
Slug Mediates MRP2 Expression in Non-Small Cell Lung Cancer Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Transfection of Slug and Mock Plasmids
2.4. mRNA Extraction and cDNA Synthesis
2.5. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.6. Electrophoresis and Western Blotting
2.7. CDCF Uptake Assay
2.8. SN-38 Uptake Assay
2.9. Cell Viability Assay
2.10. Statistical Analysis
3. Results
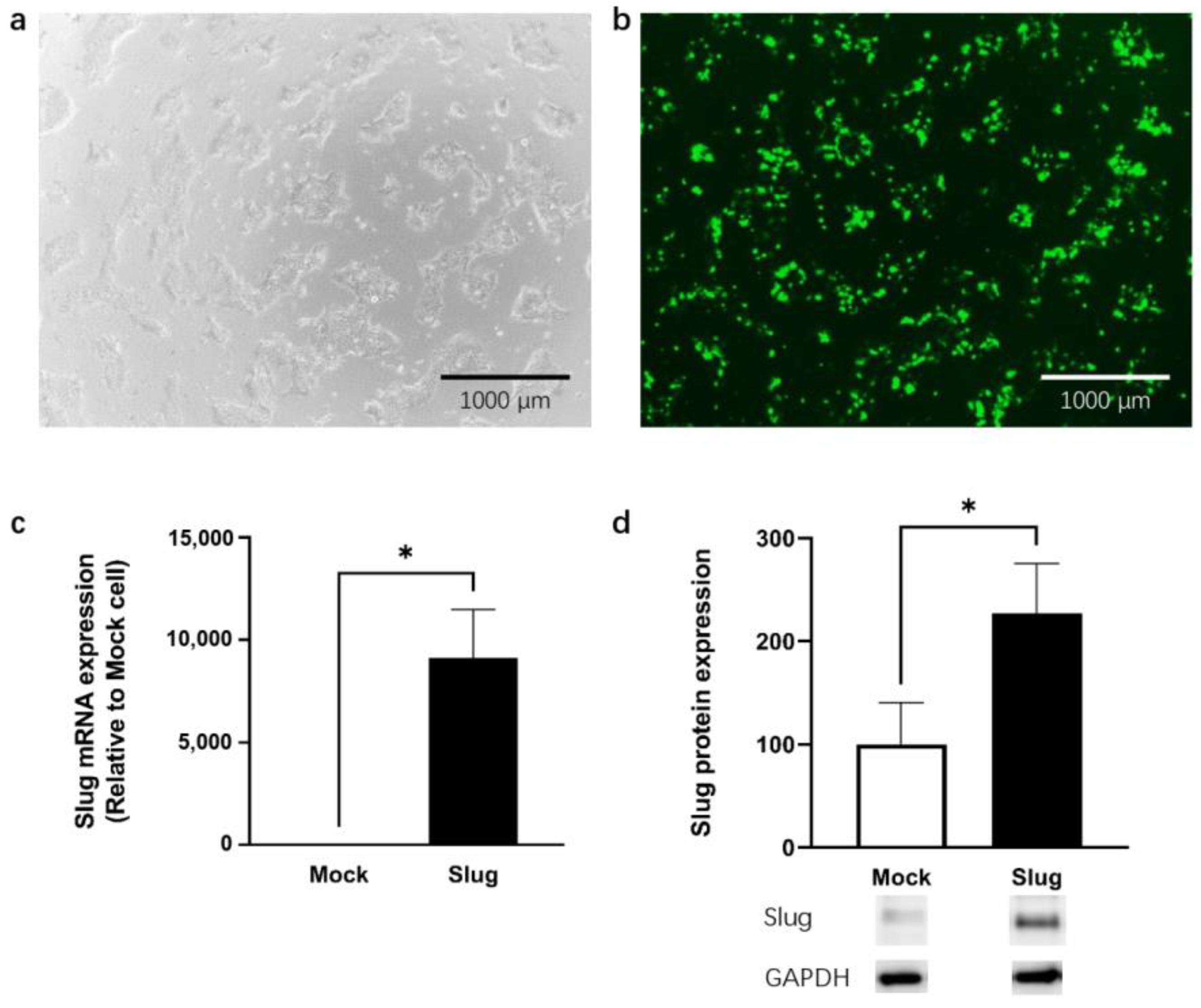
3.1. mRNA Expression Levels of Slug in Cells
3.2. mRNA Expression Levels of Efflux Transporters and ERM Proteins
3.3. Protein Expression Level of MRP2 in Slug-Overexpressing HCC827 Cells
3.4. Uptake of CDCF and SN-38 in Slug-Transfected Cells
3.5. Cell Viability in Slug-Transfected Cells after SN-38 Administration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Blandin, K.S.; Crosbie, P.A.; Balata, H.; Chudziak, J.; Hussell, T.; Dive, C. Progress and prospects of early detection in lung cancer. Open Biol. 2017, 7, 170070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.H.; Liu, Y.J.; De, J.M. Risk Factors, Prognosis, and a New Nomogram for Predicting Cancer-Specific Survival Among Lung Cancer Patients with Brain Metastasis: A Retrospective Study Based on SEER. Lung 2022, 200, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Rocco, D.; Della, G.L.; Battiloro, C.; Gridelli, C. The role of nivolumab combined to immunotherapy and/or chemotherapy in the first-line treatment of advanced Non Small Cell Lung Cancer. Expert Opin. Biol. Ther. 2021, 21, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Sheng, H.; Wan, D.; Zhu, C.; Jiang, R.; Sun, X.; Feng, J. Prognostic role of multidrug resistance-associated protein 1 expression and platelet count in operable non-small cell lung cancer. Oncol. Lett. 2018, 16, 1123–1132. [Google Scholar] [CrossRef] [Green Version]
- Leslie, E.M.; Deeley, R.G.; Cole, S.P. Multidrug resistance proteins: Role of P-glycoprotein, MRP1, MRP2, and BCRP (ABCG2) in tissue defense. Toxicol. Appl. Pharmacol. 2005, 204, 216–237. [Google Scholar] [CrossRef]
- Ogihara, T.; Mizoi, K.; Kamioka, H.; Yano, K. Physiological Roles of ERM Proteins and Transcriptional Regulators in Supporting Membrane Expression of Efflux Transporters as Factors of Drug Resistance in Cancer. Cancers 2020, 12, 3352. [Google Scholar] [CrossRef]
- Mottino, A.D.; Catania, V.A. Hepatic drug transporters and nuclear receptors: Regulation by therapeutic agents. World J. Gastroenterol. 2008, 14, 7068–7074. [Google Scholar] [CrossRef]
- Fan, Y.; Mansoor, N.; Ahmad, T.; Wu, Z.X.; Khan, R.A.; Czejka, M.; Sharib, S.; Ahmed, M.; Chen, Z.S.; Yang, D.H. Enzyme and Transporter Kinetics for CPT-11 (Irinotecan) and SN-38: An Insight on Tumor Tissue Compartment Pharmacokinetics Using PBPK. Recent Pat. Anticancer Drug Discov. 2019, 14, 177–186. [Google Scholar] [CrossRef]
- Materna, V.; Holm, P.S.; Dietel, M.; Lage, H. Kinetic characterization of ribozymes directed against the cisplatin resistance-associated ABC transporter cMOAT/MRP2/ABCC2. Cancer Gene Ther. 2001, 8, 176–184. [Google Scholar] [CrossRef] [Green Version]
- Cho, E.S.; Kang, H.E.; Kim, N.H.; Yook, J.I. Therapeutic implications of cancer epithelial-mesenchymal transition (EMT). Arch. Pharm. Res. 2019, 42, 14–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Carstens, J.L.; Kim, J.; Scheible, M.; Kaye, J.; Sugimoto, H.; Wu, C.C.; LeBleu, V.S.; Kalluri, R. Epithelial-to-mesenchymal transition is dispensable for metastasis but induces chemoresistance in pancreatic cancer. Nature 2015, 527, 525–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gras, B.; Jacqueroud, L.; Wierinckx, A.; Lamblot, C.; Fauvet, F.; Lachuer, J.; Puisieux, A.; Ansieau, S. Snail family members unequally trigger EMT and thereby differ in their ability to promote the neoplastic transformation of mammary epithelial cells. PLoS ONE 2014, 9, e92254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamioka, H.; Tomono, T.; Fujita, A.; Onozato, R.; Iijima, M.; Tsuchida, S.; Arai, T.; Fujita, Y.; Zhang, X.; Yano, K.; et al. Moesin-Mediated P-Glycoprotein Activation during Snail-Induced Epithelial-Mesenchymal Transition in Lung Cancer Cells. J. Pharm. Sci. 2020, 109, 2302–2308. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, W.; Edaki, K.; Nakazawa, Y.; Kamioka, H.; Fujita, A.; Onozato, R.; Iijima, M.; Tsuchida, S.; Arai, T.; et al. Correlations of mRNA Levels among Efflux Transporters, Transcriptional Regulators, and Scaffold Proteins in Non-Small-Cell Lung Cancer. Can. J. Infect. Dis. Med. Microbiol. 2021, 2021, 4005327. [Google Scholar] [CrossRef]
- Yano, K.; Todokoro, I.; Kamioka, H.; Tomono, T.; Ogihara, T. Functional Alterations of Multidrug Resistance-Associated Proteins 2 and 5, and Breast Cancer Resistance Protein upon Snail-Induced Epithelial-Mesenchymal Transition in HCC827 Cells. Biol. Pharm. Bull. 2021, 44, 103–111. [Google Scholar] [CrossRef]
- Yano, K.; Tomono, T.; Sakai, R.; Kano, T.; Morimoto, K.; Kato, Y.; Ogihara, T. Contribution of radixin to P-glycoprotein expression and transport activity in mouse small intestine in vivo. J. Pharm. Sci. 2013, 102, 2875–2881. [Google Scholar] [CrossRef]
- Kamioka, H.; Edaki, K.; Kasahara, H.; Tomono, T.; Yano, K.; Ogihara, T. Drug resistance via radixin-mediated increase of P-glycoprotein membrane expression during SNAI1-induced epithelial-mesenchymal transition in HepG2 cells. J. Pharm. Pharmacol. 2021, 73, 1609–1616. [Google Scholar] [CrossRef]
- Saeki, J.; Sekine, S.; Horie, T. LPS-induced dissociation of multidrug resistance-associated protein 2 (Mrp2) and radixin is associated with Mrp2 selective internalization in rats. Biochem. Pharmacol. 2011, 81, 178–184. [Google Scholar] [CrossRef]
- Yang, Q.; Onuki, R.; Nakai, C.; Sugiyama, Y. Ezrin and radixin both regulate the apical membrane localization of ABCC2 (MRP2) in human intestinal epithelial Caco-2 cells. Exp. Cell Res. 2007, 313, 3517–3525. [Google Scholar] [CrossRef]
- Kawase, A.; Inoue, Y.; Nakazaki, S.; Koizumi, E.; Iwaki, M. Radixin knockdown improves the accumulation and efficiency of methotrexate in tumor cells. Oncol. Rep. 2019, 42, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Yano, K.; Otsuka, K.; Kato, Y.; Kawabata, H.; Ohmori, S.; Arakawa, H.; Ogihara, T. Different regulation of P-glycoprotein function between Caco-2 and Caki-1 cells by ezrin, radixin and moesin proteins. J. Pharm. Pharmacol. 2016, 68, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, I.; Nishikawa, S.; Yamamoto, A.; Kono, Y.; Fujita, T. The Impact of Breast Cancer Resistance Protein (BCRP/ABCG2) on Drug Transport Across Caco-2 Cell Monolayers. Drug Metab. Dispos. 2020, 48, 491–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takara, K.; Kitada, N.; Yoshikawa, E.; Yamamoto, K.; Horibe, S.; Sakaeda, T.; Nishiguchi, K.; Ohnishi, N.; Yokoyama, T. Molecular changes to HeLa cells on continuous exposure to SN-38, an active metabolite of irinotecan hydrochloride. Cancer Lett. 2009, 278, 88–96. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Liu, W.; Edaki, K.; Nakazawa, Y.; Takahashi, S.; Sunakawa, H.; Mizoi, K.; Ogihara, T. Slug Mediates MRP2 Expression in Non-Small Cell Lung Cancer Cells. Biomolecules 2022, 12, 806. https://doi.org/10.3390/biom12060806
Zhang X, Liu W, Edaki K, Nakazawa Y, Takahashi S, Sunakawa H, Mizoi K, Ogihara T. Slug Mediates MRP2 Expression in Non-Small Cell Lung Cancer Cells. Biomolecules. 2022; 12(6):806. https://doi.org/10.3390/biom12060806
Chicago/Turabian StyleZhang, Xieyi, Wangyang Liu, Kazue Edaki, Yuta Nakazawa, Saori Takahashi, Hiroki Sunakawa, Kenta Mizoi, and Takuo Ogihara. 2022. "Slug Mediates MRP2 Expression in Non-Small Cell Lung Cancer Cells" Biomolecules 12, no. 6: 806. https://doi.org/10.3390/biom12060806
APA StyleZhang, X., Liu, W., Edaki, K., Nakazawa, Y., Takahashi, S., Sunakawa, H., Mizoi, K., & Ogihara, T. (2022). Slug Mediates MRP2 Expression in Non-Small Cell Lung Cancer Cells. Biomolecules, 12(6), 806. https://doi.org/10.3390/biom12060806