
Impact of Ca2+-Induced PI(4,5)P2 Clusters on PH-YFP Organization and Protein-Protein Interactions

 , ,
, ,  , , ,
, , ,  and
and
Abstract
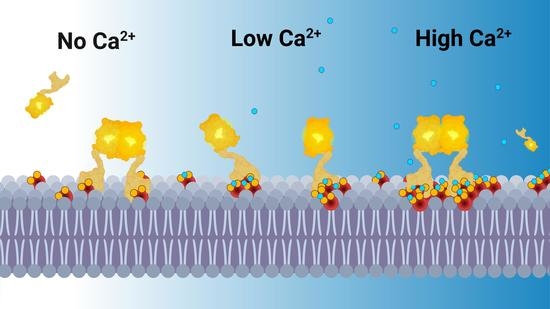
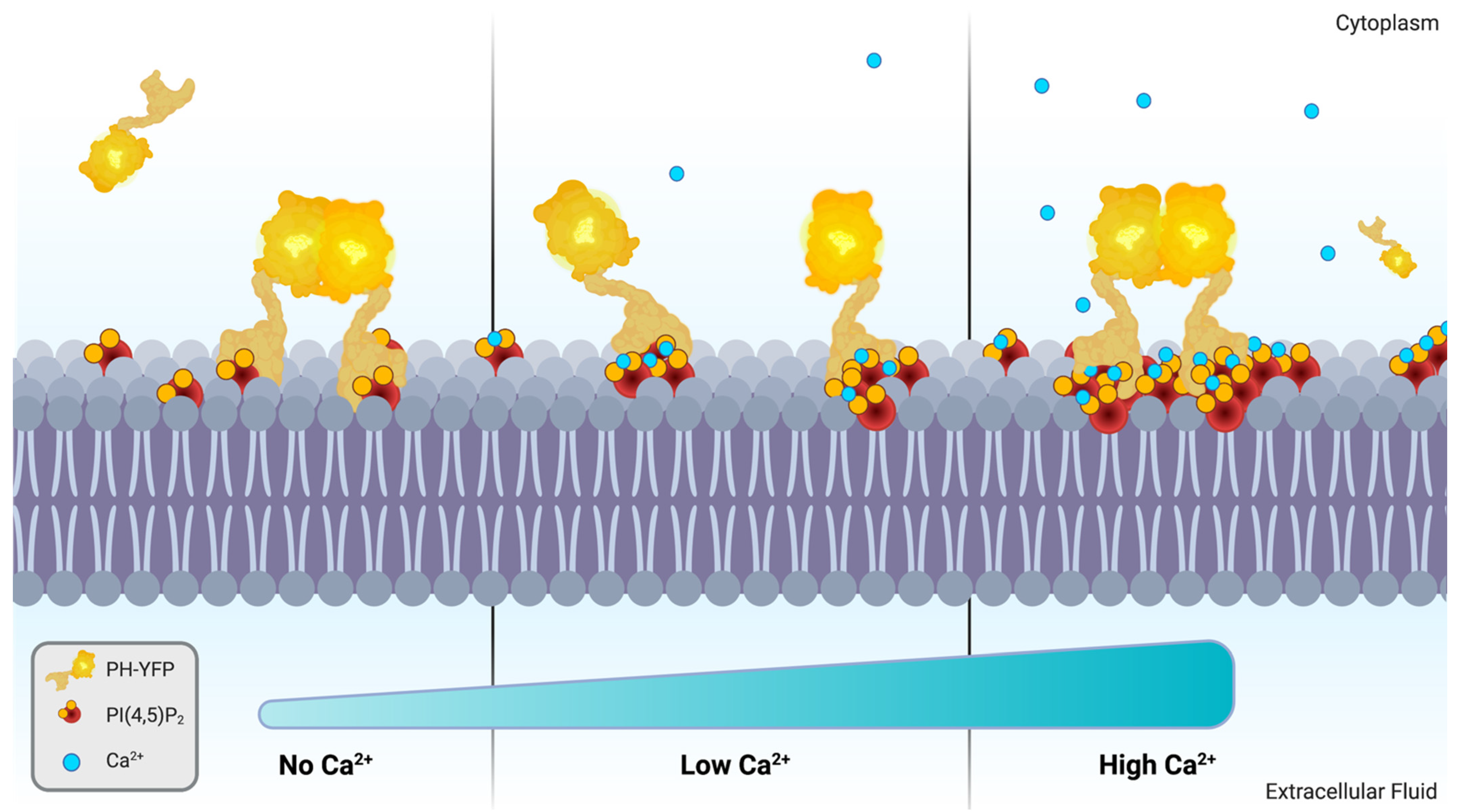
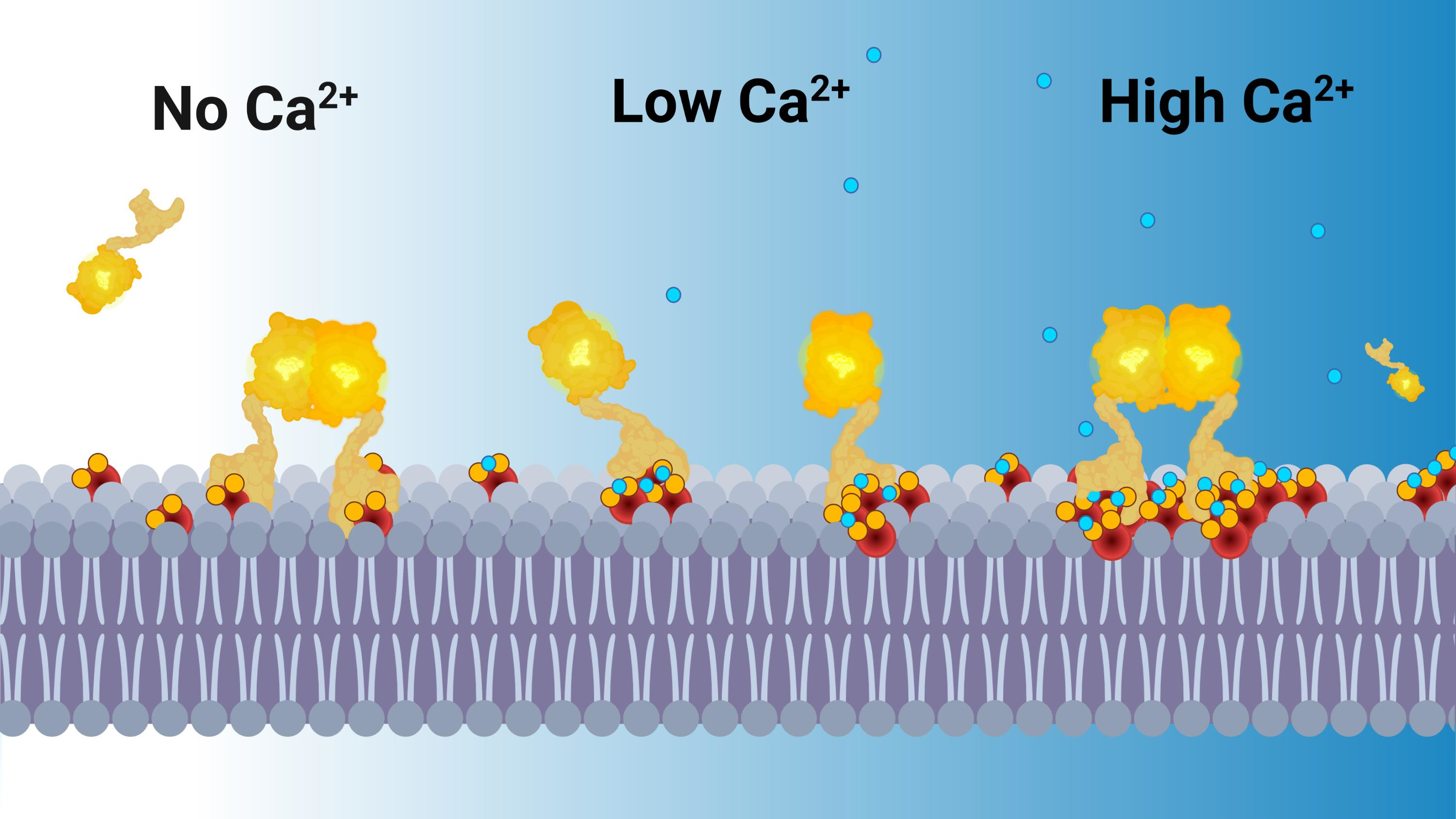
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. PH-YFP Expression and Purification
2.3. Liposome Preparation
2.4. Steady-State Fluorescence Spectroscopy
Photobleaching Assay
2.5. Confocal Fluorescence Microscopy
2.6. Fluorescence Fluctuation Spectroscopy
2.7. Statistical Analysis
3. Results
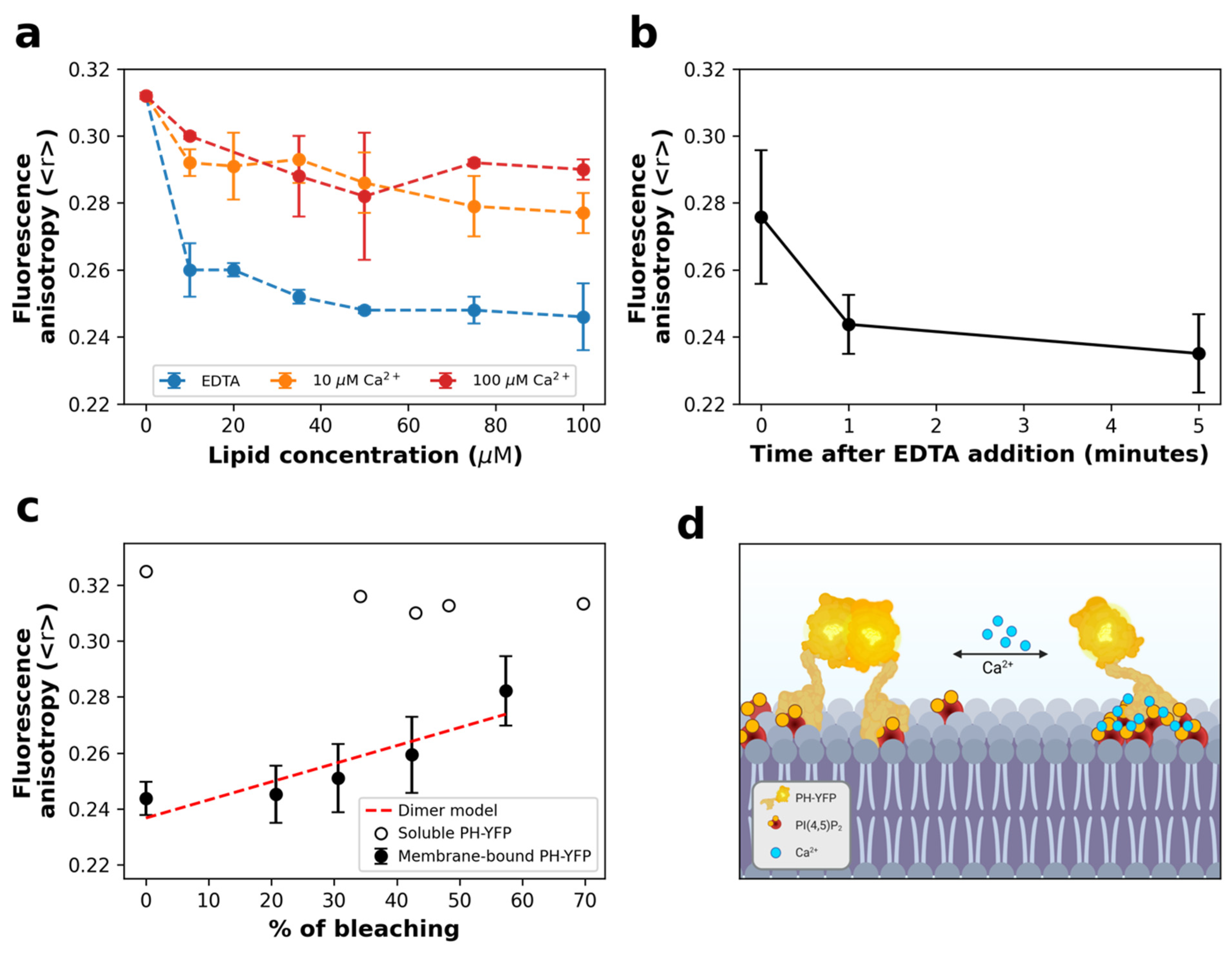
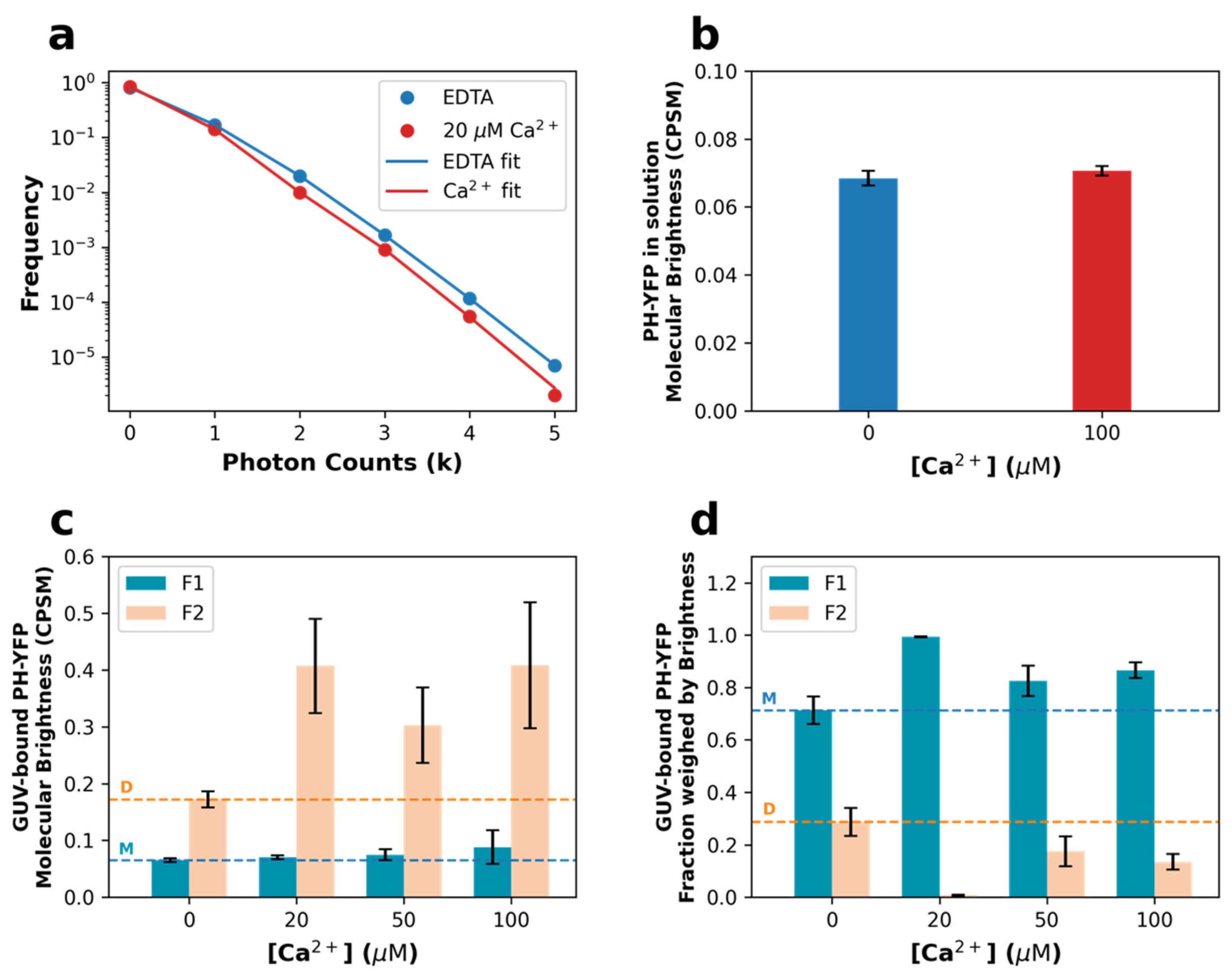
3.1. Homo-FRET Assays
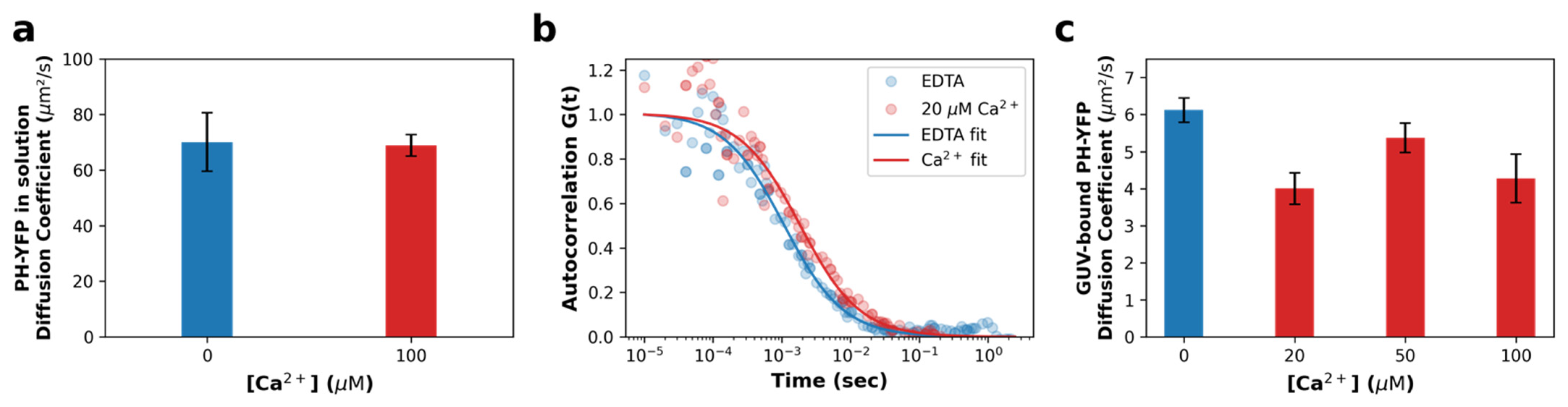
3.2. Fluorescence Fluctuation Spectroscopy (FFS) Assays
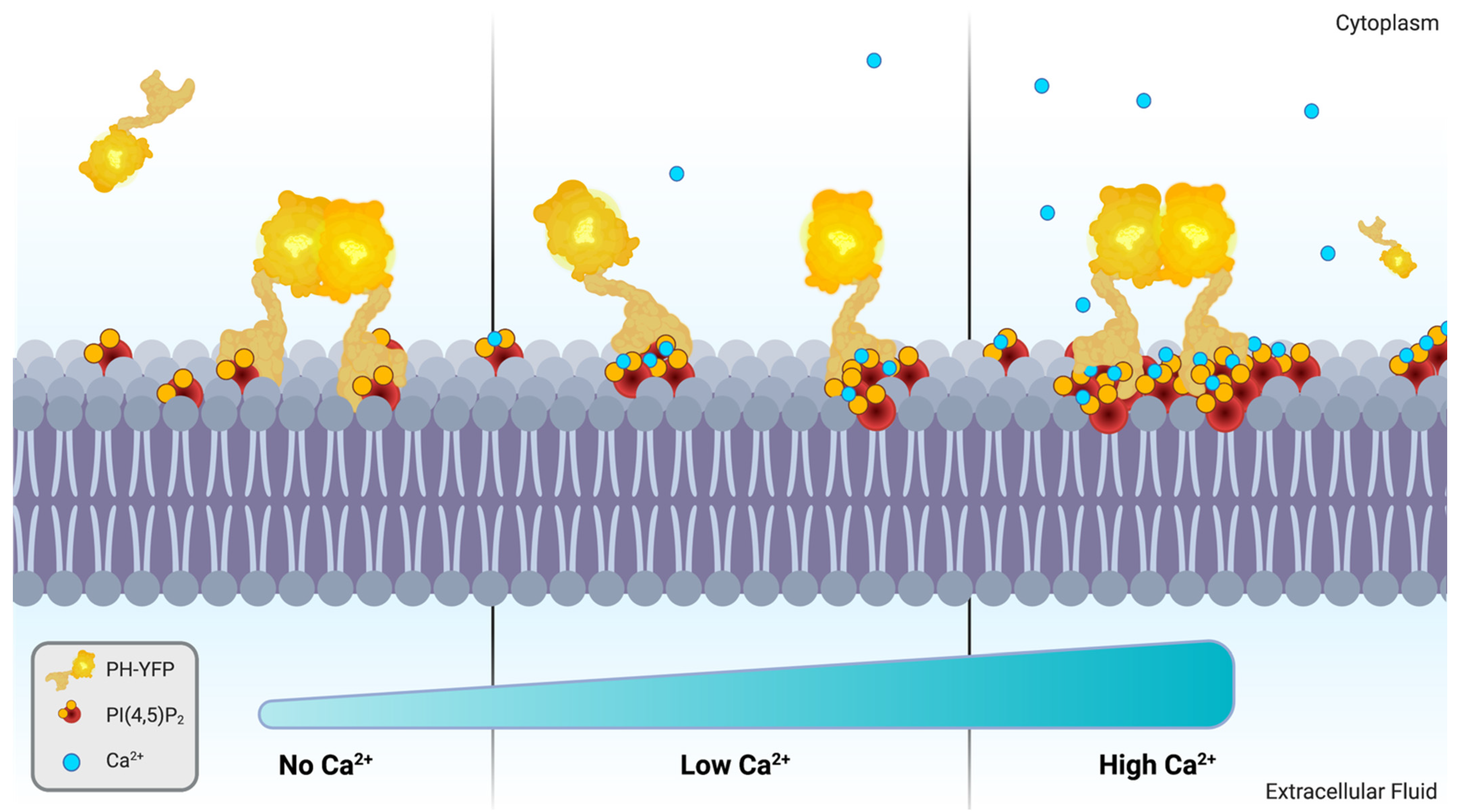
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Whited, A.M.; Johs, A. The Interactions of Peripheral Membrane Proteins with Biological Membranes. Chem. Phys. Lipids 2015, 192, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Kwiatkowska, K. One Lipid, Multiple Functions: How Various Pools of PI (4, 5) P 2 Are Created in the Plasma Membrane. Cell. Mol. Life Sci. 2010, 67, 3927–3946. [Google Scholar] [CrossRef]
- Ferrell, J.E.; Huestis, W.H. Phosphoinositide Metabolism and the Morphology of Human Erythrocytes. J. Cell Biol. 1984, 98, 1992–1998. [Google Scholar] [CrossRef]
- Balla, T. Phosphoinositides: Tiny Lipids with Giant Impact on Cell Regulation. Physiol. Rev. 2013, 93, 1019–1137. [Google Scholar] [CrossRef] [PubMed]
- Sarmento, M.J.; Borges-Araújo, L.; Pinto, S.N.; Bernardes, N.; Ricardo, J.C.; Coutinho, A.; Prieto, M.; Fernandes, F. Quantitative FRET Microscopy Reveals a Crucial Role of Cytoskeleton in Promoting PI(4,5)P2 Confinement. Int. J. Mol. Sci. 2021, 22, 11727. [Google Scholar] [CrossRef]
- Golebiewska, U.; Nyako, M.; Woturski, W.; Zaitseva, I.; Mclaughlin, S. Diffusion Coefficient of Fluorescent Phosphatidylinositol 4, 5-Bisphosphate in the Plasma Membrane of Cells. Mol. Biol. Cell 2008, 19, 1663–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Bogaart, G.; Meyenberg, K.; Risselada, H.J.; Amin, H.; Willig, K.I.; Hubrich, B.E.; Dier, M.; Hell, S.W.; Grubmüller, H.; Diederichsen, U.; et al. Membrane Protein Sequestering by Ionic Protein–Lipid Interactions. Nature 2011, 479, 552–555. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, S.; Wang, J.; Gambhir, A.; Murray, D. PIP(2) and Proteins: Interactions, Organization, and Information Flow. Annu. Rev. Biophys. Biomol. Struct. 2002, 31, 151–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges-Araújo, L.; Fernandes, F. Structure and Lateral Organization of Phosphatidylinositol 4,5-Bisphosphate. Molecules 2020, 25, 3885. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, M.E.; Sarmento, M.J.; Fernandes, F. Role of Calcium in Membrane Interactions by PI(4,5)P2-Binding Proteins. Biochem. Soc. Trans. 2014, 42, 1441–1446. [Google Scholar] [CrossRef] [PubMed]
- Di Paolo, G.; De Camilli, P. Phosphoinositides in Cell Regulation and Membrane Dynamics. Nature 2006, 443, 651–657. [Google Scholar] [CrossRef]
- Höning, S.; Ricotta, D.; Krauss, M.; Späte, K.; Spolaore, B.; Motley, A.; Robinson, M.; Robinson, C.; Haucke, V.; Owen, D.J. Phosphatidylinositol-(4,5)-Bisphosphate Regulates Sorting Signal Recognition by the Clathrin-Associated Adaptor Complex AP2. Mol. Cell 2005, 18, 519–531. [Google Scholar] [CrossRef] [Green Version]
- Wenk, M.R.; De Camilli, P. Protein-Lipid Interactions and Phosphoinositide Metabolism in Membrane Traffic: Insights from Vesicle Recycling in Nerve Terminals. Proc. Natl. Acad. Sci. USA 2004, 101, 8262–8269. [Google Scholar] [CrossRef] [Green Version]
- Simonsen, A.; Wurmser, A.E.; Emr, S.D.; Stenmark, H. The Role of Phosphoinositides in Membrane Transport. Curr. Opin. Cell Biol. 2001, 13, 485–492. [Google Scholar] [CrossRef]
- Holz, R.W.; Hlubek, M.D.; Sorensen, S.D.; Fisher, S.K.; Balla, T.; Ozaki, S.; Prestwich, G.D.; Stuenkel, E.L.; Bittner, M.A. A Pleckstrin Homology Domain Specific for Phosphatidylinositol 4,5- Bisphosphate (PtdIns-4,5-P2) and Fused to Green Fluorescent Protein Identifies Plasma Membrane PtdIns-4,5-P2 as Being Important in Exocytosis. J. Biol. Chem. 2000, 275, 17878–17885. [Google Scholar] [CrossRef] [Green Version]
- Martin, T.F.J. PI(4,5)P2 Regulation of Surface Membrane Traffic. Curr. Opin. Cell Biol. 2001, 13, 493–499. [Google Scholar] [CrossRef]
- Hilgemann, D.W.; Feng, S.; Nasuhoglu, C. The Complex and Intriguing Lives of PIP2 with Ion Channels and Transporters. Sci. STKE 2001, 2001, re19. [Google Scholar] [CrossRef]
- Yarar, D.; Surka, M.C.; Leonard, M.C.; Schmid, S.L. SNX9 Activities Are Regulated by Multiple Phosphoinositides Through Both PX and BAR Domains. Traffic 2008, 9, 133–146. [Google Scholar] [CrossRef]
- Song, X.; Xu, W.; Zhang, A.; Huang, G.; Liang, X.; Virbasius, J.V.; Czech, M.P.; Zhou, G.W. Phox Homology Domains Specifically Bind Phosphatidylinositol Phosphates. Biochemistry 2001, 40, 8940–8944. [Google Scholar] [CrossRef]
- Chiang, S.-H.; Hwang, J.; Legendre, M.; Zhang, M.; Kimura, A.; Saltiel, A.R. TCGAP, a Multidomain Rho GTPase-Activating Protein Involved in Insulin-Stimulated Glucose Transport. EMBO J. 2003, 22, 2679–2691. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Tucker, W.C.; Chapman, E.R. PIP2 Increases the Speed of Response of Synaptotagmin and Steers Its Membrane-Penetration Activity toward the Plasma Membrane. Nat. Struct. Mol. Biol. 2004, 11, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Itoh, T.; Koshiba, S.; Kigawa, T.; Kikuchi, A.; Yokoyama, S.; Takenawa, T. Role of the ENTH Domain in Phosphatidylinositol-4,5-Bisphosphate Binding and Endocytosis. Science 2001, 291, 1047–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santagata, S.; Boggon, T.J.; Baird, C.L.; Gomez, C.A.; Zhao, J.; Shan, W.S.; Myszka, D.G.; Shapiro, L. G-Protein Signaling Through Tubby Proteins. Science 2001, 292, 2041–2050. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Ferguson, K.M.; O’Brien, R.; Sigler, P.B.; Schlessinger, J. Specific and High-Affinity Binding of Inositol Phosphates to an Isolated Pleckstrin Homology Domain. Proc. Natl. Acad. Sci. USA 1995, 92, 10472–10476. [Google Scholar] [CrossRef] [Green Version]
- Achiriloaie, M.; Barylko, B.; Albanesi, J.P. Essential Role of the Dynamin Pleckstrin Homology Domain in Receptor-Mediated Endocytosis. Mol. Cell. Biol. 1999, 19, 1410–1415. [Google Scholar] [CrossRef] [Green Version]
- Lemmon, M.A.; Ferguson, K.M. Signal-Dependent Membrane Targeting by Pleckstrin Homology (PH) Domains. Biochem. J. 2000, 350, 1–18. [Google Scholar] [CrossRef]
- Garcia, P.; Gupta, R.; Shah, S.; Morris, A.J.; Rudge, S.A.; Scarlata, S.; Petrova, V.; McLaughlin, S.; Rebecchi, M.J.J. The Pleckstrin Homology Domain of Phospholipase C-61 Binds with High Affinity to Phosphatidylinositol 4,5-Bisphosphate in Bilayer Membranes? Biochemistry 1995, 34, 16228–16234. [Google Scholar] [CrossRef]
- Lemmon, M.A. Pleckstrin Homology (PH) Domains and Phosphoinositides. Biochem. Soc. Symp. 2007, 74, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, K.M.; Lemmon, M.A.; Schlessinger, J.; Sigler, P.B. Crystal Structure at 2.2 Å Resolution of the Pleckstrin Homology Domain from Human Dynamin. Cell 1994, 79, 199–209. [Google Scholar] [CrossRef]
- Ferguson, K.M.; Lemmon, M.A.; Schlessinger, J.; Sigler, P.B. Structure of the High Affinity Complex of Inositol Trisphosphate with a Phospholipase C Pleckstrin Homology Domain. Cell 1995, 83, 1037–1046. [Google Scholar] [CrossRef] [Green Version]
- Clapham, D.E. Calcium Signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [Green Version]
- Sarmento, M.J.; Coutinho, A.; Fedorov, A.; Prieto, M.; Fernandes, F. Ca2+ Induces PI(4,5)P2 Clusters on Lipid Bilayers at Physiological PI(4,5)P2 and Ca2+ Concentrations. Biochim. Biophys. Acta-Biomembr. 2014, 1838, 822–830. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-H.; Collins, A.; Guo, L.; Smith-Dupont, K.B.; Gai, F.; Svitkina, T.; Janmey, P.A. Divalent Cation-Induced Cluster Formation by Polyphosphoinosited in Model Membranes. J. Am. Chem. Soc. 2012, 134, 3387–3395. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Slochower, D.R.; Janmey, P.A. Counterion-Mediated Cluster Formation by Polyphosphoinositides. Chem. Phys. Lipids 2014, 182, 38–51. [Google Scholar] [CrossRef] [Green Version]
- Ellenbroek, W.G.; Wang, Y.-H.H.; Christian, D.A.; Discher, D.E.; Janmey, P.A.; Liu, A.J. Divalent Cation-Dependent Formation of Electrostatic PIP2 Clusters in Lipid Monolayers. Biophys. J. 2011, 101, 2178–2184. [Google Scholar] [CrossRef] [Green Version]
- Sarmento, M.J.; Coutinho, A.; Fedorov, A.; Prieto, M.; Fernandes, F. Membrane Order Is a Key Regulator of Divalent Cation-Induced Clustering of PI(3,5)P2 and PI(4,5)P2. Langmuir 2017, 33, 12463–12477. [Google Scholar] [CrossRef]
- Wen, Y.; Vogt, V.M.; Feigenson, G.W. Multivalent Cation-Bridged PI(4,5)P2 Clusters Form at Very Low Concentrations. Biophys. J. 2018, 114, 2630–2639. [Google Scholar] [CrossRef] [Green Version]
- Borges-Araújo, L.; Domingues, M.M.; Fedorov, A.; Santos, N.C.; Melo, M.N.; Fernandes, F. Acyl-Chain Saturation Regulates the Order of Phosphatidylinositol 4,5-Bisphosphate Nanodomains. Commun. Chem. 2021, 4, 164. [Google Scholar] [CrossRef]
- Seo, J.B.; Jung, S.-R.; Huang, W.; Zhang, Q.; Koh, D.-S. Charge Shielding of PIP2 by Cations Regulates Enzyme Activity of Phospholipase, C. PLoS ONE 2015, 10, e0144432. [Google Scholar] [CrossRef] [Green Version]
- Bilkova, E.; Pleskot, R.; Rissanen, S.; Sun, S.; Czogalla, A.; Cwiklik, L.; Róg, T.; Vattulainen, I.; Cremer, P.S.; Jungwirth, P.; et al. Calcium Directly Regulates Phosphatidylinositol 4,5-Bisphosphate Headgroup Conformation and Recognition. J. Am. Chem. Soc. 2017, 139, 4019–4024. [Google Scholar] [CrossRef]
- Zacharias, D.A.; Violin, J.D.; Newton, A.C.; Tsien, R.Y. Partitioning of Lipid-Modified Monomeric GFPs into Membrane Microdomains of Live Cells. Science 2002, 296, 913–916. [Google Scholar] [CrossRef] [Green Version]
- Van der Wal, J.; Habets, R.; Varnai, P.; Balla, T.; Jalink, K. Monitoring Agonist-Induced Phospholipase C Activation in Live Cells by Fluorescence Resonance Energy Transfer. J. Biol. Chem. 2001, 276, 15337–15344. [Google Scholar] [CrossRef] [Green Version]
- Mayer, L.D.; Hope, M.J.; Cullis, P.R. Vesicles of Variable Sizes Produced by a Rapid Extrusion Procedure. Biochim. Biophys. Acta 1986, 858, 161–168. [Google Scholar] [CrossRef]
- Weinberger, A.; Weinberger, A.; Tsai, F.-C.; Koenderink, G.H.H.; Schmidt, T.F.F.; Itri, R.; Meier, W.; Schmatko, T.; Schröder, A.; Marques, C. Gel-Assisted Formation of Giant Unilamellar Vesicles. Biophys. J. 2013, 105, 154–164. [Google Scholar] [CrossRef] [Green Version]
- Sarmento, M.J.; Prieto, M.; Fernandes, F. Reorganization of Lipid Domain Distribution in Giant Unilamellar Vesicles upon Immobilization with Different Membrane Tethers. Biochim. Biophys. Acta 2012, 1818, 2605–2615. [Google Scholar] [CrossRef] [Green Version]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy; Lakowicz, J.R., Ed.; Springer: Boston, MA, USA, 2006; ISBN 978-0-387-31278-1. [Google Scholar]
- Shaner, N.C.; Steinbach, P.A.; Tsien, R.Y. A Guide to Choosing Fluorescent Proteins. Nat. Methods 2005, 2, 905–909. [Google Scholar] [CrossRef]
- Shi, X.; Basran, J.; Seward, H.E.; Childs, W.; Bagshaw, C.R.; Boxer, S.G. Anomalous Negative Fluorescence Anisotropy in Yellow Fluorescent Protein (YFP 10C): Quantitative Analysis of FRET in YFP Dimers. Biochemistry 2007, 46, 14403–14417. [Google Scholar] [CrossRef] [Green Version]
- Knight, J.D.; Lerner, M.G.; Marcano-Velázquez, J.G.; Pastor, R.W.; Falke, J.J. Single Molecule Diffusion of Membrane-Bound Proteins: Window into Lipid Contacts and Bilayer Dynamics. Biophys. J. 2010, 99, 2879–2887. [Google Scholar] [CrossRef] [Green Version]
- Skakun, V.V.; Digris, A.V.; Apanasovich, V.V. Global Analysis of Autocorrelation Functions and Photon Counting Distributions in Fluorescence Fluctuation Spectroscopy. Methods Mol. Biol. 2014, 1076, 719–741. [Google Scholar] [CrossRef]
- Huang, B.; Perroud, T.D.; Zare, R.N. Photon Counting Histogram: One-Photon Excitation. ChemPhysChem 2004, 5, 1523–1531. [Google Scholar] [CrossRef]
- Ries, J.; Schwille, P. New Concepts for Fluorescence Correlation Spectroscopy on Membranes. Phys. Chem. Chem. Phys. 2008, 10, 3487–3497. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Müller, J.D.; So, P.T.C.; Gratton, E. The Photon Counting Histogram in Fluorescence Fluctuation Spectroscopy. Biophys. J. 1999, 77, 553–567. [Google Scholar] [CrossRef] [Green Version]
- Borges-Araújo, L.; Souza, P.C.T.; Fernandes, F.; Melo, M.N. Improved Parameterization of Phosphatidylinositide Lipid Headgroups for the Martini 3 Coarse-Grain Force Field. J. Chem. Theory Comput. 2022, 18, 357–373. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ca2+ Concentration (μM) | F1 (Monomeric Population) (%) | F2 (Oligomeric Population) (%) |
|---|---|---|
| 0 | 71.3 ± 5.3 | 28.7 ± 5.3 |
| 20 | 99.3 ± 0.2 | 0.7 ± 0.2 |
| 50 | 82.5 ± 5.7 | 17.5 ± 5.7 |
| 100 | 86.5 ± 3.0 | 13.5 ± 3.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borges-Araújo, L.; Monteiro, M.E.; Mil-Homens, D.; Bernardes, N.; Sarmento, M.J.; Coutinho, A.; Prieto, M.; Fernandes, F. Impact of Ca2+-Induced PI(4,5)P2 Clusters on PH-YFP Organization and Protein-Protein Interactions. Biomolecules 2022, 12, 912. https://doi.org/10.3390/biom12070912
Borges-Araújo L, Monteiro ME, Mil-Homens D, Bernardes N, Sarmento MJ, Coutinho A, Prieto M, Fernandes F. Impact of Ca2+-Induced PI(4,5)P2 Clusters on PH-YFP Organization and Protein-Protein Interactions. Biomolecules. 2022; 12(7):912. https://doi.org/10.3390/biom12070912
Chicago/Turabian StyleBorges-Araújo, Luís, Marina E. Monteiro, Dalila Mil-Homens, Nuno Bernardes, Maria J. Sarmento, Ana Coutinho, Manuel Prieto, and Fábio Fernandes. 2022. "Impact of Ca2+-Induced PI(4,5)P2 Clusters on PH-YFP Organization and Protein-Protein Interactions" Biomolecules 12, no. 7: 912. https://doi.org/10.3390/biom12070912
APA StyleBorges-Araújo, L., Monteiro, M. E., Mil-Homens, D., Bernardes, N., Sarmento, M. J., Coutinho, A., Prieto, M., & Fernandes, F. (2022). Impact of Ca2+-Induced PI(4,5)P2 Clusters on PH-YFP Organization and Protein-Protein Interactions. Biomolecules, 12(7), 912. https://doi.org/10.3390/biom12070912