
Exosomes Derived from BM-MSCs Mitigate the Development of Chronic Kidney Damage Post-Menopause via Interfering with Fibrosis and Apoptosis

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
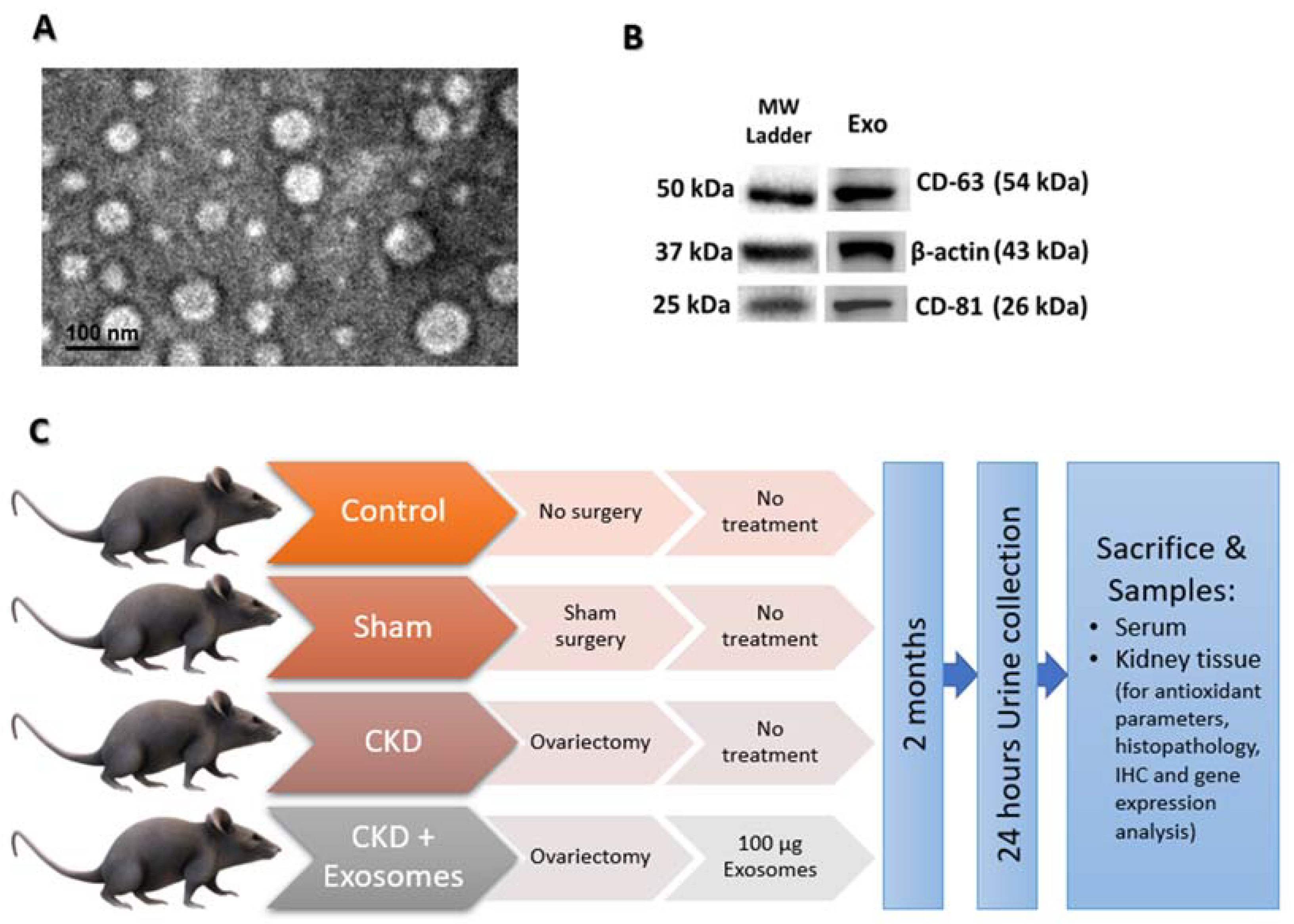
2.1. Study Animals and Experimental Design
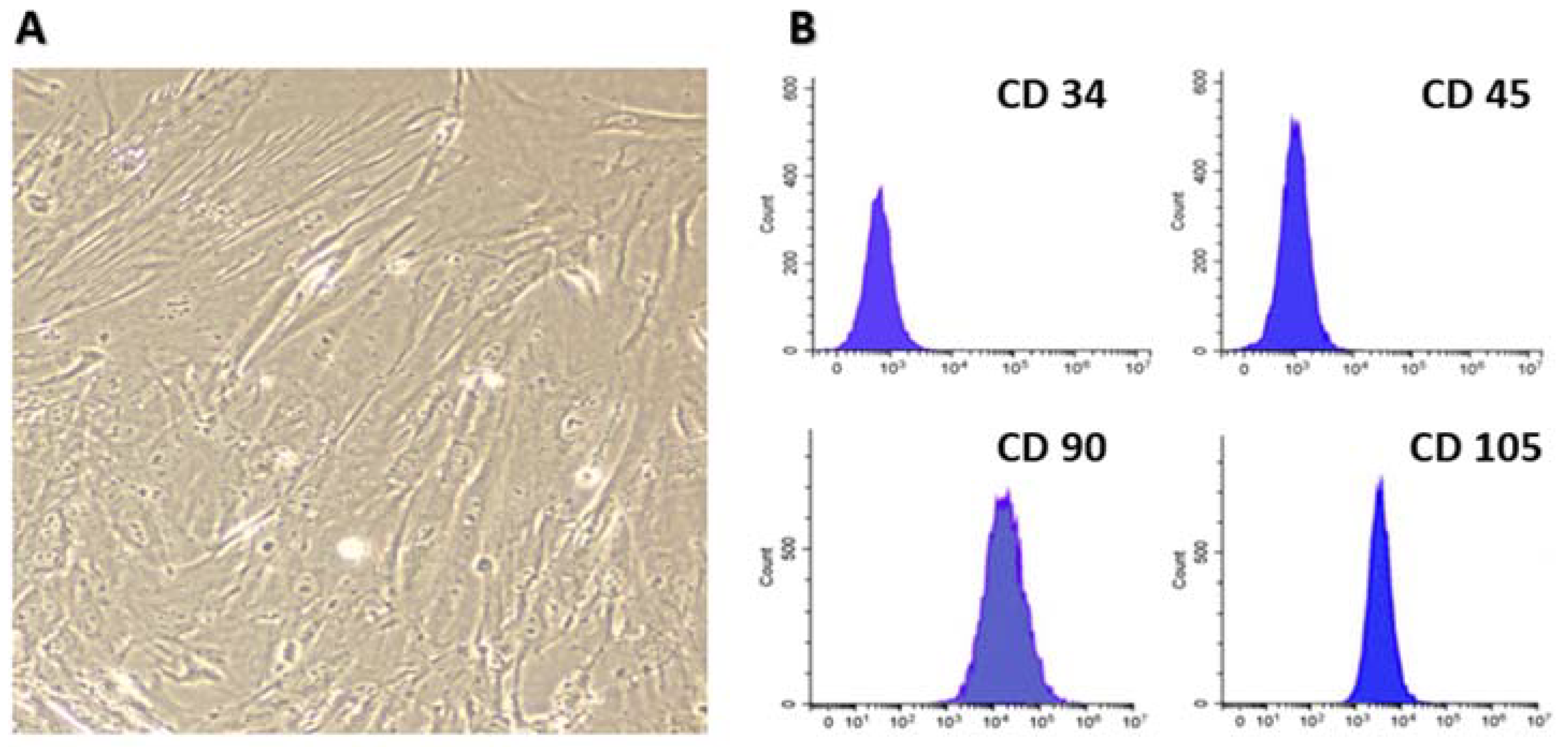
2.2. Isolation and Characterization of Bone Marrow MSCs
2.3. Isolation and Characterization of Exosomes from BM-MSCs
2.4. Post-Menopause Chronic Kidney Disease (CKD) Model
2.5. Urine Analyses
2.6. Administration of Exosomes Derived from Bone Marrow MSCs
2.7. Serum Biochemical Assays and Kidney Tissue Antioxidants Detection
2.8. Histopathologic Examination
2.9. Immunohistochemistry of Caspase 3
2.10. Real Time qPCR
2.11. Statistical Analyses
3. Results
3.1. BM-MSCs Isolation and Collection of Their Exosomes
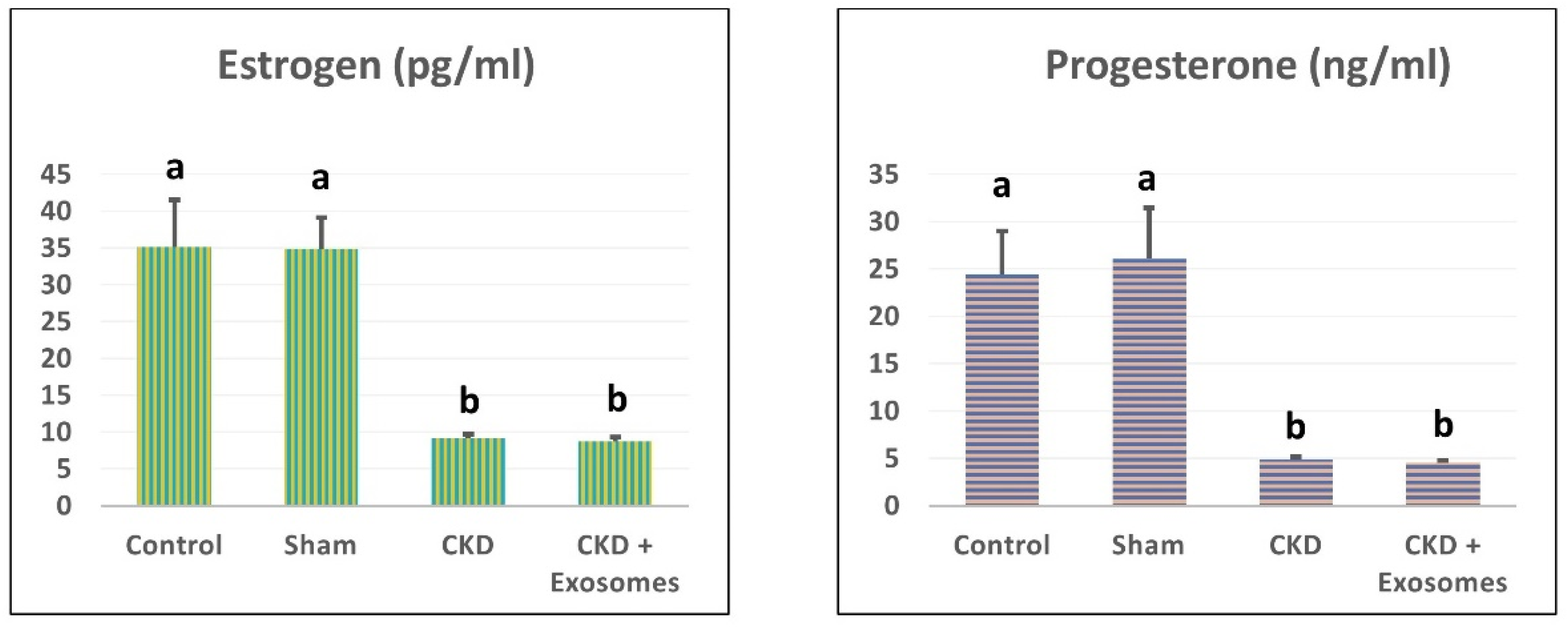
3.2. Changes in Serum Estrogen and Progesterone Levels in Study Rats
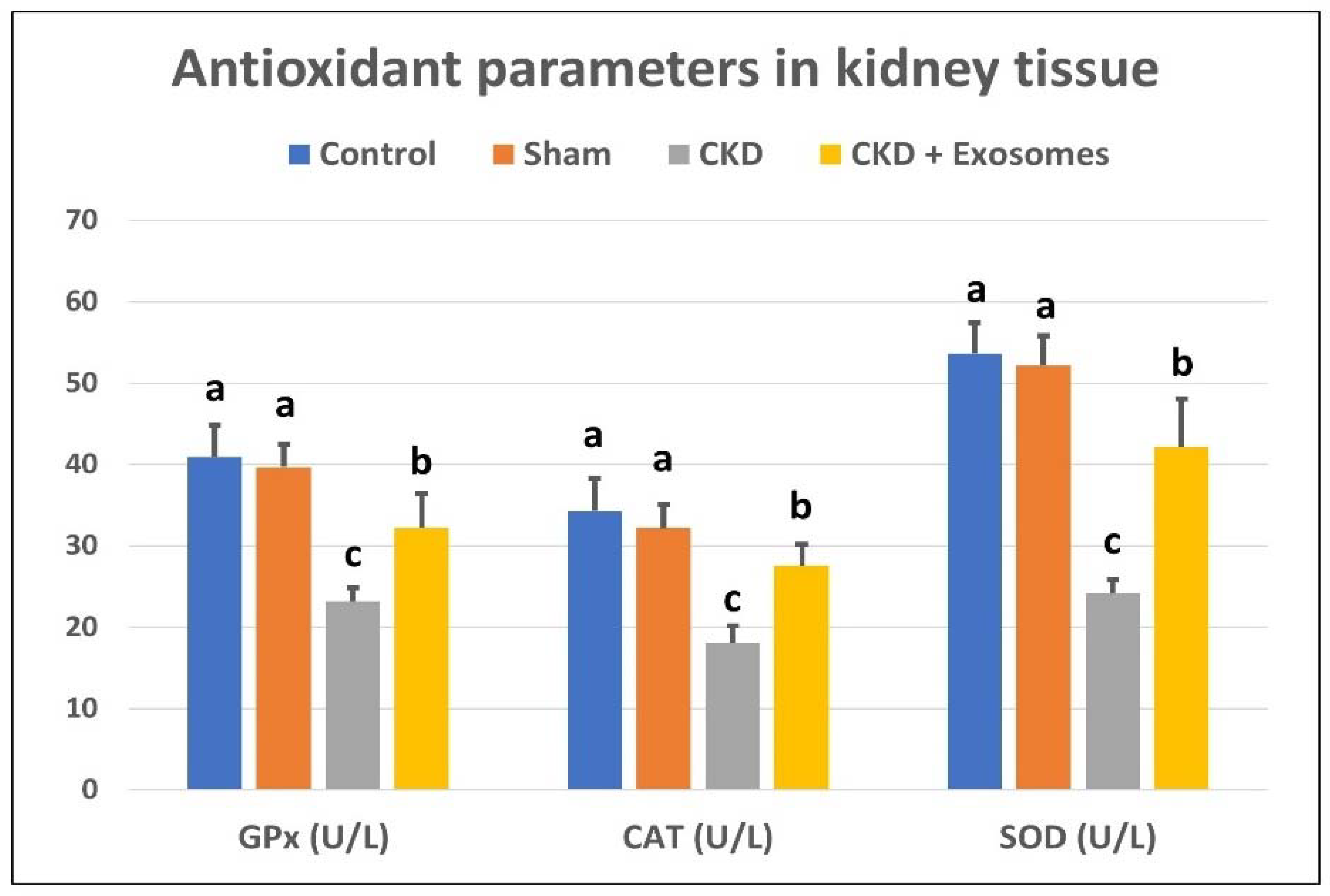
3.3. Changes in Serum/Tissue Biochemical Parameters
3.4. Changes in GFR and Urinary Protein Excretion
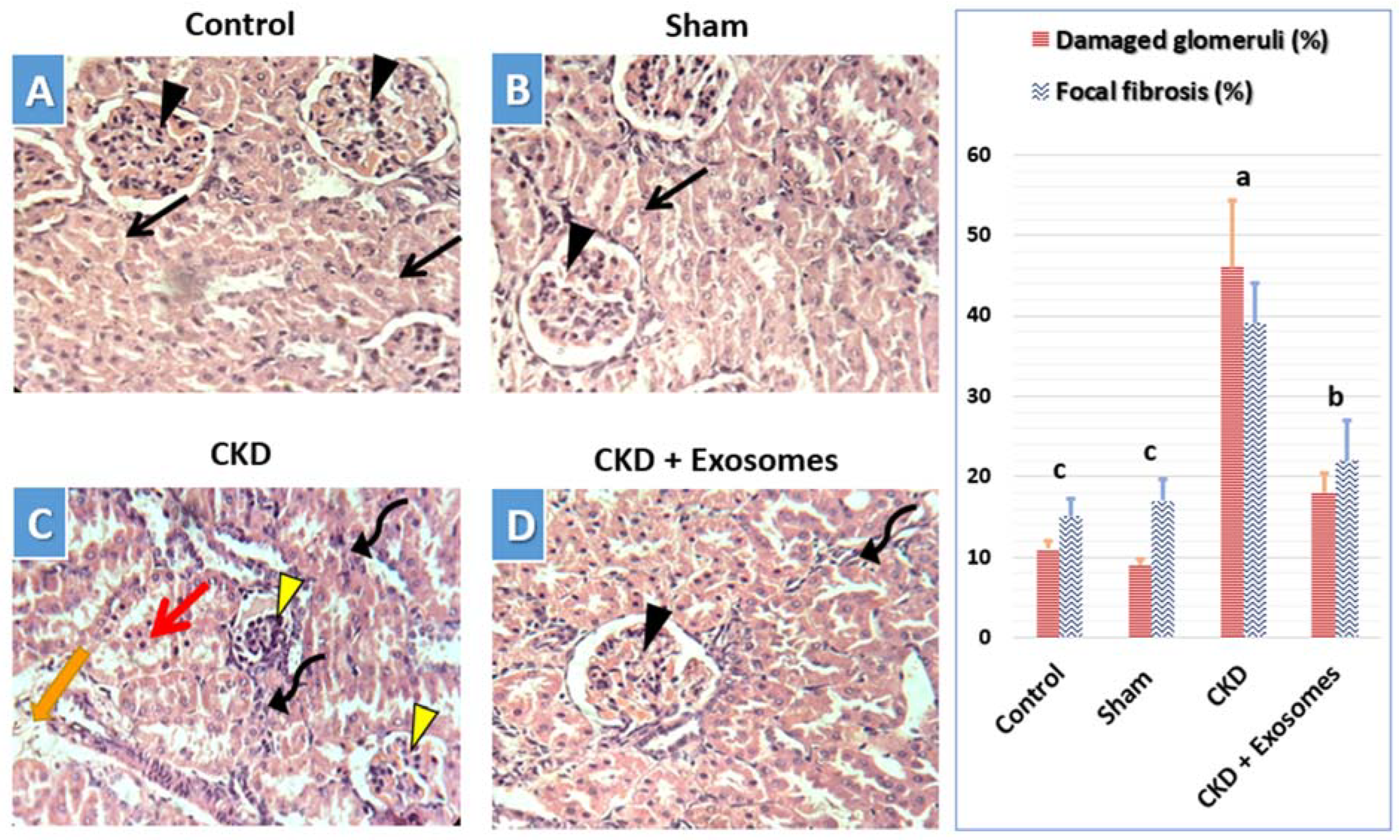
3.5. Histological Changes in Kidney Tissue
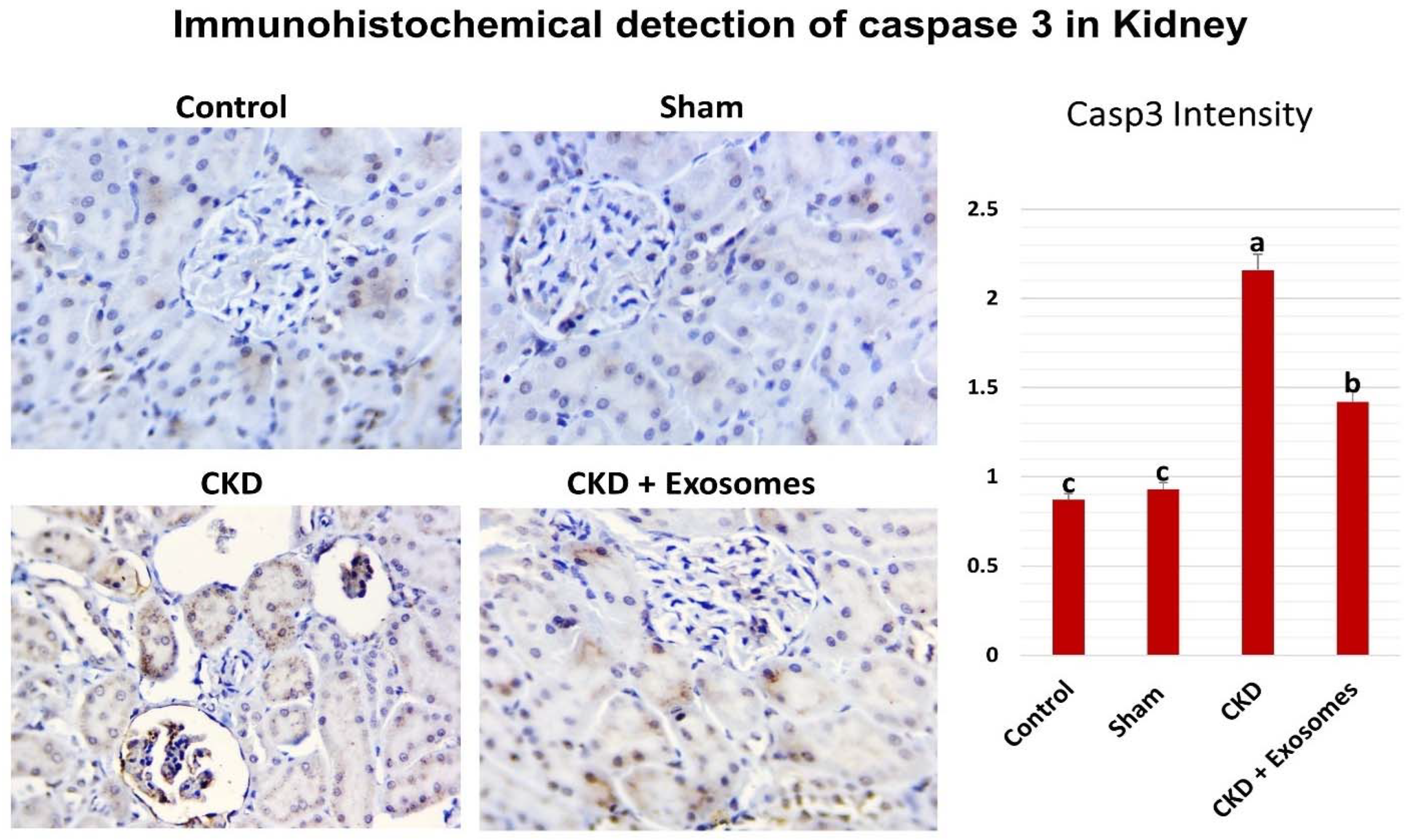
3.6. Detection of Caspase 3 Expression in the Kidney by Immunohistochemistry
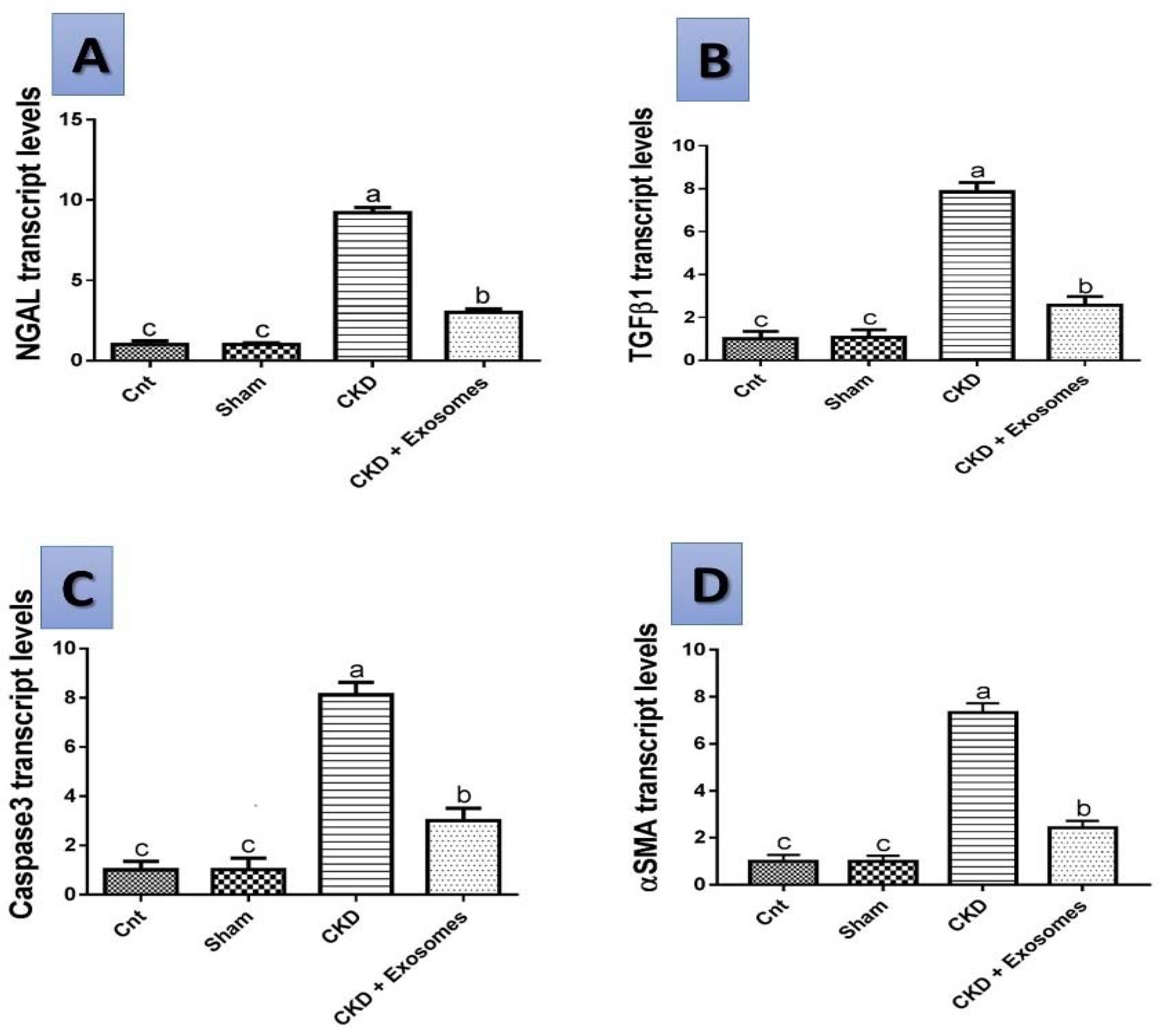
3.7. Gene Expression Results by qPCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hill, N.R.; Fatoba, S.T.; Oke, J.L.; Hirst, J.A.; O’Callaghan, C.A.; Lasserson, D.S.; Hobbs, F.D.R. Global prevalence of chronic kidney disease—A systematic review and meta-analysis. PLoS ONE 2016, 11, e0158765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Kondo, K. Chronic kidney disease in postmenopausal women. Hypertens. Res. 2012, 35, 142–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.M.; Tang, P.M.K.; Li, J.; Lan, H.Y. TGF-ß/Smad signaling in renal fibrosis. Front. Physiol. 2015, 6, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Yang, T.; Lu, D.W.; Zhao, H.; Feng, Y.L.; Chen, H.; Chen, D.Q.; Vaziri, N.D.; Zhao, Y.Y. Central role of dysregulation of TGF-β/Smad in CKD progression and potential targets of its treatment. Biomed. Pharm. 2018, 101, 670–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagita, M. Inhibitors/antagonists of TGF-β system in kidney fibrosis. Nephrol. Dial. Transpl. 2012, 27, 3686–3691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiyama, H.; Kashihara, N.; Makino, H.; Yamasaki, Y.; Ota, Z. Apoptosis in glomerular sclerosis. Kidney Int. 1996, 49, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Thomas, G.L.; Yang, B.; Wagner, B.E.; Savill, J.; el Nahas, A.M. Cellular apoptosis and proliferation in experimental renal fibrosis. Nephrol. Dial. Transpl. 1998, 13, 2216–2226. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; el Nahas, A.M.; Thomas, G.L.; Haylor, J.L.; Watson, P.F.; Wagner, B.; Johnson, T.S. Caspase-3 and apoptosis in experimental chronic renal scarring. Kidney Int. 2001, 60, 1765–1776. [Google Scholar] [CrossRef] [Green Version]
- Michels, K.B.; Manson, J.E. Postmenopausal Hormone Therapy in the 21st Century: Reconciling Findings from Observational Studies and Randomized Clinical Trials, 3rd ed.; Lobo, R.A., Ed.; Treat. Postmenopausal Woman; Academic Press: New York, NY, USA, 2007; pp. 619–626. [Google Scholar] [CrossRef]
- El-Gendy, A.A.; Elsaed, W.M.; Abdallah, H.I. Potential role of estradiol in ovariectomy-induced derangement of renal endocrine functions. Ren. Fail. 2019, 41, 507–520. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, M.A.; Hutson, D.D.; Trimmer, E.H.; Kashyap, S.N.; Duong, J.L.; Murphy, B.; Grissom, E.M.; Daniel, J.M.; Lindsey, S.H. Long- but not short-term estradiol treatment induces renal damage in midlife ovariectomized Long-Evans rats. Am. J. Physiol. Ren. Physiol. 2017, 312, F305–F311. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.S.; Wong, V.W.; Gurtner, G.C. Therapeutic potential of bone marrow-derived mesenchymal stem cells for cutaneous wound healing. Front. Immunol. 2012, 3, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biancone, L.; Bruno, S.; Deregibus, M.C.; Tetta, C.; Camussi, G. Therapeutic potential of mesenchymal stem cell-derived microvesicles. Nephrol. Dial. Transpl. 2012, 27, 3037–3042. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.M.; Shah, J.; Srivastava, A.S. Therapeutic potential of mesenchymal stem cells in regenerative medicine. Stem Cells Int. 2013, 2013, 496218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simons, M.; Raposo, G. Exosomes—Vesicular carriers for intercellular communication. Curr. Opin. Cell Biol. 2009, 21, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, A.K.; Giebel, B. Exosomes: Small vesicles participating in intercellular communication. Int. J. Biochem. Cell Biol. 2012, 44, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Murray, L.M.A.; Krasnodembskaya, A.D. Concise Review: Intercellular Communication Via Organelle Transfer in the Biology and Therapeutic Applications of Stem Cells. Stem Cells 2019, 37, 14–25. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; el Andaloussi, S.; Wood, M.J.A. Exosomes and microvesicles: Extracellular vesicles for genetic information transfer and gene therapy. Hum. Mol. Genet. 2012, 21, R125–R134. [Google Scholar] [CrossRef] [Green Version]
- Bruno, S.; Chiabotto, G.; Camussi, G. Concise Review: Different Mesenchymal Stromal/Stem Cell Populations Reside in the Adult Kidney. Stem Cells Transl. Med. 2014, 3, 1451–1455. [Google Scholar] [CrossRef]
- Klinkhammer, B.M.; Kramann, R.; Mallau, M.; Makowska, A.; van Roeyen, C.R.; Rong, S.; Buecher, E.B.; Boor, P.; Kovacova, K.; Zok, S.; et al. Mesenchymal Stem Cells from Rats with Chronic Kidney Disease Exhibit Premature Senescence and Loss of Regenerative Potential. PLoS ONE 2014, 9, e92115. [Google Scholar] [CrossRef] [Green Version]
- Lin, K.C.; Yip, H.K.; Shao, P.L.; Wu, S.C.; Chen, K.H.; Chen, Y.T.; Yang, C.C.; Sun, C.K.; Kao, G.S.; Chen, S.Y.; et al. Combination of adipose-derived mesenchymal stem cells (ADMSC) and ADMSC-derived exosomes for protecting kidney from acute ischemia-reperfusion injury. Int. J. Cardiol. 2016, 216, 173–185. [Google Scholar] [CrossRef]
- Cao, J.; Wang, B.; Tang, T.; Lv, L.; Ding, Z.; Li, Z.; Hu, R.; Wei, Q.; Shen, A.; Fu, Y.; et al. Three-dimensional culture of MSCs produces exosomes with improved yield and enhanced therapeutic efficacy for cisplatin-induced acute kidney injury. Stem Cell Res. Ther. 2020, 11, 206. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Ha, D.H.; Go, H.K.; Youn, J.; Kim, H.K.; Jin, R.C.; Miller, R.B.; Kim, D.H.; Cho, B.S.; Yi, Y.W. Reproducible Large-Scale Isolation of Exosomes from Adipose Tissue-Derived Mesenchymal Stem/Stromal Cells and Their Application in Acute Kidney Injury. Int. J. Mol. Sci. 2020, 21, 4774. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Zuo, B.; Wang, Y.; Li, S.; Yang, J.; Sun, D. Protective function of exosomes from adipose tissue-derived mesenchymal stem cells in acute kidney injury through SIRT1 pathway. Life Sci. 2020, 255, 117719. [Google Scholar] [CrossRef] [PubMed]
- Nowak, N.; Yamanouchi, M.; Satake, E. The Nephroprotective Properties of Extracellular Vesicles in Experimental Models of Chronic Kidney Disease: A Systematic Review. Stem Cell Rev. Rep. 2022, 18, 902–932. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, M.; Nadri, S. A protocol for isolation and culture of mesenchymal stem cells from mouse bone marrow. Nat. Protoc. 2009, 4, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, F.A.; El-Magd, M.A.; Abdelfattah-Hassan, A.; Saleh, A.A.; Saadeldin, I.M.; El-Shetry, E.S.; Badawy, A.A.; Alkarim, S. Potential Effect of Exosomes Derived from Cancer Stem Cells and MSCs on Progression of DEN-Induced HCC in Rats. Stem Cells Int. 2018, 2018, 8058979. [Google Scholar] [CrossRef] [Green Version]
- el Nashar, E.M.; Alghamdi, M.A.; Alasmari, W.A.; Hussein, M.M.A.; Hamza, E.; Taha, R.I.; Ahmed, M.M.; Al-Khater, K.M.; Abdelfattah-Hassan, A. Autophagy Promotes the Survival of Adipose Mesenchymal Stem/Stromal Cells and Enhances Their Therapeutic Effects in Cisplatin-Induced Liver Injury via Modulating TGF-β1/Smad and PI3K/AKT Signaling Pathways. Cells 2021, 10, 2475. [Google Scholar] [CrossRef]
- Lobb, R.J.; Becker, M.; Wen, S.W.; Wong, C.S.F.; Wiegmans, A.P.; Leimgruber, A.; Möller, A. Optimized exosome isolation protocol for cell culture supernatant and human plasma. J. Extracell. Vesicles 2015, 4, 27031. [Google Scholar] [CrossRef]
- Steele, M.S.; Bennett, R.A. Clinical Technique: Dorsal Ovariectomy in Rodents. J. Exot. Pet Med. 2011, 20, 222–226. [Google Scholar] [CrossRef]
- Vorland, C.J.; Lachcik, P.J.; Swallow, E.A.; Metzger, C.E.; Allen, M.R.; Chen, N.X.; Moe, S.M.; Gallant, K.M.H. Effect of ovariectomy on the progression of chronic kidney disease-mineral bone disorder (CKD-MBD) in female Cy/+ rats. Sci. Rep. 2019, 9, 7936. [Google Scholar] [CrossRef]
- Ke, H.Z.; Chen, H.K.; Simmons, H.A.; Qi, H.; Crawford, D.T.; Pirie, C.M.; Chidsey-Frink, K.L.; Ma, Y.F.; Jee, W.S.S.; Thompson, D.D. Comparative effects of droloxifene, tamoxifen, and estrogen on bone, serum cholesterol, and uterine histology in the ovariectomized rat model. Bone 1997, 20, 31–39. [Google Scholar] [CrossRef]
- Abdelfattah-Hassan, A.; Shalaby, S.I.; Khater, S.I.; El-Shetry, E.S.; el Fadil, H.A.; Elsayed, S.A. Panax ginseng is superior to vitamin E as a hepatoprotector against cyclophosphamide-induced liver damage. Complement. Ther. Med. 2019, 46, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Li, Y.; Liu, M.; Li, Y.; Cong, B. Renal neutrophil gelatinase associated lipocalin expression in lipopolysaccharide-induced acute kidney injury in the rat. BMC Nephrol. 2012, 13, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, I.; Uddin, M.J.; Lee, G.; Jiang, S.; Pak, E.S.; Ha, H. Peroxiredoxin 3 deficiency accelerates chronic kidney injury in mice through interactions between macrophages and tubular epithelial cells. Free Radic. Biol. Med. 2019, 131, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Adil, M.; Kandhare, A.D.; Visnagri, A.; Bodhankar, S.L. Naringin ameliorates sodium arsenite-induced renal and hepatic toxicity in rats: Decisive role of KIM-1, Caspase-3, TGF-β, and TNF-α. Ren. Fail. 2015, 37, 1396–1407. [Google Scholar] [CrossRef] [PubMed]
- Thongboonkerd, V. Roles for exosome in various kidney diseases and disorders. Front. Pharm. 2020, 10, 1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wise, P.M.; Rainer, A. Effect of Ovariectomy on Plasma LH, FSH, Estradiol, and Progesterone and Medial Basal Hypothalamic LHRH Concentrations in Old and Young Rats. Neuroendocrinology 1980, 30, 15–19. [Google Scholar] [CrossRef]
- Wronski, T.J.; Schenk, P.A.; Cintrón, M.; Walsh, C.C. Effect of body weight on osteopenia in ovariectomized rats. Calcif. Tissue Int. 1987, 40, 155–159. [Google Scholar] [CrossRef]
- Li, L.H.; Wang, Z.C.; Yu, J.; Zhang, Y.Q. Ovariectomy Results in Variable Changes in Nociception, Mood and Depression in Adult Female Rats. PLoS ONE 2014, 9, e94312. [Google Scholar] [CrossRef] [Green Version]
- Hao, F.; Gu, Y.; Tan, X.; Deng, Y.; Wu, Z.T.; Xu, M.J.; Wang, W.Z. Estrogen Replacement Reduces Oxidative Stress in the Rostral Ventrolateral Medulla of Ovariectomized Rats. Oxid. Med. Cell. Longev. 2016, 2016, 2158971. [Google Scholar] [CrossRef] [Green Version]
- Diwan, V.; Small, D.; Kauter, K.; Gobe, G.C.; Brown, L. Gender differences in adenine-induced chronic kidney disease and cardiovascular complications in rats. Am. J. Physiol. Ren. Physiol. 2014, 307, F1169–F1178. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Gua, C.; Wu, B.; Chen, Y. Increased circulating trimethylamine N-oxide contributes to endothelial dysfunction in a rat model of chronic kidney disease. Biochem. Biophys. Res. Commun. 2018, 495, 2071–2077. [Google Scholar] [CrossRef] [PubMed]
- Nangaku, M.; Pippin, J.; Couser, W.G. C6 Mediates Chronic Progression of Tubulointerstitial Damage in Rats with Remnant Kidneys. J. Am. Soc. Nephrol. 2002, 13, 928–936. [Google Scholar] [CrossRef]
- Al Za’abi, M.; Al Salam, S.; Al Suleimani, Y.; Ashique, M.; Manoj, P.; Nemmar, A.; Ali, B.H. Effects of repeated increasing doses of cisplatin as models of acute kidney injury and chronic kidney disease in rats. Naunyn. Schmiedebergs. Arch. Pharm. 2021, 394, 249–259. [Google Scholar] [CrossRef]
- Chow, J.; Hartley, R.B.; Jagger, C.; Dilly, S.A. ICAM-1 expression in renal disease. J. Clin. Pathol. 1992, 45, 880. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.H.; Kim, H.J.; Rodriguez-Iturbe, B.; Vaziri, N.D. Niacin ameliorates oxidative stress, inflammation, proteinuria, and hypertension in rats with chronic renal failure. Am. J. Physiol. Ren. Physiol. 2009, 297, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Karadeniz, A.; Simsek, N.; Karakus, E.; Yildirim, S.; Kara, A.; Can, I.; Kisa, F.; Emre, H.; Turkeli, M. Royal jelly modulates oxidative stress and apoptosis in liver and kidneys of rats treated with cisplatin. Oxid. Med. Cell. Longev. 2011, 2011, 981793. [Google Scholar] [CrossRef] [Green Version]
- Supavekin, S.; Zhang, W.; Kucherlapati, R.; Kaskel, F.J.; Moore, L.C.; Devarajan, P. Differential gene expression following early renal ischemia/reperfusion. Kidney Int. 2003, 63, 1714–1724. [Google Scholar] [CrossRef] [Green Version]
- Bolignano, D.; Donato, V.; Coppolino, G.; Campo, S.; Buemi, A.; Lacquaniti, A.; Buemi, M. Neutrophil Gelatinase–Associated Lipocalin (NGAL) as a Marker of Kidney Damage. Am. J. Kidney Dis. 2008, 52, 595–605. [Google Scholar] [CrossRef]
- Mori, K.; Nakao, K. Neutrophil gelatinase-associated lipocalin as the real-time indicator of active kidney damage. Kidney Int. 2007, 71, 967–970. [Google Scholar] [CrossRef] [Green Version]
- Han, M.; Li, Y.; Wen, D.; Liu, M.; Ma, Y.; Cong, B. NGAL protects against endotoxin-induced renal tubular cell damage by suppressing apoptosis. BMC Nephrol. 2018, 19, 168. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Hendrickson, E.A.; Bremner, T.A.; Wyche, J.H. A Sequential Two-Step Mechanism for the Production of the Mature p17:p12 Form of Caspase-3 In Vitro. J. Biol. Chem. 1997, 272, 13432–13436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, R.; Wang, Y.; Minto, A.W.; Quigg, R.J.; Cunningham, P.N. Acute Renal Failure in Endotoxemia is Dependent on Caspase Activation. J. Am. Soc. Nephrol. 2004, 15, 3093–3102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sureshbabu, A.; Muhsin, S.A.; Choi, M.E. TGF-β signaling in the kidney: Profibrotic and protective effects. Am. J. Physiol. Ren. Physiol. 2016, 310, F596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeisberg, M.; Hanai, J.I.; Sugimoto, H.; Mammoto, T.; Charytan, D.; Strutz, F.; Kalluri, R. BMP-7 counteracts TGF-β1–induced epithelial-to-mesenchymal transition and reverses chronic renal injury. Nat. Med. 2003, 9, 964–968. [Google Scholar] [CrossRef]
- Bolati, D.; Shimizu, H.; Higashiyama, Y.; Nishijima, F.; Niwa, T. Indoxyl Sulfate Induces Epithelial-to-Mesenchymal Transition in Rat Kidneys and Human Proximal Tubular Cells. Am. J. Nephrol. 2011, 34, 318–323. [Google Scholar] [CrossRef]
- Cho, J.H.; Ryu, H.M.; Oh, E.J.; Yook, J.M.; Ahn, J.S.; Jung, H.Y.; Choi, J.Y.; Park, S.H.; Kim, Y.L.; Kwak, I.S.; et al. Alpha1-Antitrypsin Attenuates Renal Fibrosis by Inhibiting TGF-β1-Induced Epithelial Mesenchymal Transition. PLoS ONE 2016, 11, e0162186. [Google Scholar] [CrossRef]
- Xiao, Q.; Guan, Y.; Li, C.; Liu, L.; Zhao, D.; Wang, H. Decreased expression of transforming growth factor-β1 and α-smooth muscle actin contributes to the protection of lotensin against chronic renal failure in rats. Ren. Fail. 2018, 40, 583. [Google Scholar] [CrossRef] [Green Version]
- Nagaishi, K.; Mizue, Y.; Chikenji, T.; Otani, M.; Nakano, M.; Konari, N.; Fujimiya, M. Mesenchymal stem cell therapy ameliorates diabetic nephropathy via the paracrine effect of renal trophic factors including exosomes. Sci. Rep. 2016, 6, 34842. [Google Scholar] [CrossRef]
- Kuppe, C.; Kramann, R. Role of mesenchymal stem cells in kidney injury and fibrosis. Curr. Opin. Nephrol. Hypertens. 2016, 25, 372–377. [Google Scholar] [CrossRef]
- Zhuang, Q.; Ma, R.; Yin, Y.; Lan, T.; Yu, M.; Ming, Y. Mesenchymal Stem Cells in Renal Fibrosis: The Flame of Cytotherapy. Stem Cells Int. 2019, 2019, 8387350. [Google Scholar] [CrossRef] [Green Version]
- Ji, C.; Zhang, J.; Zhu, Y.; Shi, H.; Yin, S.; Sun, F.; Wang, Q.; Zhang, L.; Yan, Y.; Zhang, X.; et al. Exosomes derived from hucMSC attenuate renal fibrosis through CK1δ/β-TRCP-mediated YAP degradation. Cell Death Dis. 2020, 11, 327. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Hu, D.; Zhou, Y.; Yu, Y.; Shen, L.; Long, C.; Butnaru, D.; Timashev, P.; He, D.; Lin, T.; et al. Exosomes released by human umbilical cord mesenchymal stem cells protect against renal interstitial fibrosis through ROS-mediated P38MAPK/ERK signaling pathway. Am. J. Transl. Res. 2020, 12, 4998. [Google Scholar] [PubMed]
- Ji, C.; Zhang, J.; Zhou, Z.; Shi, H.; Liu, W.; Sun, F.; Zhang, C.; Zhang, L.; Sun, Z.; Qian, H. Platelet-rich plasma promotes MSCs exosomes paracrine to repair acute kidney injury via AKT/Rab27 pathway. Am. J. Transl. Res. 2021, 13, 1445. [Google Scholar] [PubMed]
- Ebrahim, N.; Ahmed, I.A.; Hussien, N.I.; Dessouky, A.A.; Farid, A.S.; Elshazly, A.M.; Mostafa, O.; El Gazzar, W.B.; Sorour, S.M.; Seleem, Y.; et al. Mesenchymal Stem Cell-Derived Exosomes Ameliorated Diabetic Nephropathy by Autophagy Induction through the mTOR Signaling Pathway. Cells 2018, 7, 226. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence (5′-3′) | Accession No. | Ref. |
|---|---|---|---|
| NGAL | F- TTGGGACAGGGAAGACGA R- TCACGCTGGGCAACATTA | XM_032901803.1 | [34] |
| TGFβ1 | F- CAGGAGCGCACAATCATGTT R- CTTTAGGAAGGACCTGGGTT | XM_032894155.1 | [35] |
| αSMA | F- GTCCCAGACATCAGGGAGTAA R- TCGGATACTTCAGCGTCAGGA | XM_032891814.1 | [35] |
| Caspase 3 | F- CTCGGTCTGGTACAGATGTCGATG R- GGTTAACCCGGGTAAGAATGTGCA | XM_032896303.1 | [36] |
| GAPDH | F- AGACAGCCGCATCTTCTTGT R- TTCCCATTCTCAGCCTTGAC | NM_017008.4 | [28] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alasmari, W.A.; Abdelfattah-Hassan, A.; El-Ghazali, H.M.; Abdo, S.A.; Ibrahim, D.; ElSawy, N.A.; El-Shetry, E.S.; Saleh, A.A.; Abourehab, M.A.S.; Mahfouz, H. Exosomes Derived from BM-MSCs Mitigate the Development of Chronic Kidney Damage Post-Menopause via Interfering with Fibrosis and Apoptosis. Biomolecules 2022, 12, 663. https://doi.org/10.3390/biom12050663
Alasmari WA, Abdelfattah-Hassan A, El-Ghazali HM, Abdo SA, Ibrahim D, ElSawy NA, El-Shetry ES, Saleh AA, Abourehab MAS, Mahfouz H. Exosomes Derived from BM-MSCs Mitigate the Development of Chronic Kidney Damage Post-Menopause via Interfering with Fibrosis and Apoptosis. Biomolecules. 2022; 12(5):663. https://doi.org/10.3390/biom12050663
Chicago/Turabian StyleAlasmari, Wardah A., Ahmed Abdelfattah-Hassan, Hanaa M. El-Ghazali, Samar A. Abdo, Doaa Ibrahim, Naser A. ElSawy, Eman S. El-Shetry, Ayman A. Saleh, Mohammed A. S. Abourehab, and Hala Mahfouz. 2022. "Exosomes Derived from BM-MSCs Mitigate the Development of Chronic Kidney Damage Post-Menopause via Interfering with Fibrosis and Apoptosis" Biomolecules 12, no. 5: 663. https://doi.org/10.3390/biom12050663
APA StyleAlasmari, W. A., Abdelfattah-Hassan, A., El-Ghazali, H. M., Abdo, S. A., Ibrahim, D., ElSawy, N. A., El-Shetry, E. S., Saleh, A. A., Abourehab, M. A. S., & Mahfouz, H. (2022). Exosomes Derived from BM-MSCs Mitigate the Development of Chronic Kidney Damage Post-Menopause via Interfering with Fibrosis and Apoptosis. Biomolecules, 12(5), 663. https://doi.org/10.3390/biom12050663