
Transplantation of Human Umbilical Cord Mesenchymal Stem Cells-Derived Neural Stem Cells Pretreated with Neuregulin1β Ameliorate Cerebral Ischemic Reperfusion Injury in Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Middle Cerebral Artery Occlusion/Reperfusion (MCAO/R) Model
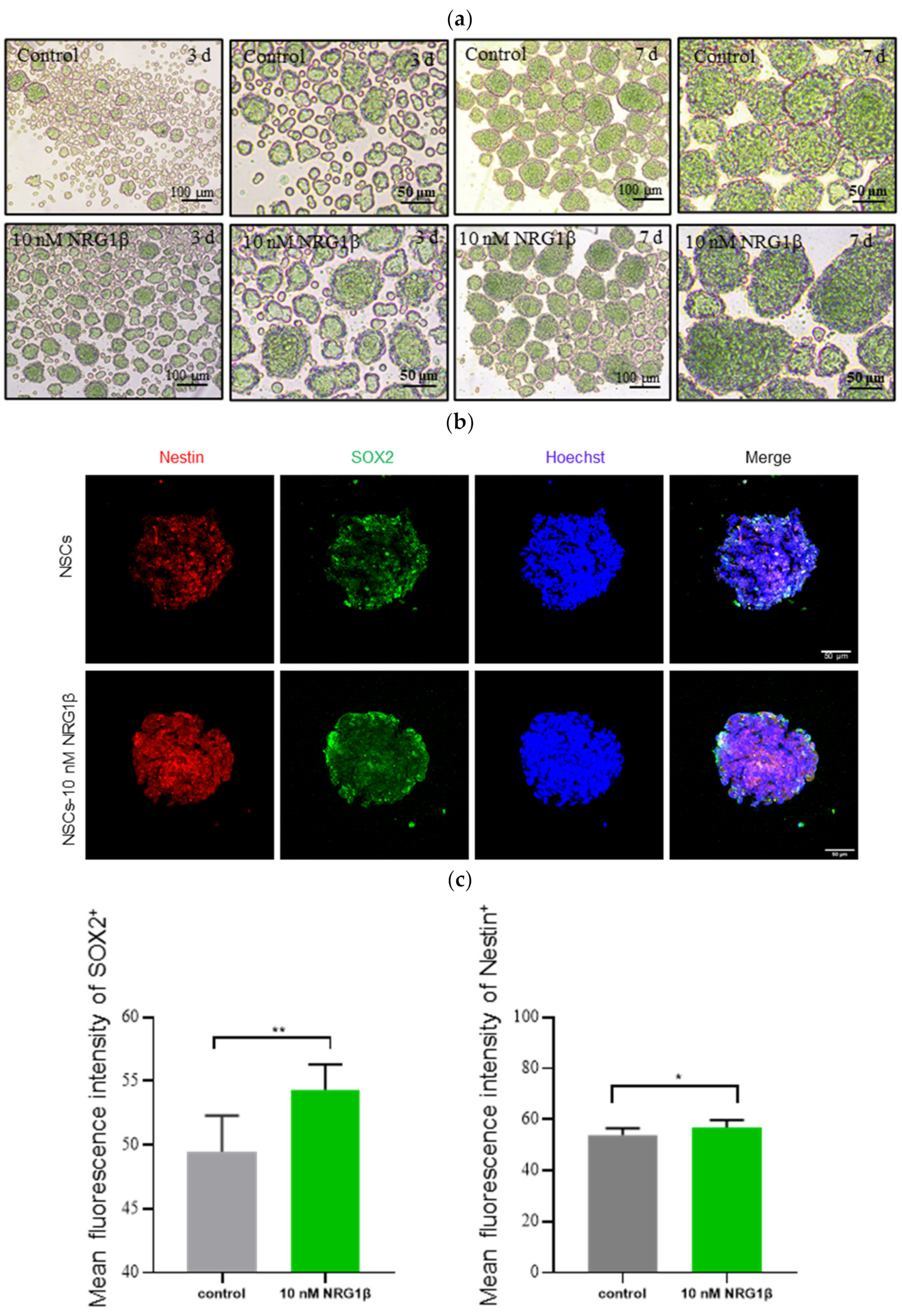
2.2. Preparation of NSCs and Immunofluorescence
2.3. Cells Transplantation
2.4. Behavioral Tests
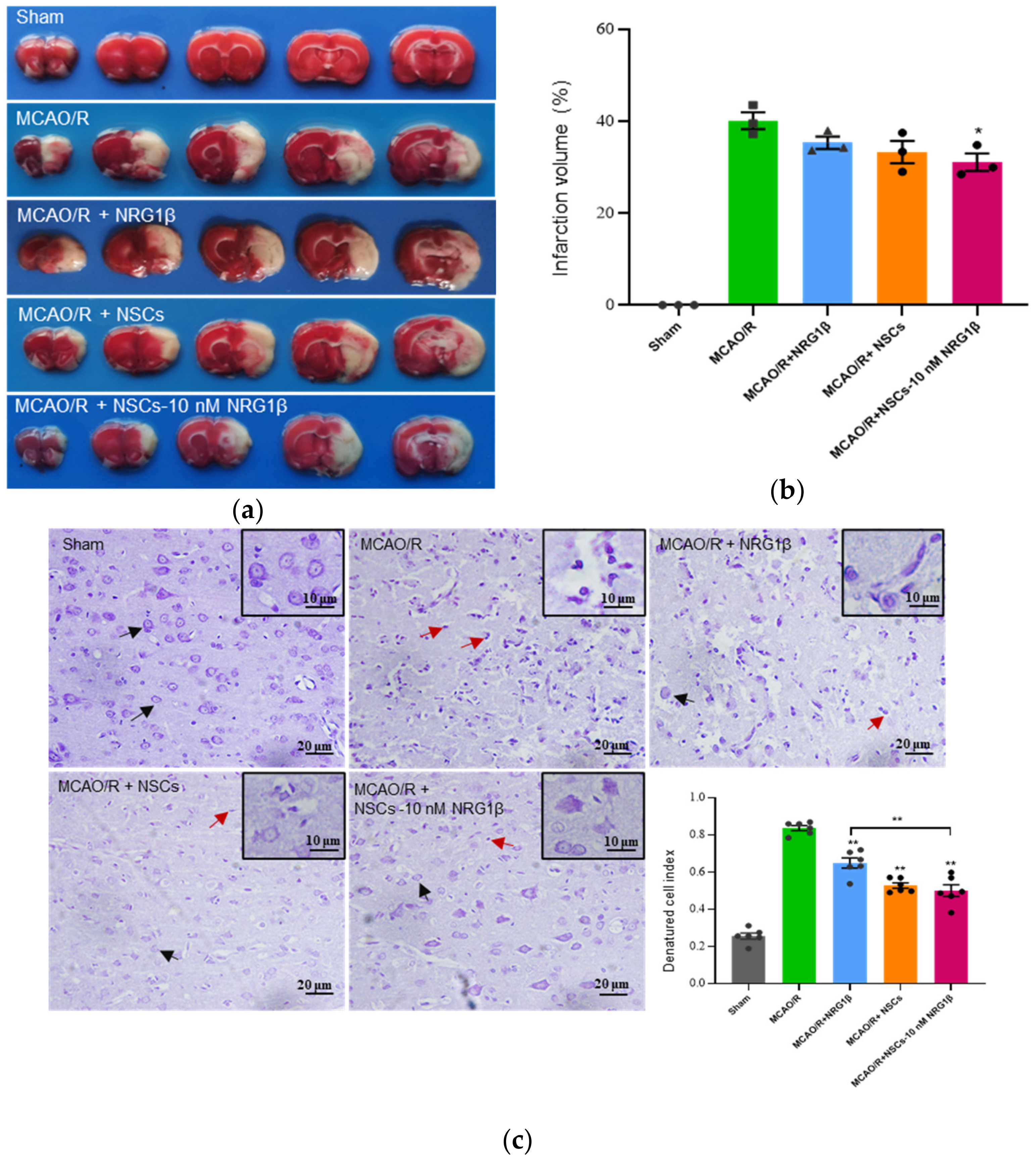
2.5. Triphenyltetrazolium Chloride (TTC) Staining and Analysis of Infarct Size
2.6. Nissl Staining
2.7. Measurement of Ferroptosis Index in Brain Tissue
2.8. Transmission Electron Microscope (TEM)
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
3.1. Transplantation of hUC-MSCs Derived NSCs Pretreated with NRG1β Ameliorate Neurological Deficits
3.2. Transplantation of NSCs Pretreated with NRG1β DECREASE Infarct Volumes and Damage of Cortical Neuron
3.3. Transplantation of NSCs Pretreated with NRG1β Change the Expression of Ferroptosis-Related Index in the Cerebral Cortex
3.4. Transplantation of NSCs Pretreated with NRG1β Alleviates Mitochondrial Damage in Cerebral Cortex Neurons
3.5. Transplantation of NSCs Pretreated with NRG1β Affects Ferroptosis in Cerebral Cortex of Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Katan, M.; Luft, A. Global Burden of Stroke. Semin. Neurol. 2018, 38, 208–211. [Google Scholar] [CrossRef] [Green Version]
- Sun, K.; Fan, J.; Han, J. Ameliorating effects of traditional Chinese medicine preparation, Chinese materia medica and active compounds on ischemia/reperfusion-induced cerebral microcirculatory disturbances and neuron damage. Acta Pharm. Sin. B 2015, 5, 8–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catanese, L.; Tarsia, J.; Fisher, M. Acute Ischemic Stroke Therapy Overview. Circ. Res. 2017, 120, 541–558. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.E.; Saver, J.L.; Cox, M.; Liang, L.; Matsouaka, R.; Xian, Y.; Bhatt, D.L.; Fonarow, G.C.; Schwamm, L.H. Increase in Endovascular Therapy in Get with The Guidelines-Stroke After the Publication of Pivotal Trials. Circulation 2017, 136, 2303–2310. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Sapolsky, R.M.; Steinberg, G.K. Interrupting reperfusion as a stroke therapy: Ischemic postconditioning reduces infarct size after focal ischemia in rats. J. Cereb. Blood Flow Metab. 2006, 26, 1114–1121. [Google Scholar] [CrossRef] [Green Version]
- Aronowski, J.; Strong, R.; Grotta, J.C. Reperfusion injury: Demonstration of brain damage produced by reperfusion after transient focal ischemia in rats. J. Cereb. Blood Flow Metab. 1997, 17, 1048–1056. [Google Scholar] [CrossRef]
- Li, J.; Ma, X.; Yu, W.; Lou, Z.; Mu, D.; Wang, Y.; Shen, B.; Qi, S. Reperfusion promotes mitochondrial dysfunction following focal cerebral ischemia in rats. PLoS ONE 2012, 7, e46498. [Google Scholar] [CrossRef]
- Arsava, E.M.; Kim, G.M.; Oliveira-Filho, J.; Gungor, L.; Noh, H.J.; Lordelo, M.J.; Avery, R.; Maier, I.L.; Ay, H. Prediction of Early Recurrence After Acute Ischemic Stroke. JAMA Neurol. 2016, 73, 396–401. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Zhang, L. Neural stem cell therapies and hypoxic-ischemic brain injury. Prog. Neurobiol. 2019, 173, 1–17. [Google Scholar] [CrossRef]
- Chen, Q.; Li, L.; Xie, H. Research progress of different types of stem cells in treatment of ischemic stroke. Chin. J. Reparative Reconstr. Surg. 2021, 35, 111–117. [Google Scholar]
- Zhang, G.-L.; Zhu, Z.-H.; Wang, Y.-Z. Neural stem cell transplantation therapy for brain ischemic stroke: Review and perspectives. World J. Stem Cells 2019, 11, 817–830. [Google Scholar] [CrossRef]
- Burden, S.; Yarden, Y. Neuregulins and their receptors: A versatile signaling module in organogenesis and oncogenesis. Neuron. 1997, 18, 847–855. [Google Scholar] [CrossRef] [Green Version]
- Ritch, P.S.; Carroll, S.L.; Sontheimer, H. Neuregulin-1 enhances survival of human astrocytic glioma cells. Glia 2005, 51, 217–228. [Google Scholar] [CrossRef] [Green Version]
- Bublil, E.M.; Yarden, Y. The EGF receptor family: Spearheading a merger of signaling and therapeutics. Curr. Opin. Cell Biol. 2007, 19, 124–134. [Google Scholar] [CrossRef]
- Mei, L.; Xiong, W.C. Neuregulin 1 in neural development, synaptic plasticity and schizophrenia. Nat. Rev. Neurosci. 2008, 9, 437–452. [Google Scholar] [CrossRef]
- Rosler, T.W.; Depboylu, C.; Arias-Carrion, O.; Wozny, W.; Carlsson, T.; Hollerhage, M.; Oertel, W.H.; Schrattenholz, A.; Hoglinger, G.U. Biodistribution and brain permeability of the extracellular domain of neuregulin-1-beta1. Neuropharmacology 2011, 61, 1413–1418. [Google Scholar] [CrossRef]
- Xu, Z.; Croslan, D.R.; Harris, A.E.; Ford, G.D.; Ford, B.D. Extended therapeutic window and functional recovery after intraarterial administration of neuregulin-1 after focal ischemic stroke. J. Cereb. Blood Flow Metab. 2006, 26, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Carlsson, T.; Schindler, F.R.; Hollerhage, M.; Depboylu, C.; Arias-Carrion, O.; Schnurrbusch, S.; Rosler, T.W.; Wozny, W.; Schwall, G.P.; Groebe, K.; et al. Systemic administration of neuregulin-1beta1 protects dopaminergic neurons in a mouse model of Parkinson’s disease. J. Neurochem. 2011, 117, 1066–1074. [Google Scholar] [CrossRef]
- Jiang, Q.; Chen, S.; Hu, C.; Huang, P.; Shen, H.; Zhao, W. Neuregulin-1 (Nrg1) signaling has a preventive role and is altered in the frontal cortex under the pathological conditions of Alzheimer’s disease. Mol. Med. Rep. 2016, 14, 2614–2624. [Google Scholar] [CrossRef] [Green Version]
- Carlos, C.J.; Mingli, L.; Adriana, H.; Annette, N.; John, O.; Watson, C.H.; K, B.P.; Wesley, S.; Precious, A.; Sri, K.; et al. Neuregulin in Health and Disease. Int. J. Brain Disord. Treat. 2018, 4, 24. [Google Scholar] [CrossRef]
- Xie, Y.; Hou, W.; Song, X.; Yu, Y.; Huang, J.; Sun, X.; Kang, R.; Tang, D. Ferroptosis: Process and function. Cell Death Differ. 2016, 23, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Doll, S.; Proneth, B.; Tyurina, Y.Y.; Panzilius, E.; Kobayashi, S.; Ingold, I.; Irmler, M.; Beckers, J.; Aichler, M.; Walch, A.; et al. ACSL4 dictates ferroptosis sensitivity by shaping cellular lipid composition. Nat. Chem. Biol. 2017, 13, 91–98. [Google Scholar] [CrossRef]
- Liu, J.; Guo, Z.N.; Yan, X.L.; Huang, S.; Ren, J.X.; Luo, Y.; Yang, Y. Crosstalk Between Autophagy and Ferroptosis and Its Putative Role in Ischemic Stroke. Front. Cell Neurosci. 2020, 14, 577403. [Google Scholar] [CrossRef]
- Guan, X.; Li, X.; Yang, X.; Yan, J.; Shi, P.; Ba, L.; Cao, Y.; Wang, P. The neuroprotective effects of carvacrol on ischemia/reperfusion-induced hippocampal neuronal impairment by ferroptosis mitigation. Life Sci. 2019, 235, 116795. [Google Scholar] [CrossRef]
- Lan, B.; Ge, J.W.; Cheng, S.W.; Zheng, X.L.; Liao, J.; He, C.; Rao, Z.Q.; Wang, G.Z. Extract of Naotaifang, a compound Chinese herbal medicine, protects neuron ferroptosis induced by acute cerebral ischemia in rats. J. Integr. Med. 2020, 18, 344–350. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, C.; Ji, Y.; Teng, L.; Guo, Y. Neuregulin-1beta plays a neuroprotective role by inhibiting the Cdk5 signaling pathway after cerebral ischemia-reperfusion injury in rats. J. Mol. Neurosci. 2018, 66, 261–272. [Google Scholar] [CrossRef]
- Gerecke, K.M.; Wyss, J.M.; Carroll, S.L. Neuregulin-1beta induces neurite extension and arborization in cultured hippocampal neurons. Mol. Cell Neurosci. 2004, 27, 379–393. [Google Scholar] [CrossRef]
- Ryu, J.; Hong, B.H.; Kim, Y.J.; Yang, E.J.; Choi, M.; Kim, H.; Ahn, S.; Baik, T.K.; Woo, R.S.; Kim, H.S. Neuregulin-1 attenuates cognitive function impairments in a transgenic mouse model of Alzheimer’s disease. Cell Death Dis. 2016, 7, e2117. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.Y.; Yoo, J.Y.; Kim, H.B.; Baik, T.K.; Lee, J.H.; Woo, R.S. Neuregulin-1 Protects Neuronal Cells Against Damage due to CoCl2-Induced Hypoxia by Suppressing Hypoxia-Inducible Factor-1alpha and P53 in SH-SY5Y Cells. Int. Neurourol. J. 2019, 23 (Suppl. S2), S111–S118. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.H.; Kim, G.H.; Lee, J.S.; Joo, I.S.; Suh-Kim, H.; Kim, S.S.; Hong, J.M. Comparison of MSC-Neurogenin1 administration modality in MCAO rat model. Transl. Neurosci. 2016, 7, 164–172. [Google Scholar] [CrossRef]
- Chua, J.Y.; Pendharkar, A.V.; Wang, N.; Choi, R.; Andres, R.H.; Gaeta, X.; Zhang, J.; Moseley, M.E.; Guzman, R. Intra-arterial injection of neural stem cells using a microneedle technique does not cause microembolic strokes. J. Cereb. Blood Flow Metab. 2011, 31, 1263–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Joseph, B.; Saatman, K.E.; Chen, L. Intra-Arterial Delivery of Neural Stem Cells to the Rat and Mouse Brain: Application to Cerebral Ischemia. J. Vis. Exp. 2020, 160, e61119. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Sanberg, P.R.; Li, Y.; Wang, L.; Lu, M.; Willing, A.E.; Sanchez-Ramos, J.; Chopp, M. Intravenous administration of human umbilical cord blood reduces behavioral deficits after stroke in rats. Stroke 2001, 32, 2682–2688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yavagal, D.R.; Lin, B.; Raval, A.P.; Garza, P.S.; Dong, C.; Zhao, W.; Rangel, E.B.; McNiece, I.; Rundek, T.; Sacco, R.L.; et al. Efficacy and dose-dependent safety of intra-arterial delivery of mesenchymal stem cells in a rodent stroke model. PLoS ONE 2014, 9, e93735. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.; Choi, J.; Kim, M. Combining Human Umbilical Cord Blood Cells with Erythropoietin Enhances Angiogenesis/Neurogenesis and Behavioral Recovery After Stroke. Front. Neurol. 2019, 10, 357. [Google Scholar] [CrossRef]
- Shi, Y.H.; Zhang, X.L.; Ying, P.J.; Wu, Z.Q.; Lin, L.L.; Chen, W.; Zheng, G.Q.; Zhu, W.Z. Neuroprotective Effect of Astragaloside IV on Cerebral Ischemia/Reperfusion Injury Rats Through Sirt1/Mapt Pathway. Front. Pharmacol. 2021, 12, 639898. [Google Scholar] [CrossRef]
- Liu, X.; Ye, R.; Yan, T.; Yu, S.P.; Wei, L.; Xu, G.; Fan, X.; Jiang, Y.; Stetler, R.A.; Liu, G.; et al. Cell based therapies for ischemic stroke: From basic science to bedside. Prog. Neurobiol. 2014, 115, 92–115. [Google Scholar] [CrossRef] [Green Version]
- Kelly, S.; Bliss, T.M.; Shah, A.K.; Sun, G.H.; Ma, M.; Foo, W.C.; Masel, J.; Yenari, M.A.; Weissman, I.L.; Uchida, N.; et al. Transplanted human fetal neural stem cells survive, migrate, and differentiate in ischemic rat cerebral cortex. Proc. Natl. Acad. Sci. USA 2004, 101, 11839–11844. [Google Scholar] [CrossRef] [Green Version]
- Boese, A.C.; Le, Q.E.; Pham, D.; Hamblin, M.H.; Lee, J.P. Neural stem cell therapy for subacute and chronic ischemic stroke. Stem. Cell Res. Ther. 2018, 9, 154. [Google Scholar] [CrossRef]
- Mukai, T.; Nagamura-Inoue, T.; Shimazu, T.; Mori, Y.; Takahashi, A.; Tsunoda, H.; Yamaguchi, S.; Tojo, A. Neurosphere formation enhances the neurogenic differentiation potential and migratory ability of umbilical cord-mesenchymal stromal cells. Cytotherapy 2016, 18, 229–241. [Google Scholar] [CrossRef]
- Feng, N.; Han, Q.; Li, J.; Wang, S.; Li, H.; Yao, X.; Zhao, R.C. Generation of highly purified neural stem cells from human adipose-derived mesenchymal stem cells by Sox1 activation. Stem. Cells Dev. 2014, 23, 515–529. [Google Scholar] [CrossRef] [Green Version]
- Rismanchi, N.; Floyd, C.L.; Berman, R.F.; Lyeth, B.G. Cell death and long-term maintenance of neuron-like state after differentiation of rat bone marrow stromal cells: A comparison of protocols. Brain Res. 2003, 991, 46–55. [Google Scholar] [CrossRef]
- Chu, K.; Park, K.I.; Lee, S.T.; Jung, K.H.; Ko, S.Y.; Kang, L.; Sinn, D.I.; Lee, Y.S.; Kim, S.U.; Kim, M.; et al. Combined treatment of vascular endothelial growth factor and human neural stem cells in experimental focal cerebral ischemia. Neurosci. Res. 2005, 53, 384–390. [Google Scholar] [CrossRef]
- Zhang, G.; Guo, X.; Chen, L.; Li, B.; Gu, B.; Wang, H.; Wu, G.; Kong, J.; Chen, W.; Yu, Y. Interferon-gamma promotes neuronal repair by transplanted neural stem cells in ischemic rats. Stem. Cells Dev. 2018, 27, 355–366. [Google Scholar] [CrossRef]
- Li, J.; Tang, Y.; Wang, Y.; Tang, R.; Jiang, W.; Yang, G.Y.; Gao, W.Q. Neurovascular recovery via co-transplanted neural and vascular progenitors leads to improved functional restoration after ischemic stroke in rats. Stem. Cell Rep. 2014, 3, 101–114. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Liao, J.; Mei, Z.; Liu, X.; Ge, J. Insight into Crosstalk between Ferroptosis and Necroptosis: Novel Therapeutics in Ischemic Stroke. Oxid. Med. Cell Longev. 2021, 2021, 9991001. [Google Scholar] [CrossRef]
- Cao, J.Y.; Dixon, S.J. Mechanisms of ferroptosis. Cell. Mol. Life Sci. 2016, 73, 2195–2209. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Liu, J.; Huang, Y.; Tang, X.; Xiao, H.; Hu, Z. Oxidative Stress, Inflammation, and Autophagy: Potential Targets of Mesenchymal Stem Cells-Based Therapies in Ischemic Stroke. Front. Neurosci. 2021, 15, 641157. [Google Scholar] [CrossRef]
- Maiorino, M.; Conrad, M.; Ursini, F. GPx4, Lipid Peroxidation, and Cell Death: Discoveries, Rediscoveries, and Open Issues. Antioxid. Redox. Signal 2018, 29, 61–74. [Google Scholar] [CrossRef]
- Murphy, M.E. Ironing out how p53 regulates ferroptosis. Proc. Natl. Acad. Sci. USA 2016, 113, 12350–12352. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhang, C.; Wang, J.; Hu, W.; Feng, Z. The Regulation of Ferroptosis by Tumor Suppressor p53 and its Pathway. Int. J. Mol. Sci. 2020, 21, 8387. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Scores | |
|---|---|---|
| Raising rat by tail | Flexion of forelimb | 1 |
| Flexion of hindlimb | 1 | |
| Head moved >10° to vertical axis within 30 s | 1 | |
| Placing rat on floor (normal = 0, maximum = 3) | Normal walk | 0 |
| Inability to walk straight | 1 | |
| Circling toward paretic side | 2 | |
| Falls to paretic side | 3 | |
| Sensory tests | Placing test (visual and tactile test) | 1 |
| Proprioceptive test (deep sensation, pushing paw against table edge to stimulate limb muscles) | 1 | |
| Beam balance tests (normal = 0; maximum = 6) | Balances with steady posture | 0 |
| Grasps side of beam | 1 | |
| Hugs beam and one limb fall from beam | 2 | |
| Hugs beam and two limbs fall from beam, or spins on beam (>60 s) | 3 | |
| Attempts to balance on beam but falls off (>40 s) | 4 | |
| Attempts to balance on beam but falls off (>20 s) | 5 | |
| Falls off; no attempt to balance or hang on to beam (<20 s) | 6 | |
| Reflex absence and abnormal movements | Pinna reflex (head shake when auditory meatus is touched) | 1 |
| Corneal reflex (eye blink when cornea is lightly touched with cotton) | 1 | |
| Startle reflex (motor response to a brief noise from snapping a clipboard paper) | 1 | |
| Seizures, myoclonus, myodystonia | 1 | |
| Antibody | Vendor | Cat. No | Dilution | Applications |
|---|---|---|---|---|
| ACTIN | Proteintech | 20536-1-AP | 1:1000 | Western blot |
| GPX4 | ZENBIO | 381958 | 1:1000 | Western blot |
| SLC7A11 | ABclonal | A15604 | 1:1000 | Western blot |
| p53 | ABclonal | A3185 | 1:1000 | Western blot |
| TFR1 | ABclonal | A10231 | 1:1000 | Western blot |
| DMT1 | ABclonal | A5865 | 1:1000 | Western blot |
| Sham | MCAO/R | MCAO/R + NRG1β | MCAO/R + NSCs | MCAO/R + NSCs- 10 nM NRG1β | |
|---|---|---|---|---|---|
| Day 0 | 0.00 ± 0.00 | 11.00 ± 0.76 | 10.88 ± 0.83 | 10.88 ± 0.64 | 10.75 ± 0.71 |
| Day 1 | 0.00 ± 0.00 | 10.00 ± 0.76 | 9.13 ± 0.64 | 8.63 ± 0.74 | 8.50 ± 1.07 |
| Day 2 | 0.00 ± 0.00 | 9.63 ± 0.52 | 8.88 ± 0.64 | 8.37 ± 0.52 | 8.00 ± 1.20 |
| Day 3 | 0.00 ± 0.00 | 9.38 ± 0.74 | 8.75 ± 0.71 | 8.13 ± 0.83 | 7.75 ± 1.28 |
| Day 4 | 0.00 ± 0.00 | 9.00 ± 0.75 | 8.38 ± 0.74 | 7.75 ± 0.71 | 7.13 ± 0.83 |
| Day 5 | 0.00 ± 0.00 | 8.75 ± 0.71 | 7.88 ± 0.99 | 7.38 ± 0.52 | 6.88 ± 0.99 |
| Day 6 | 0.00 ± 0.00 | 8.38 ± 0.74 | 7.50 ± 0.93 | 7.13 ± 0.83 | 6.75 ± 0.89 |
| Day 7 | 0.00 ± 0.00 | 8.25 ±0.71 | 7.38 ± 1.06 | 6.88 ± 0.64 | 6.50 ± 0.76 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, Q.-Y.; Ren, Y.-Q.; Ni, Q.-S.; Song, Z.-H.; Ge, K.-L.; Guo, Y.-L. Transplantation of Human Umbilical Cord Mesenchymal Stem Cells-Derived Neural Stem Cells Pretreated with Neuregulin1β Ameliorate Cerebral Ischemic Reperfusion Injury in Rats. Biomolecules 2022, 12, 428. https://doi.org/10.3390/biom12030428
Zhai Q-Y, Ren Y-Q, Ni Q-S, Song Z-H, Ge K-L, Guo Y-L. Transplantation of Human Umbilical Cord Mesenchymal Stem Cells-Derived Neural Stem Cells Pretreated with Neuregulin1β Ameliorate Cerebral Ischemic Reperfusion Injury in Rats. Biomolecules. 2022; 12(3):428. https://doi.org/10.3390/biom12030428
Chicago/Turabian StyleZhai, Qiu-Yue, Yu-Qian Ren, Qin-Shuai Ni, Zhen-Hua Song, Ke-Li Ge, and Yun-Liang Guo. 2022. "Transplantation of Human Umbilical Cord Mesenchymal Stem Cells-Derived Neural Stem Cells Pretreated with Neuregulin1β Ameliorate Cerebral Ischemic Reperfusion Injury in Rats" Biomolecules 12, no. 3: 428. https://doi.org/10.3390/biom12030428