
Anti-Inflammatory Effects of GM1 Ganglioside on Endotoxin-Induced Uveitis in Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies
2.2. Animal Care
2.3. Endotoxin-Induced Uveitis (EIU) and Experimental Design
2.4. Evaluation of Clinical Manifestations
2.5. Determination of Cell Counts and Protein Concentration in the AqH
2.6. Histopathological Evaluation of Rats with EIU
2.7. Immunohistochemistry
2.8. RAW 264.7 Cell Culture
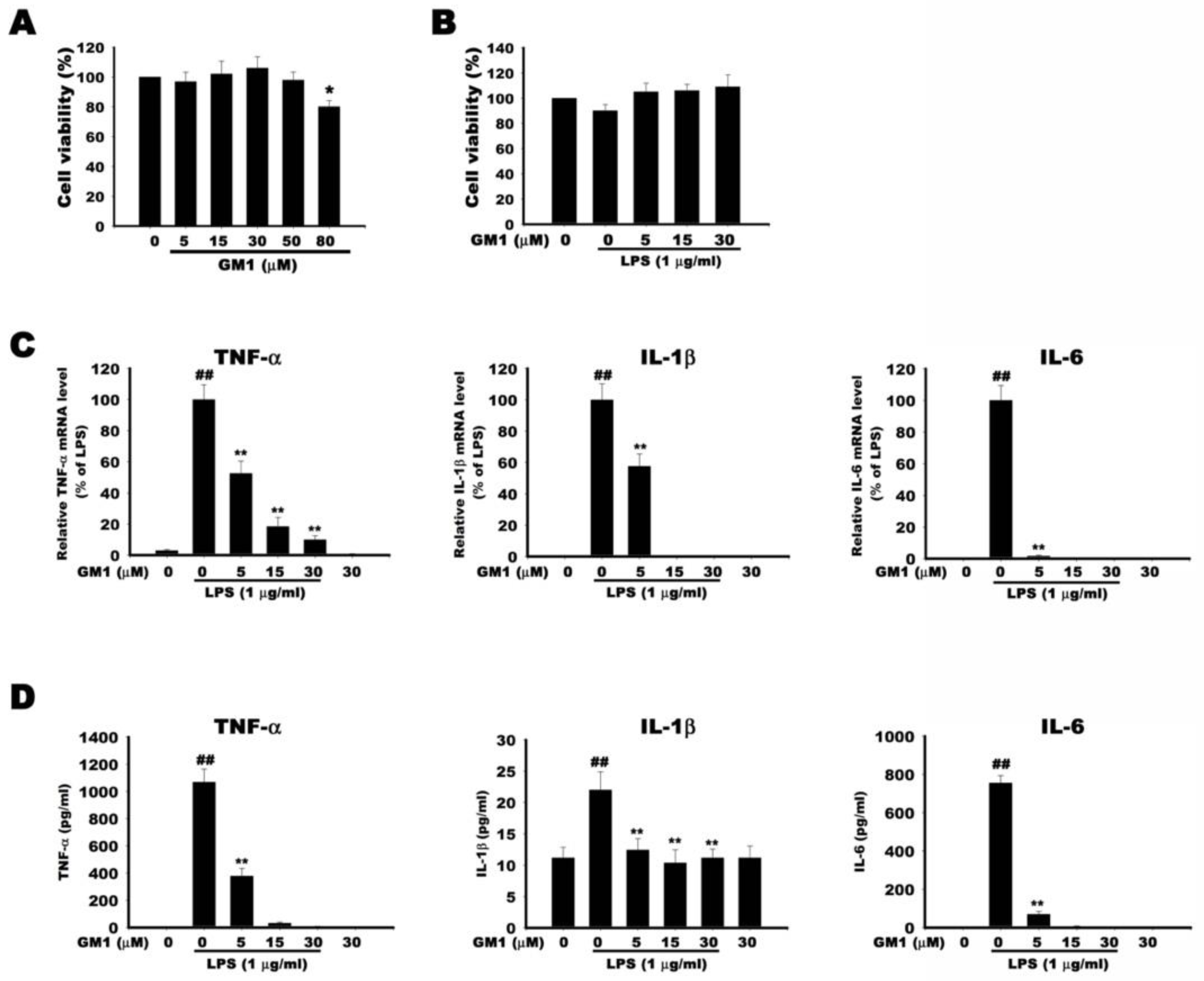
2.9. Cell Viability Assays
2.10. Enzyme-Linked Immunosorbent Assay (ELISA)
2.11. RT-qPCR Analysis
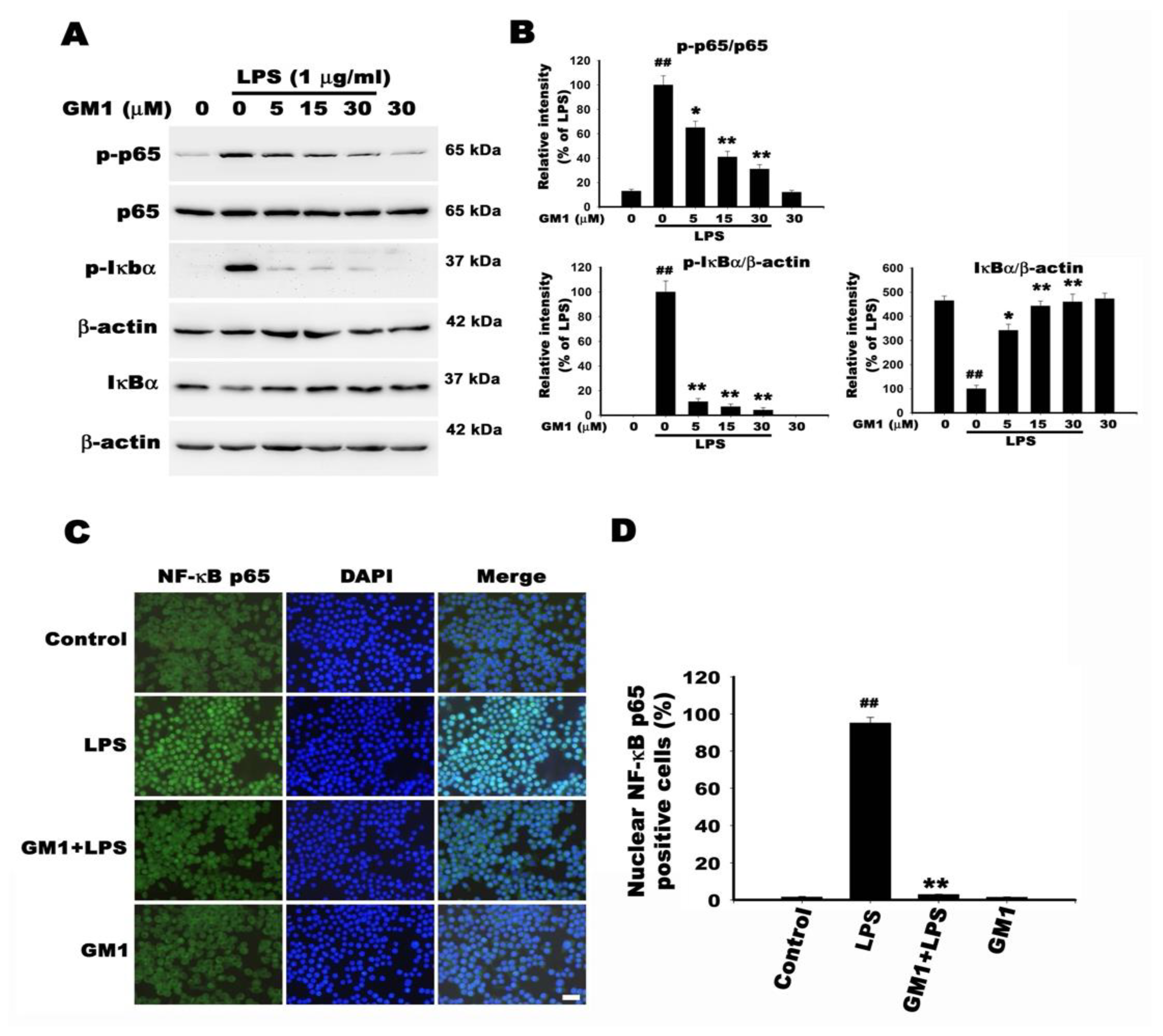
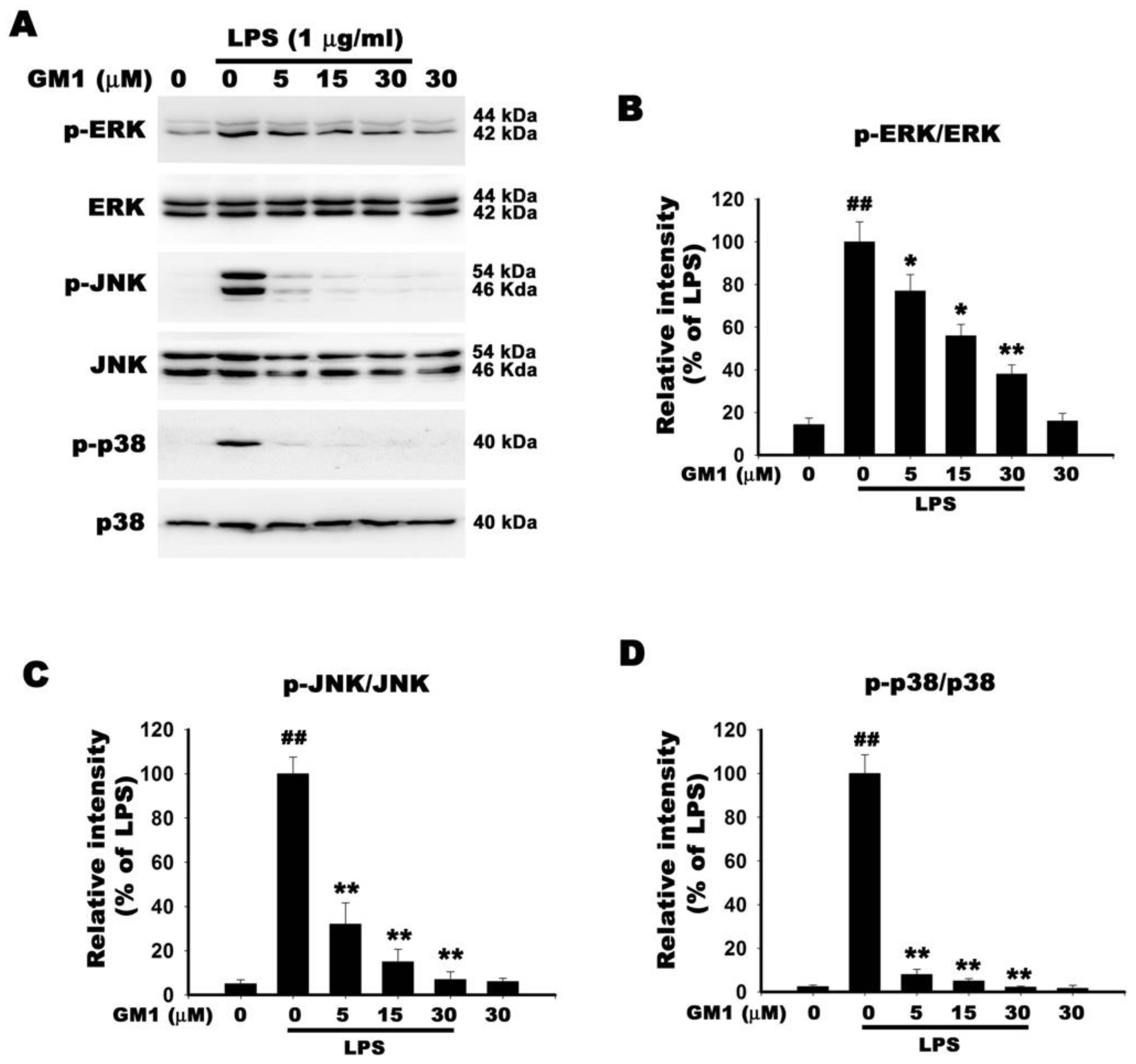
2.12. Western Blot
2.13. Immunofluorescence Staining
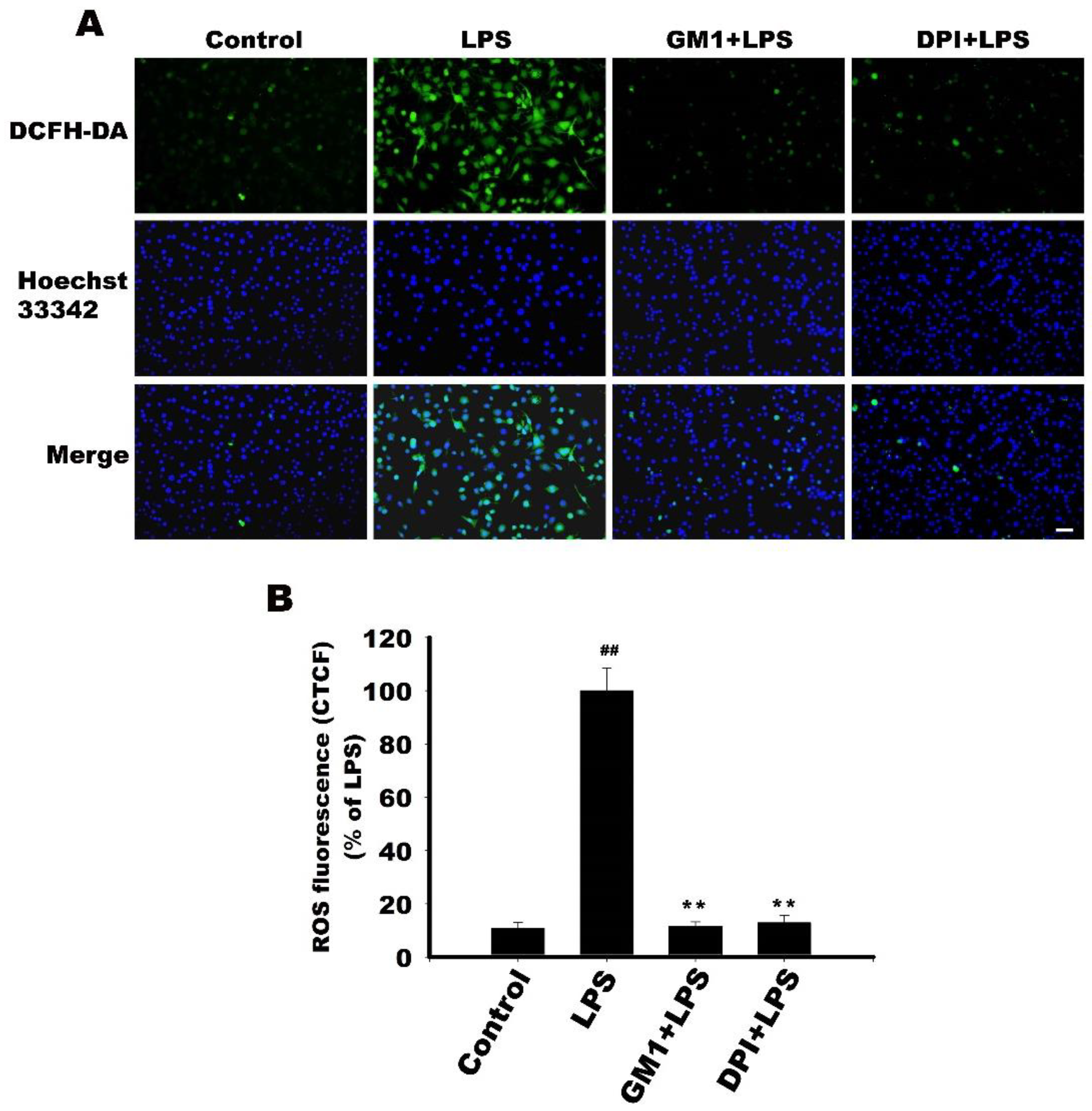
2.14. Detection of Reactive Oxygen Species (ROS)
2.15. Statistics
3. Results
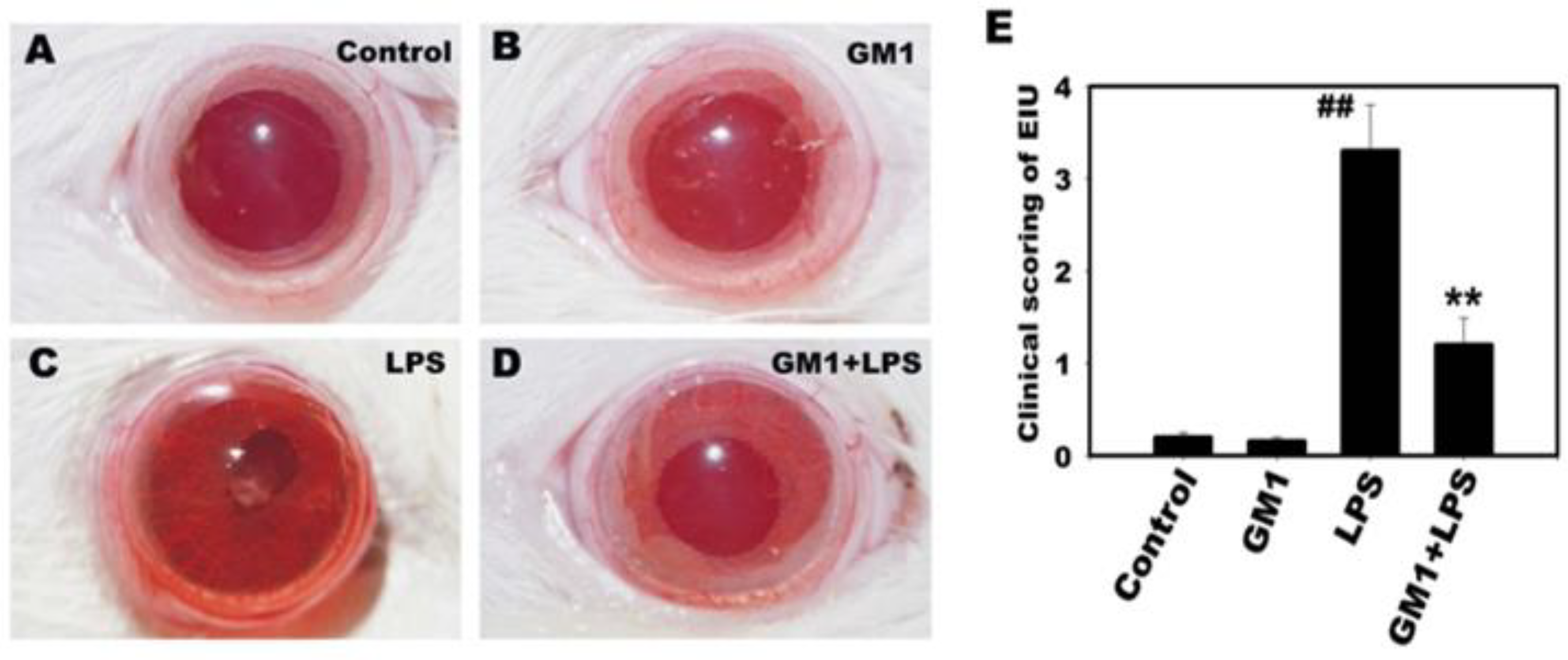
3.1. Effect of GM1 on the Inflammatory Manifestation of EIU in Rats
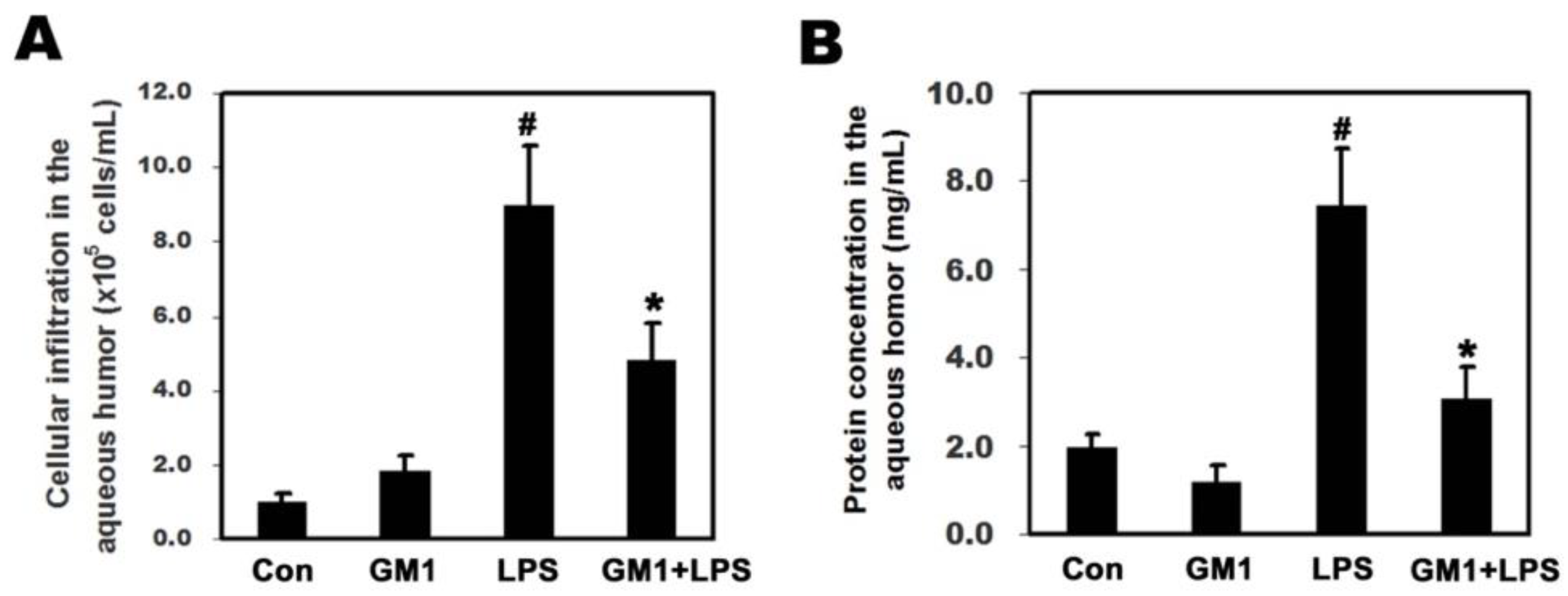
3.2. Effects of GM1 on LPS-Induced Cellular Infiltration and Protein Concentration in the AqH of Rats with EIU
3.3. Effects of GM1 on the Histopathological Changes Adjacent to the Iris-Ciliary Body (ICB) of EIU Rats
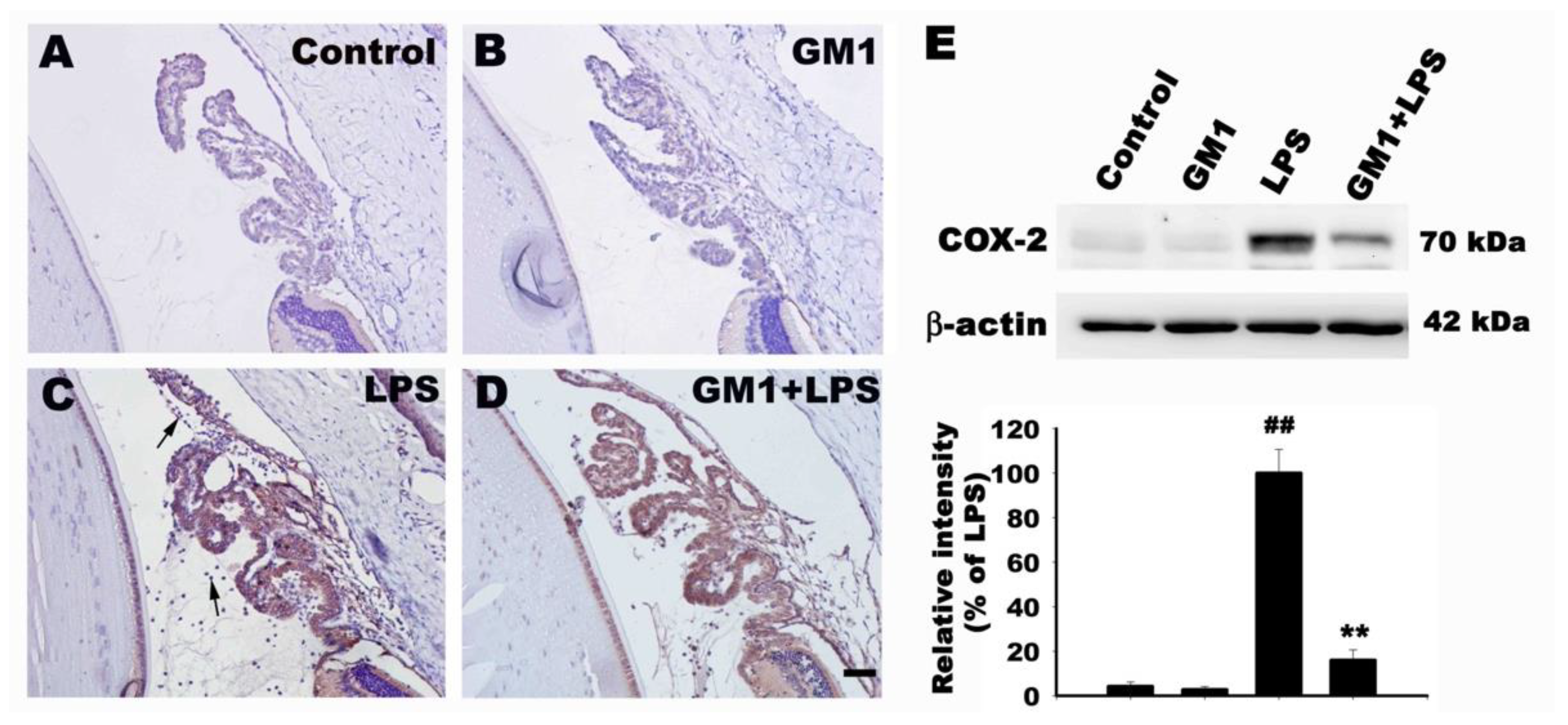
3.4. Effect of GM1 on the Recruitment of COX-2-Positive Cells Adjacent to the ICB in Rats with EIU
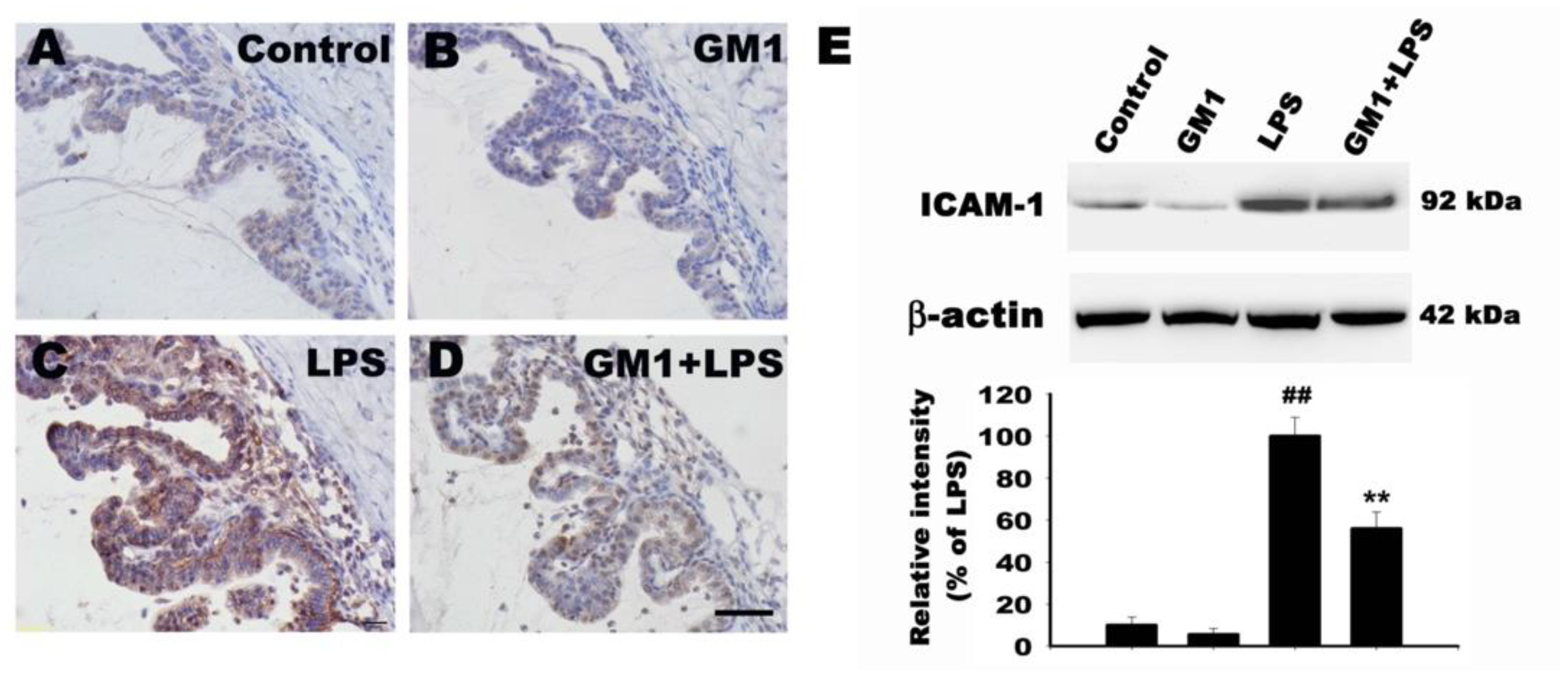
3.5. Effects of GM1 on ICAM-1 Expression in the ICB of Rats with EIU
3.6. Effect of GM1 on the Transcribed and Secreted Level of Proinflammatory Factors in LPS-Stimulated RAW 264.7 Cells
3.7. Effect of GM1 on the Protein Level of Proinflammatory Enzymes and Factors in LPS-Stimulated RAW 264.7 Cells
3.8. Effect of GM1 on NF-κB Activation in LPS-Stimulated RAW 264.7 Cells
3.9. Effect of GM1 on MAPK Activation in LPS-Stimulated RAW 264.7 Cells
3.10. Effect of GM1 on ROS Production by LPS-Stimulated RAW 264.7 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Durrani, O.M.; Tehrani, N.N.; Marr, J.E.; Moradi, P.; Stavrou, P.; Murray, P.I. Degree, duration, and causes of visual loss in uveitis. Br. J. Ophthalmol. 2004, 88, 1159–1162. [Google Scholar] [CrossRef] [Green Version]
- Harthan, J.; Fromstein, S.; Morettin, C.; Opitz, D. Diagnosis and treatment of anterior uveitis: Optometric management. Clin. Optom. 2016, 8, 23–35. [Google Scholar] [CrossRef] [Green Version]
- Krishna, U.; Ajanaku, D.; Denniston, A.; Gkika, T. Uveitis: A sight-threatening disease which can impact all systems. Postgrad. Med. J. 2017, 93, 766–773. [Google Scholar] [CrossRef]
- Impellizzeri, D.; Ahmad, A.; Bruschetta, G.; Di Paola, R.; Crupi, R.; Paterniti, I.; Esposito, E.; Cuzzocrea, S. The anti-inflammatory effects of palmitoylethanolamide (PEA) on endotoxin-induced uveitis in rats. Eur. J. Pharmacol. 2015, 761, 28–35. [Google Scholar] [CrossRef]
- Yadav, U.C.S.; Ramana, K.V. Endotoxin-Induced Uveitis in Rodents. In Mouse Models of Innate Immunity. Methods in Molecular Biology; Humana Press: New York, NY, USA, 2019; Volume 1960, pp. 161–168. [Google Scholar] [CrossRef]
- Lin, F.-L.; Ho, J.-D.; Cheng, Y.-W.; Chiou, G.C.Y.; Yen, J.-L.; Chang, H.-M.; Lee, T.-H.; Hsiao, G. Theissenolactone C Exhibited Ocular Protection of Endotoxin-Induced Uveitis by Attenuating Ocular Inflammatory Responses and Glial Activation. Front. Pharmacol. 2018, 9, 326. [Google Scholar] [CrossRef] [Green Version]
- Bhattacherjee, P.; Williams, R.N.; E Eakins, K. An evaluation of ocular inflammation following the injection of bacterial endotoxin into the rat foot pad. Investig. Ophthalmol. Vis. Sci. 1983, 24, 196–202. [Google Scholar]
- Hoekzema, R.; Verhagen, C.; Van Haren, M.; Kijlstra, A. Endotoxin-induced uveitis in the rat. The significance of intraocular interleukin-6. Investig. Ophthalmol. Vis. Sci. 1992, 33, 532–539. [Google Scholar]
- McMenamin, P.G.; Crewe, J. Endotoxin-induced uveitis. Kinetics and phenotype of the inflammatory cell infiltrate and the response of the resident tissue macrophages and dendritic cells in the iris and ciliary body. Investig. Ophthalmol. Vis. Sci. 1995, 36. [Google Scholar]
- Chen, W.; Hu, X.; Zhao, L.; Li, S.; Lu, H. Expression of toll-like receptor 4 in uvea-resident tissue macrophages during endotoxin-induced uveitis. Mol. Vis. 2009, 15, 619–628. [Google Scholar]
- Yuan, Z.; Chen, X.; Yang, W.; Lou, B.; Ye, N.; Liu, Y. The anti-inflammatory effect of minocycline on endotoxin-induced uveitis and retinal inflammation in rats. 2019, 25, 359–372. Mol. Vis. 2019, 25, 359–372. [Google Scholar]
- Cousins, S.W.; Guss, R.B.; Howes, E.L.; Rosenbaum, J.T. Endotoxin-induced uveitis in the rat: Observations on altered vascular permeability, clinical findings, and histology. Exp. Eye Res. 1984, 39, 665–676. [Google Scholar] [CrossRef]
- Park, J.; Kim, J.T.; Lee, S.J.; Kim, J.C. The anti-inflammatory effects of angiogenin in an endotoxin induced uveitis in rats. Int. J. Mol. Sci. 2022, 21, 413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, F.F.; Foster, C.S. Pharmacotherapy of uveitis. Expert Opin. Pharmacother. 2010, 11, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, E.; Papaliodis, G.N.; Lobo, A.-M.; Sobrin, L. Side-Effects of Anti-Inflammatory Therapy in Uveitis. Semin. Ophthalmol. 2014, 29, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Andreev, K. The Structural Role of Gangliosides: Insights from X-ray Scattering on Model Membranes. Curr. Med. Chem. 2020, 27, 6548–6570. [Google Scholar] [CrossRef] [PubMed]
- Sonnino, S.; Chiricozzi, E.; Grassi, S.; Mauri, L.; Prioni, S.; Prinetti, A. Gangliosides in Membrane Organization. Prog. Mol. Biol. Transl. Sci. 2018, 156, 83–120. [Google Scholar] [CrossRef]
- Xia, J.M.; Miao, Z.; Xie, C.L.; Zhang, J.W.; Yang, X.W. Chemical Constituents and Bioactivities of Starfishes: An Update. Chem. Biodivers. 2020, 17, e1900638. [Google Scholar] [CrossRef] [Green Version]
- Parton, R.G. Ultrastructural localization of gangliosides; GM1 is concentrated in caveolae. J. Histochem. Cytochem. 1994, 42, 155–166. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.K.; Nakatani, Y.; Yanagisawa, M. The role of glycosphingolipid metabolism in the developing brain. J. Lipid Res. 2009, 50, S440–S445. [Google Scholar] [CrossRef] [Green Version]
- Vyas, A.A.; Patel, H.V.; Fromholt, S.E.; Heffer-Lauc, M.; Vyas, K.A.; Dang, J.; Schachner, M.; Schnaar, R.L. Gangliosides are functional nerve cell ligands for myelin-associated glycoprotein (MAG), an inhibitor of nerve regeneration. Proc. Natl. Acad. Sci. USA 2002, 99, 8412–8417. [Google Scholar] [CrossRef] [Green Version]
- Ryckman, A.E.; Brockhausen, I.; Walia, J.S. Metabolism of Glycosphingolipids and Their Role in the Pathophysiology of Lysosomal Storage Disorders. Int. J. Mol. Sci. 2020, 21, 6881. [Google Scholar] [CrossRef] [PubMed]
- Sipione, S.; Monyror, J.; Galleguillos, D.; Steinberg, N.; Kadam, V. Gangliosides in the Brain: Physiology, Pathophysiology and Therapeutic Applications. Front. Neurosci. 2020, 14, 572965. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Ohmi, Y.; Tajima, O.; Ohkawa, Y.; Kondo, Y.; Shuting, J.; Hashimoto, N.; Furukawa, K. Gangliosides in Inflammation and Neurodegeneration. Prog. Mol. Biol. Transl. Sci. 2018, 156, 265–287. [Google Scholar] [CrossRef] [PubMed]
- Ohmi, Y.; Tajima, O.; Ohkawa, Y.; Yamauchi, Y.; Sugiura, Y.; Furukawa, K.; Furukawa, K. Gangliosides are essential in the protection of inflammation and neurodegeneration via maintenance of lipid rafts: Elucidation by a series of ganglioside-deficient mutant mice. J. Neurochem. 2011, 116, 926–935. [Google Scholar] [CrossRef]
- Chiricozzi, E.; Lunghi, G.; Di Biase, E.; Fazzari, M.; Sonnino, S.; Mauri, L. GM1 Ganglioside Is A Key Factor in Maintaining the Mammalian Neuronal Functions Avoiding Neurodegeneration. Int. J. Mol. Sci. 2020, 21, 868. [Google Scholar] [CrossRef] [Green Version]
- Nikolaeva, S.; Bayunova, L.; Sokolova, T.; Vlasova, Y.; Bachteeva, V.; Avrova, N.; Parnova, R. GM1 and GD1a gangliosides modulate toxic and inflammatory effects of E. coli lipopolysaccharide by preventing TLR4 translocation into lipid rafts. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2015, 1851, 239–247. [Google Scholar] [CrossRef]
- Vieira, K.P.; Zollner, A.R.D.A.E.S.L.; Malaguti, C.; Vilella, C.A.; Zollner, R.D.L. Ganglioside GM1 effects on the expression of nerve growth factor (NGF), Trk-A receptor, proinflammatory cytokines and on autoimmune diabetes onset in non-obese diabetic (NOD) mice. Cytokine 2008, 42, 92–104. [Google Scholar] [CrossRef]
- Cutrullis, R.A.; Poklépovich, T.J.; Postan, M.; Freilij, H.L.; Petray, P.B. Immunomodulatory and anti-fibrotic effects of ganglioside therapy on the cardiac chronic form of experimental Trypanosoma cruzi infection. Int. Immunopharmacol. 2011, 11, 1024–1031. [Google Scholar] [CrossRef]
- Gong, G.; Yin, L.; Yuan, L.; Sui, D.; Sun, Y.; Fu, H.; Chen, L.; Wang, X. Ganglioside GM1 protects against high altitude cerebral edema in rats by suppressing the oxidative stress and inflammatory response via the PI3K/AKT-Nrf2 pathway. Mol. Immunol. 2018, 95, 91–98. [Google Scholar] [CrossRef]
- Smith, J.; Hart, P.H.; Williams, K.A. Basic pathogenic mechanisms operating in experimental models of acute anterior uveitis. Immunol. Cell Biol. 1998, 76, 497–512. [Google Scholar] [CrossRef]
- Qin, Y.J.; Chu, K.O.; Yip, Y.W.Y.; Li, W.Y.; Yang, Y.P.; Chan, K.P.; Ren, J.L.; Chan, S.O.; Pang, C.P. Green Tea Extract Treatment Alleviates Ocular Inflammation in a Rat Model of Endotoxin-Induced Uveitis. PLoS ONE 2014, 9, e103995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Moreno, J.; Thillaye, B.; De Kozak, Y. Retino-Choroidal Changes in Endotoxin-Induced Uveitis in the Rat. Ophthalmic Res. 1992, 24, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Kanai, K.; Itoh, N.; Ito, Y.; Nagai, N.; Hori, Y.; Chikazawa, S.; Hoshi, F.; Higuchi, S. Anti-inflammatory potency of oral disulfiram compared with dexamethasone on endotoxin-induced uveitis in rats. J. Vet. Med. Sci. 2011, 73, 517–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fazzari, M.; Frasca, A.; Bifari, F.; Landsberger, N. Aminoglycoside drugs induce efficient read-through of CDKL5 nonsense mutations, slightly restoring its kinase activity. RNA Biol. 2019, 16, 1414–1423. [Google Scholar] [CrossRef]
- Kanagawa, T.; Matsuda, S.; Mikawa, Y.; Kogiso, M.; Nagasawa, H.; Himeno, K.; Hashimoto, Y.; Mimura, Y. Role of ICAM-1 and LFA-1 in endotoxin-induced uveitis in mice. Jpn. J. Ophthalmol. 1996, 40, 174–180. [Google Scholar]
- Liang, X.-M.; Guo, G.-F.; Huang, X.-H.; Duan, W.-L.; Zeng, Z.-L. Isotetrandrine protects against lipopolysaccharide-induced acute lung injury by suppression of mitogen-activated protein kinase and nuclear factor-kappa B. J. Surg. Res. 2014, 187, 596–604. [Google Scholar] [CrossRef]
- Bode, J.G.; Ehlting, C.; Häussinger, D. The macrophage response towards LPS and its control through the p38MAPK–STAT3 axis. Cell. Signal. 2012, 24, 1185–1194. [Google Scholar] [CrossRef]
- Ostareck, D.H.; Ostareck-Lederer, A. RNA-Binding Proteins in the Control of LPS-Induced Macrophage Response. Front. Genet. 2019, 10, 31. [Google Scholar] [CrossRef] [Green Version]
- Herb, M.; Schramm, M. Functions of ROS in Macrophages and Antimicrobial Immunity. Antioxidants 2021, 10, 313. [Google Scholar] [CrossRef]
- Bai, S.K.; Lee, S.J.; Na, H.J.; Ha, K.S.; Han, J.A.; Lee, H.; Kwon, Y.G.; Chung, C.K.; Kim, Y.M. beta-Carotene inhibits inflammatory gene expression in lipopolysaccharide-stimulated macrophages by suppressing redox-based NF-kappaB activation. Exp. Mol. Med. 2005, 37, 323–334. [Google Scholar]
- Bedard, K.; Krause, K.-H. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Cheng, C.; Zou, Y.; Peng, J. Oregano Essential Oil Attenuates RAW264.7 Cells from Lipopolysaccharide-Induced Inflammatory Response through Regulating NADPH Oxidase Activation-Driven Oxidative Stress. Molecules 2018, 23, 1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathan, C.; Cunningham-Bussel, A. Beyond oxidative stress: An immunologist’s guide to reactive oxygen species, Nature reviews. Immunology 2013, 13, 349–361. [Google Scholar] [PubMed] [Green Version]
- Kim, H.G.; Yoon, D.H.; Lee, W.H.; Han, S.K.; Shrestha, B.; Kim, C.H.; Lim, M.H.; Chang, W.; Lim, S.; Choi, S.; et al. Phellinus linteus inhibits inflammatory mediators by suppressing redox-based NF-kappaB and MAPKs activation in lipopolysaccharide-induced RAW 264.7 macrophage. J. Ethnopharmacol. 2007, 114, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Streilein, J.W.; Ohta, K.; Mo, J.S.; Taylor, A.W. Ocular Immune Privilege and the Impact of Intraocular Inflammation. DNA Cell Biol. 2002, 21, 453–459. [Google Scholar] [CrossRef]
- Chen, C.-L.; Chen, J.-T.; Liang, C.-M.; Tai, M.-C.; Lu, D.-W.; Chen, Y.-H. Silibinin treatment prevents endotoxin-induced uveitis in rats in vivo and in vitro. PLoS ONE 2017, 12, e0174971. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.-H.; Horng, C.-T.; Chen, Y.-H.; Chen, P.-L.; Chen, C.-L.; Liang, C.-M.; Chien, M.-W.; Chen, J.-T. Inhibitory Effects of Glucosamine on Endotoxin-Induced Uveitis in Lewis Rats. Investig. Ophthalmol. Vis. Sci. 2008, 49, 5441–5449. [Google Scholar] [CrossRef]
- Mérida, S.; Palacios, E.; Navea, A.; Morell, F.B. Macrophages and Uveitis in Experimental Animal Models. Mediat. Inflamm. 2015, 2015, 671417. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Lu, H.; Hu, X.; Chen, W.; Xu, Y.; Wang, J. Expression of TLR4-MyD88 and NF-kappaB in the iris during endotoxin-induced uveitis. Mediat. Inflamm. 2010, 2010, 748218. [Google Scholar] [CrossRef] [Green Version]
- Uchida, T.; Honjo, M.; Yamagishi, R.; Aihara, M. The Anti-Inflammatory Effect of Ripasudil (K-115), a Rho Kinase (ROCK) Inhibitor, on Endotoxin-Induced Uveitis in Rats. Investig. Opthalmol. Vis. Sci. 2017, 58, 5584–5593. [Google Scholar] [CrossRef] [Green Version]
- Guha, M.; Mackman, N. LPS induction of gene expression in human monocytes. Cell. Signal. 2001, 13, 85–94. [Google Scholar] [CrossRef]
- Brown, J.; Wang, H.; Hajishengallis, G.N.; Martin, M. TLR-signaling networks: An integration of adaptor molecules, kinases, and cross-talk. J. Dent. Res. 2011, 90, 417–427. [Google Scholar] [CrossRef]
- Denkers, E.Y.; Butcher, B.A.; Del Rio, L.; Kim, L. Manipulation of mitogen-activated protein kinase/nuclear factor-kappaB-signaling cascades during intracellular Toxoplasma gondii infection. Immunol. Rev. 2004, 201, 191–205. [Google Scholar] [CrossRef]
- Cargnello, M.; Roux, P.P. Activation and Function of the MAPKs and Their Substrates, the MAPK-Activated Protein Kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83, Correction in Microbiol. Mol. Biol. Rev. 2012, 76, 496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthur, J.S.C.; Ley, S.C. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 2013, 13, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Osthoff, K.; Ferrari, D.; Riehemann, K.; Wesselborg, S. Regulation of NF-kappa B activation by MAP kinase cascades. Immunobiology 1997, 198, 35–49. [Google Scholar] [CrossRef]
- Canton, M.; Sánchez-Rodríguez, R.; Spera, I.; Venegas, F.C.; Favia, M.; Viola, A.; Castegna, A. Reactive Oxygen Species in Macrophages: Sources and Targets. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Park, J.; Kwak, C.H.; Ha, S.H.; Kwon, K.M.; Abekura, F.; Cho, S.H.; Chang, Y.C.; Lee, Y.C.; Ha, K.T.; Chung, T.W.; et al. Ganglioside GM3 suppresses lipopolysaccharide-induced inflammatory responses in rAW 264.7 macrophage cells through NF-kappaB, AP-1, and MAPKs signaling. J. Cell. Biochem. 2018, 119, 1173–1182. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, Y.; Cao, F.; Qin, Y.; Li, W.; Zhang, J. Ganglioside GD1a suppresses LPS-induced pro-inflammatory cytokines in RAW264.7 macrophages by reducing MAPKs and NF-kappaB signaling pathways through TLR4. Int. Immunopharmacol. 2015, 28, 136–145. [Google Scholar] [CrossRef]
- Walters, K.M.; Woessner, K.M. An Overview of Nonsteroidal Antiinflammatory Drug Reactions. Immunol. Allergy Clin. N. Am. 2016, 36, 625–641. [Google Scholar] [CrossRef]
- Pleyer, U.; Ursell, P.G.; Rama, P. Intraocular Pressure Effects of Common Topical Steroids for Post-Cataract Inflammation: Are They All the Same? Ophthalmol. Ther. 2013, 2, 55–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhar, S.; Seth, J.; Parikh, D. Systemic side-effects of topical corticosteroids. Indian J. Dermatol. 2014, 59, 460–464. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wang, Y.; Jiang, C.; Cao, H.; Jiang, J.; Xu, B.; Sun, T. Ganglioside Monosialic Acid Alleviates Peripheral Neuropathy Induced by Utidelone Plus Capecitabine in Metastatic Breast Cancer From a Phase III Clinical Trial. Front. Oncol. 2020, 10, 524223. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weng, T.-H.; Ke, C.-C.; Huang, Y.-S. Anti-Inflammatory Effects of GM1 Ganglioside on Endotoxin-Induced Uveitis in Rats. Biomolecules 2022, 12, 727. https://doi.org/10.3390/biom12050727
Weng T-H, Ke C-C, Huang Y-S. Anti-Inflammatory Effects of GM1 Ganglioside on Endotoxin-Induced Uveitis in Rats. Biomolecules. 2022; 12(5):727. https://doi.org/10.3390/biom12050727
Chicago/Turabian StyleWeng, Tzu-Heng, Chang-Chih Ke, and Yuahn-Sieh Huang. 2022. "Anti-Inflammatory Effects of GM1 Ganglioside on Endotoxin-Induced Uveitis in Rats" Biomolecules 12, no. 5: 727. https://doi.org/10.3390/biom12050727
APA StyleWeng, T.-H., Ke, C.-C., & Huang, Y.-S. (2022). Anti-Inflammatory Effects of GM1 Ganglioside on Endotoxin-Induced Uveitis in Rats. Biomolecules, 12(5), 727. https://doi.org/10.3390/biom12050727