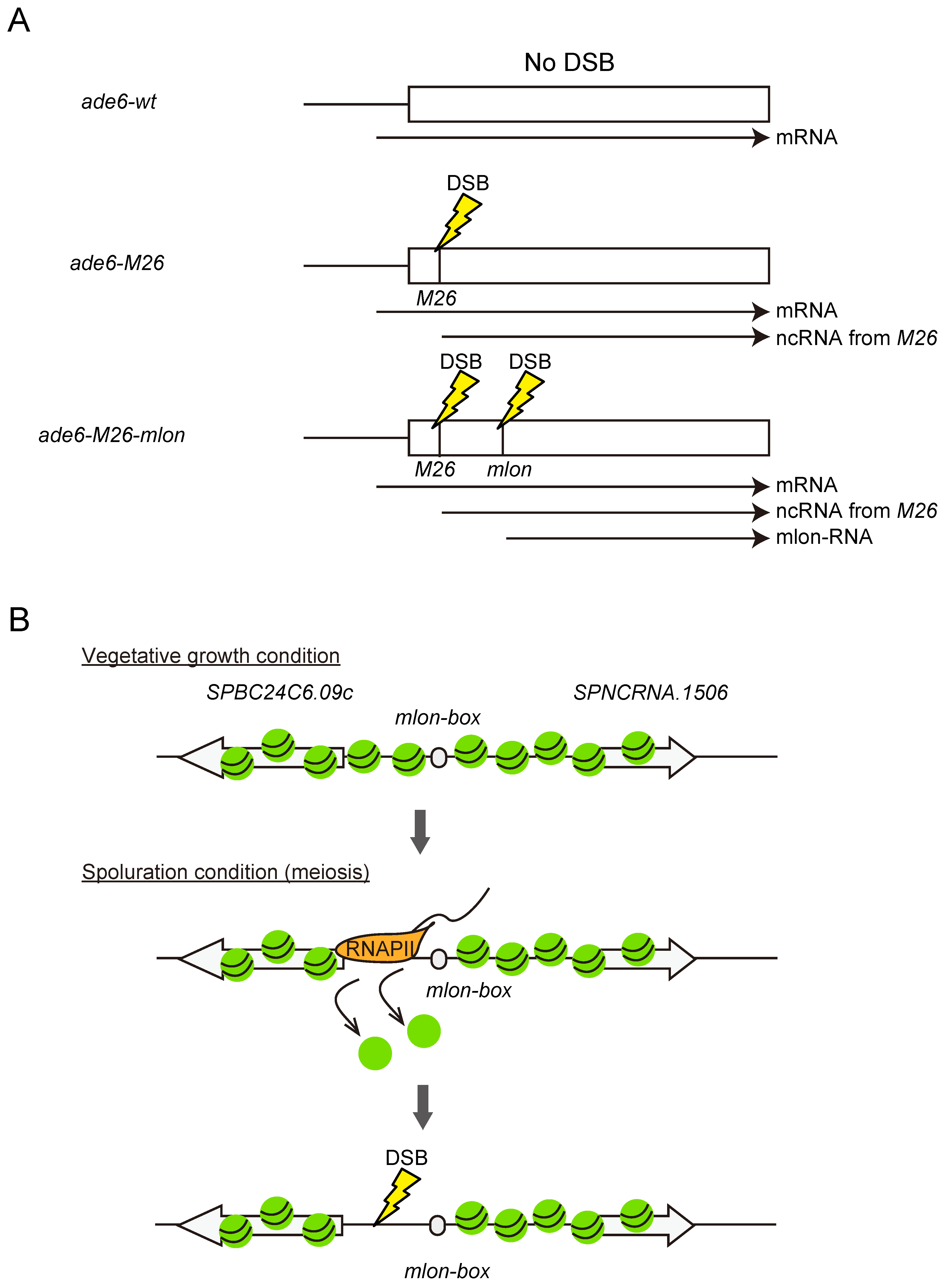
5. mlonRNA Transcription Plays Roles in the Regulation of General Chromosomal Function in Fission Yeast Genome
6. Tup Co-Repressor Mediated Multi-Layered Regulations
6.1. Local Proximity of Two TF-Binding Motifs Integrates Distinct Signal Pathway on Genome for Antagonizing Tup-Mediated Inhibition of Atf1 and Rst2 Binding
6.2. Role of Genome-Local Loop Structure in the Precise TF-Binding and Transcriptional Initiation
7. Summary and Perspective
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Wouters, J.; Atak, Z.K.; Aerts, S. Decoding transcriptional states in cancer. Curr. Opin. Genet. Dev. 2017, 43, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Deplancke, B.; Alpern, D.; Gardeux, V. The Genetics of Transcription Factor DNA Binding Variation. Cell 2016, 166, 538–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacQuarrie, K.L.; Fong, A.P.; Morse, R.H.; Tapscott, S.J. Genome-wide transcription factor binding: Beyond direct target regulation. Trends Genet. 2011, 27, 141–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klemm, S.L.; Shipony, Z.; Greenleaf, W.J. Chromatin accessibility and the regulatory epigenome. Nat. Rev. Genet. 2019, 20, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Wolffe, A.P. Nucleosome positioning and modification: Chromatin structures that potentiate transcription. Trends Biochem. Sci. 1994, 19, 240–244. [Google Scholar] [CrossRef]
- Wolffe, A.P. Histones, nucleosomes and the roles of chromatin structure in transcriptional control. Biochem. Soc. Trans. 1997, 25, 354–358. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Xie, W. The role of 3D genome organization in development and cell differentiation. Nat. Rev. Mol. Cell Biol. 2019, 20, 535–550. [Google Scholar] [CrossRef]
- Turner, B.M. Cellular Memory and the Histone Code. Cell 2002, 111, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Grant, P.A.; Sterner, D.E.; Duggan, L.J.; Workman, J.L.; Berger, S.L. The SAGA unfolds: Convergence of transcription regulators in chromatin-modifying complexes. Trends Cell Biol. 1998, 8, 193–197. [Google Scholar] [CrossRef]
- Kuo, M.H.; Allis, C.D. Roles of histone acetyltransferases and deacetylases in gene regulation. Bioessays 1998, 20, 615–626. [Google Scholar] [CrossRef]
- Travers, A. An Engine for Nucleosome Remodeling. Cell 1999, 96, 311–314. [Google Scholar] [CrossRef] [Green Version]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forrest, A.R.; Kawaji, H.; Rehli, M.; Baillie, J.K.; de Hoon, M.J.; Haberle, V.; Lassmann, T.; Kulakovskiy, I.V.; Lizio, M.; Itoh, M.; et al. A promoter-level mammalian expression atlas. Nature 2014, 507, 462–470. [Google Scholar] [PubMed] [Green Version]
- Grewal, S.I. RNAi-dependent formation of heterochromatin and its diverse functions. Curr. Opin. Genet. Dev. 2010, 20, 134–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Wolfswinkel, J.C.; Ketting, R.F. The role of small non-coding RNAs in genome stability and chromatin organization. J. Cell Sci. 2010, 123, 1825–1839. [Google Scholar] [CrossRef] [Green Version]
- Bonasio, R.; Shiekhattar, R. Regulation of Transcription by Long Noncoding RNAs. Annu. Rev. Genet. 2014, 48, 433–455. [Google Scholar] [CrossRef] [Green Version]
- Marchese, F.P.; Raimondi, I.; Huarte, M. The multidimensional mechanisms of long noncoding RNA function. Genome Biol. 2017, 18, 206. [Google Scholar] [CrossRef] [Green Version]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Vassarotti, A.; Friesen, J.D. Isolation of the fructose-1,6-bisphosphatase gene of the yeast Schizosaccharomyces pombe. Evidence for transcriptional regulation. J. Biol. Chem. 1985, 260, 6348–6353. [Google Scholar] [CrossRef]
- Benkovic, S.J.; Demaine, M.M. Mechanism of Action of Fructose 1,6-Bisphosphatase. Adv. Enzymol. Relat. Areas Mol. Biol. 1982, 53, 45–82. [Google Scholar] [CrossRef]
- Holzer, H. Proteolytic catabolite inactivation in Saccharomyces cerevisiae. Revis. Sobre Biol. Cel. RBC 1989, 21, 305–319. [Google Scholar]
- Sedivy, J.M.; Fraenkel, D.G. Fructose bisphosphatase of Saccharomyces cerevisiae: Cloning, disruption and regulation of the FBP1 structural gene. J. Mol. Biol. 1985, 186, 307–319. [Google Scholar] [CrossRef]
- Byrne, S.; Hoffman, C. Six git genes encode a glucose-induced adenylate cyclase activation pathway in the fission yeast Schizosaccharomyces pombe. J. Cell Sci. 1993, 105, 1095–1100. [Google Scholar] [CrossRef]
- Hoffman, C.; Winston, F. Isolation and characterization of mutants constitutive for expression of the fbp1 gene of Schizosaccharomyces pombe. Genetics 1990, 124, 807–816. [Google Scholar] [CrossRef]
- Hoffman, C.S.; Winston, F. Glucose repression of transcription of the Schizosaccharomyces pombe fbp1 gene occurs by a cAMP signaling pathway. Genes Dev. 1991, 5, 561–571. [Google Scholar] [CrossRef] [Green Version]
- Landry, S.; Hoffman, C.S. The git5 Gbeta and git11 Ggamma form an atypical Gbetagamma dimer acting in the fission yeast glucose/cAMP pathway. Genetics 2001, 157, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Landry, S.; Pettit, M.T.; Apolinario, E.; Hoffman, C.S. The fission yeast git5 gene encodes a Gbeta subunit required for glucose-triggered adenylate cyclase activation. Genetics 2000, 154, 1463–1471. [Google Scholar] [CrossRef] [PubMed]
- Nocero, M.; Isshiki, T.; Yamamoto, M.; Hoffman, C. Glucose repression of fbp1 transcription of Schizosaccharomyces pombe is partially regulated by adenylate cyclase activation by a G protein alpha subunit encoded by gpa2 (git8). Genetics 1994, 138, 39–45. [Google Scholar] [CrossRef]
- Schadick, K.; Fourcade, H.M.; Boumenot, P.; Seitz, J.J.; Morrell, J.L.; Chang, L.; Gould, K.L.; Partridge, J.F.; Allshire, R.C.; Kitagawa, K.; et al. Schizosaccharomyces pombe Git7p, a member of the Saccharomyces cerevisiae Sgtlp family, is required for glucose and cyclic AMP signaling, cell wall integrity, and septation. Eukaryot. Cell 2002, 1, 558–567. [Google Scholar] [CrossRef] [Green Version]
- Welton, R.M.; Hoffman, C.S. Glucose monitoring in fission yeast via the Gpa2 galpha, the git5 Gbeta and the git3 putative glucose receptor. Genetics 2000, 156, 513–521. [Google Scholar] [CrossRef]
- Isshiki, T.; Mochizuki, N.; Maeda, T.; Yamamoto, M. Characterization of a fission yeast gene, gpa2, that encodes a G alpha subunit involved in the monitoring of nutrition. Genes Dev. 1992, 6, 2455–2462. [Google Scholar] [CrossRef] [PubMed]
- Kunitomo, H.; Higuchi, T.; Iino, Y.; Yamamoto, M. A Zinc-Finger Protein, Rst2p, regulates transcription of the fission yeast ste11(+) gene, Which Encodes a Pivotal Transcription Factor for Sexual Development. Mol. Biol. Cell 2000, 11, 3205–3217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, T.; Mochizuki, N.; Yamamoto, M. Adenylyl cyclase is dispensable for vegetative cell growth in the fission yeast Schizosaccharomyces pombe. Proc. Natl. Acad. Sci. USA 1990, 87, 7814–7818. [Google Scholar] [CrossRef] [Green Version]
- Maeda, T.; Watanabe, Y.; Kunitomo, H.; Yamamoto, M. Cloning of the pka1 gene encoding the catalytic subunit of the cAMP-dependent protein kinase in Schizosaccharomyces pombe. J. Biol. Chem. 1994, 269, 9632–9637. [Google Scholar] [CrossRef]
- Higuchi, T.; Watanabe, Y.; Yamamoto, M. Protein Kinase A Regulates Sexual Development and Gluconeogenesis through Phosphorylation of the Zn Finger Transcriptional Activator Rst2p in Fission Yeast. Mol. Cell. Biol. 2002, 22, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hirota, K.; Hoffman, C.S.; Ohta, K. Reciprocal Nuclear Shuttling of Two Antagonizing Zn Finger Proteins Modulates Tup Family Corepressor Function To Repress Chromatin Remodeling. Eukaryot. Cell 2006, 5, 1980–1989. [Google Scholar] [CrossRef] [Green Version]
- Takeda, T.; Toda, T.; Kominami, K.; Kohnosu, A.; Yanagida, M.; Jones, N. Schizosaccharomyces pombe atf1+ encodes a transcription factor required for sexual development and entry into stationary phase. EMBO J. 1995, 14, 6193–6208. [Google Scholar] [CrossRef]
- Kanoh, J.; Watanabe, Y.; Ohsugi, M.; Iino, Y.; Yamamoto, M. Schizosaccharomyces pombe gad7 + encodes a phosphoprotein with a bZIP domain, which is required for proper G1 arrest and gene expression under nitrogen starvation. Genes Cells 1996, 1, 391–408. [Google Scholar] [CrossRef]
- Neely, L.A.; Hoffman, C.S. Protein kinase A and mitogen-activated protein kinase pathways antagonistically regulate fission yeast fbp1 transcription by employing different modes of action at two upstream activation sites. Mol. Cell Biol. 2000, 20, 6426–6434. [Google Scholar] [CrossRef]
- Shiozaki, K.; Russell, P. Conjugation, meiosis, and the osmotic stress response are regulated by Spc1 kinase through Atf1 transcription factor in fission yeast. Genes Dev. 1996, 10, 2276–2288. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, M.G.; Samuels, M.; Takeda, T.; Toone, W.M.; Shieh, J.C.; Toda, T.; Millar, J.B.; Jones, N. The Atf1 transcription factor is a target for the Sty1 stress-activated MAP kinase pathway in fission yeast. Genes Dev. 1996, 10, 2289–2301. [Google Scholar] [CrossRef] [PubMed]
- Kon, N.; Krawchuk, M.D.; Warren, B.G.; Smith, G.R.; Wahls, W.P. Transcription factor Mts1/Mts2 (Atf1/Pcr1, Gad7/Pcr1) activates the M26 meiotic recombination hotspot in Schizosaccharomyces pombe. Proc. Natl. Acad. Sci. USA 1997, 94, 13765–13770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asada, R.; Umeda, M.; Adachi, A.; Senmatsu, S.; Abe, T.; Iwasaki, H.; Ohta, K.; Hoffman, C.; Hirota, K. Recruitment and delivery of the fission yeast Rst2 transcription factor via a local genome structure counteracts repression by Tup1-family corepressors. Nucleic Acids Res. 2017, 45, 9361–9371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janoo, R.T.K.; Neely, L.A.; Braun, B.R.; Whitehall, S.K.; Hoffman, C.S. Transcriptional Regulators of the Schizosaccharomyces pombe fbp1 Gene Include Two Redundant Tup1p-like Corepressors and the CCAAT Binding Factor Activation Complex. Genetics 2001, 157, 1205–1215. [Google Scholar] [CrossRef] [PubMed]
- Dolfini, D.; Gatta, R.; Mantovani, R. NF-Y and the transcriptional activation of CCAAT promoters. Crit. Rev. Biochem. Mol. Biol. 2011, 47, 29–49. [Google Scholar] [CrossRef]
- McNabb, D.S.; Tseng, K.A.; Guarente, L. The Saccharomyces cerevisiae Hap5p homolog from fission yeast reveals two conserved domains that are essential for assembly of heterotetrameric CCAAT-binding factor. Mol. Cell. Biol. 1997, 17, 7008–7018. [Google Scholar] [CrossRef] [Green Version]
- Oldfield, A.J.; Henriques, T.; Kumar, D.; Burkholder, A.B.; Cinghu, S.; Paulet, D.; Bennett, B.D.; Yang, P.; Scruggs, B.S.; Lavender, C.A.; et al. NF-Y controls fidelity of transcription initiation at gene promoters through maintenance of the nucleosome-depleted region. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Asada, R.; Takemata, N.; Hoffman, C.; Ohta, K.; Hirota, K. Antagonistic Controls of Chromatin and mRNA Start Site Selection by Tup Family Corepressors and the CCAAT-Binding Factor. Mol. Cell. Biol. 2015, 35, 847–855. [Google Scholar] [CrossRef] [Green Version]
- Malavé, T.M.; Dent, S.Y. Transcriptional repression by Tup1-Ssn6. Biochem. Cell Biol. 2006, 84, 437–443. [Google Scholar] [CrossRef]
- Han, B.K.; Emr, S.D. Phosphoinositide [PI(3,5)P2] lipid-dependent regulation of the general transcriptional regulator Tup1. Genes Dev. 2011, 25, 984–995. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-J.; Swanson, M.J.; Qiu, H.; Govind, C.K.; Hinnebusch, A.G. Activator Gcn4p and Cyc8p/Tup1p Are Interdependent for Promoter Occupancy at ARG1 In Vivo. Mol. Cell. Biol. 2005, 25, 11171–11183. [Google Scholar] [CrossRef] [PubMed]
- Hirota, K.; Hasemi, T.; Yamada, T.; Mizuno, K.I.; Hoffman, C.S.; Shibata, T.; Ohta, K. Fission yeast global repressors regulate the specificity of chromatin alteration in response to distinct environmental stresses. Nucleic. Acids Res. 2004, 32, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Hirota, K.; Hoffman, C.S.; Shibata, T.; Ohta, K. Fission Yeast Tup1-Like Repressors Repress Chromatin Remodeling at the fbp1+ Promoter and the ade6-M26 Recombination Hotspot. Genetics 2003, 165, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Hirota, K.; Miyoshi, T.; Kugou, K.; Hoffman, C.S.; Shibata, T.; Ohta, K. Stepwise chromatin remodelling by a cascade of transcription initiation of non-coding RNAs. Nature 2008, 456, 130–134. [Google Scholar] [CrossRef]
- Hirota, K.; Ohta, K. Cascade transcription of mRNA-type long non-coding RNAs (mlonRNAs) and local chromatin remodeling. Epigenetics 2009, 4, 5–7. [Google Scholar] [CrossRef] [Green Version]
- Hirota, K.; Ohta, K. Transcription of mRNA-type long non-coding RNAs (mlonRNAs) disrupts chromatin array. Commun. Integr. Biol. 2009, 2, 25–26. [Google Scholar] [CrossRef]
- Galipon, J.; Miki, A.; Oda, A.; Inada, T.; Ohta, K. Stress-induced lncRNAs evade nuclear degradation and enter the translational machinery. Genes Cells 2013, 18, 353–368. [Google Scholar] [CrossRef]
- Oda, A.; Takemata, N.; Hirata, Y.; Miyoshi, T.; Suzuki, Y.; Sugano, S.; Ohta, K. Dynamic transition of transcription and chromatin landscape during fission yeast adaptation to glucose starvation. Genes Cells 2015, 20, 392–407. [Google Scholar] [CrossRef]
- Umeda, M.; Tsunekawa, C.; Senmatsu, S.; Asada, R.; Abe, T.; Ohta, K.; Hoffman, C.S.; Hirota, K. Histone Chaperone Asf1 Is Required for the Establishment of Repressive Chromatin in Schizosaccharomyces pombe fbp1 Gene Repression. Mol. Cell. Biol. 2018, 38, e00194-18. [Google Scholar] [CrossRef] [Green Version]
- Hammond, C.; Strømme, C.B.; Huang, H.; Patel, H.H.D.J.; Groth, C.M.H.C.B.S.A. Histone chaperone networks shaping chromatin function. Nat. Rev. Mol. Cell Biol. 2017, 18, 141–158. [Google Scholar] [CrossRef] [Green Version]
- Adachi, A.; Senmatsu, S.; Asada, R.; Abe, T.; Hoffman, C.S.; Ohta, K.; Hirota, K. Interplay between chromatin modulators and histone acetylation regulates the formation of accessible chromatin in the upstream regulatory region of fission yeast fbp1. Genet. Syst. 2017, 92, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Takemata, N.; Oda, A.; Yamada, T.; Galipon, J.; Miyoshi, T.; Suzuki, Y.; Sugano, S.; Hoffman, C.S.; Hirota, K.; Ohta, K. Local potentiation of stress-responsive genes by upstream noncoding transcription. Nucleic Acids Res. 2016, 44, 5174–5189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senmatsu, S.; Asada, R.; Abe, T.; Hoffman, C.; Ohta, K.; Hirota, K. lncRNA transcriptional initiation induces chromatin remodeling within a limited range in the fission yeast fbp1 promoter. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Whitehouse, I.; Rando, O.J.; Delrow, J.; Tsukiyama, T. Chromatin remodelling at promoters suppresses antisense transcription. Nature 2007, 450, 1031–1035. [Google Scholar] [CrossRef]
- Fry, C.J.; Peterson, C.L. Chromatin remodeling enzymes: Who’s on first? Curr. Biol. 2001, 11, R185–R197. [Google Scholar] [CrossRef] [Green Version]
- Asada, R.; Senmatsu, S.; Montpetit, B.; Hirota, K. Topoisomerase activity is linked to altered nucleosome positioning and transcriptional regulation in the fission yeast fbp1 gene. PLoS ONE 2020, 15, e0242348. [Google Scholar] [CrossRef] [PubMed]
- Koster, D.A.; Crut, A.; Shuman, S.; Bjornsti, M.-A.; Dekker, N.H. Cellular Strategies for Regulating DNA Supercoiling: A Single-Molecule Perspective. Cell 2010, 142, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.C. Cellular roles of DNA topoisomerases: A molecular perspective. Nat. Rev. Mol. Cell Biol. 2002, 3, 430–440. [Google Scholar] [CrossRef]
- Kouzine, F.; Gupta, A.; Baranello, L.; Wojtowicz, D.; Ben-Aissa, K.; Liu, J.; Przytycka, T.M.; Levens, D. Transcription-dependent dynamic supercoiling is a short-range genomic force. Nat. Struct. Mol. Biol. 2013, 20, 396–403. [Google Scholar] [CrossRef] [Green Version]
- Teves, S.S.; Henikoff, S. Transcription-generated torsional stress destabilizes nucleosomes. Nat. Struct. Mol. Biol. 2013, 21, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Zlatanova, J.; Tomschik, M. Nucleosome Assembly Depends on the Torsion in the DNA Molecule: A Magnetic Tweezers Study. Biophys. J. 2009, 97, 3150–3157. [Google Scholar] [CrossRef] [PubMed]
- Gavin, I.; Horn, P.J.; Peterson, C.L. SWI/SNF Chromatin Remodeling Requires Changes in DNA Topology. Mol. Cell 2001, 7, 97–104. [Google Scholar] [CrossRef]
- Senmatsu, S.; Asada, R.; Oda, A.; Hoffman, C.S.; Ohta, K.; Hirota, K. lncRNA transcription induces meiotic recombination through chromatin remodelling in fission yeast. Commun. Biol. 2021, 4, 1–10. [Google Scholar] [CrossRef]
- Lam, I.; Keeney, S. Mechanism and Regulation of Meiotic Recombination Initiation. Cold Spring Harb. Perspect. Biol. 2014, 7, a016634. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Sasaki, M.; Kniewel, R.; Murakami, H.; Blitzblau, H.G.; Tischfield, S.E.; Zhu, X.; Neale, M.J.; Jasin, M.; Socci, N.D.; et al. A Hierarchical Combination of Factors Shapes the Genome-wide Topography of Yeast Meiotic Recombination Initiation. Cell 2011, 144, 719–731. [Google Scholar] [CrossRef] [Green Version]
- Ponticelli, A.S.; Sena, E.P.; Smith, G. Genetic and physical analysis of the M26 recombination hotspot of Schizosaccharomyces pombe. Genetics 1988, 119, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Okamura, M.; Oda, A.; Murakami, H.; Ohta, K.; Yamada, T. Correlation of Meiotic DSB Formation and Transcription Initiation Around Fission Yeast Recombination Hotspots. Genetics 2017, 206, 801–809. [Google Scholar] [CrossRef] [Green Version]
- Koda, W.; Senmatsu, S.; Abe, T.; Hoffman, C.S.; Hirota, K. Reciprocal stabilization of transcription factor binding integrates two signaling pathways to regulate fission yeast fbp1 transcription. Nucleic Acids Res. 2021, 49, 9809–9820. [Google Scholar] [CrossRef]
- McCord, R.P.; Kaplan, N.; Giorgetti, L. Chromosome Conformation Capture and Beyond: Toward an Integrative View of Chromosome Structure and Function. Mol. Cell 2020, 77, 688–708. [Google Scholar] [CrossRef]
- Singh, B.; Hampsey, M. A Transcription-Independent Role for TFIIB in Gene Looping. Mol. Cell 2007, 27, 806–816. [Google Scholar] [CrossRef]
- Tan-Wong, S.M.; French, J.D.; Proudfoot, N.J.; Brown, M.A. Dynamic interactions between the promoter and terminator regions of the mammalian BRCA1 gene. Proc. Natl. Acad. Sci. USA 2008, 105, 5160–5165. [Google Scholar] [CrossRef] [PubMed]
- Tan-Wong, S.M.; Wijayatilake, H.D.; Proudfoot, N.J. Gene loops function to maintain transcriptional memory through interaction with the nuclear pore complex. Genes Dev. 2009, 23, 2610–2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adelman, K.; Lis, J.T. Promoter-proximal pausing of RNA polymerase II: Emerging roles in metazoans. Nat. Rev. Genet. 2012, 13, 720–731. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asada, R.; Hirota, K. Multi-Layered Regulations on the Chromatin Architectures: Establishing the Tight and Specific Responses of Fission Yeast fbp1 Gene Transcription. Biomolecules 2022, 12, 1642. https://doi.org/10.3390/biom12111642
Asada R, Hirota K. Multi-Layered Regulations on the Chromatin Architectures: Establishing the Tight and Specific Responses of Fission Yeast fbp1 Gene Transcription. Biomolecules. 2022; 12(11):1642. https://doi.org/10.3390/biom12111642
Chicago/Turabian StyleAsada, Ryuta, and Kouji Hirota. 2022. "Multi-Layered Regulations on the Chromatin Architectures: Establishing the Tight and Specific Responses of Fission Yeast fbp1 Gene Transcription" Biomolecules 12, no. 11: 1642. https://doi.org/10.3390/biom12111642
APA StyleAsada, R., & Hirota, K. (2022). Multi-Layered Regulations on the Chromatin Architectures: Establishing the Tight and Specific Responses of Fission Yeast fbp1 Gene Transcription. Biomolecules, 12(11), 1642. https://doi.org/10.3390/biom12111642