
Toxicity Assessment of an Anti-Cancer Drug of p-Toluene Sulfonamide in Zebrafish Larvae Based on Cardiovascular and Locomotion Activities


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Zebrafish Maintenance
2.2. Acute Toxicity Test
2.3. Cardiotoxicity Assessment
2.4. Vascular Performance Assessment
2.5. Photomotor Response and Morphology Assessment
2.6. Vibrational Startle Response Assessment
2.7. Respiratory Rate Assessment
2.8. Molecular Docking of p-TSA to Its Target Proteins
2.9. Gene Expression Analysis
2.10. Statistical Analyses
3. Results
3.1. Acute Toxicity Test of p-TSA
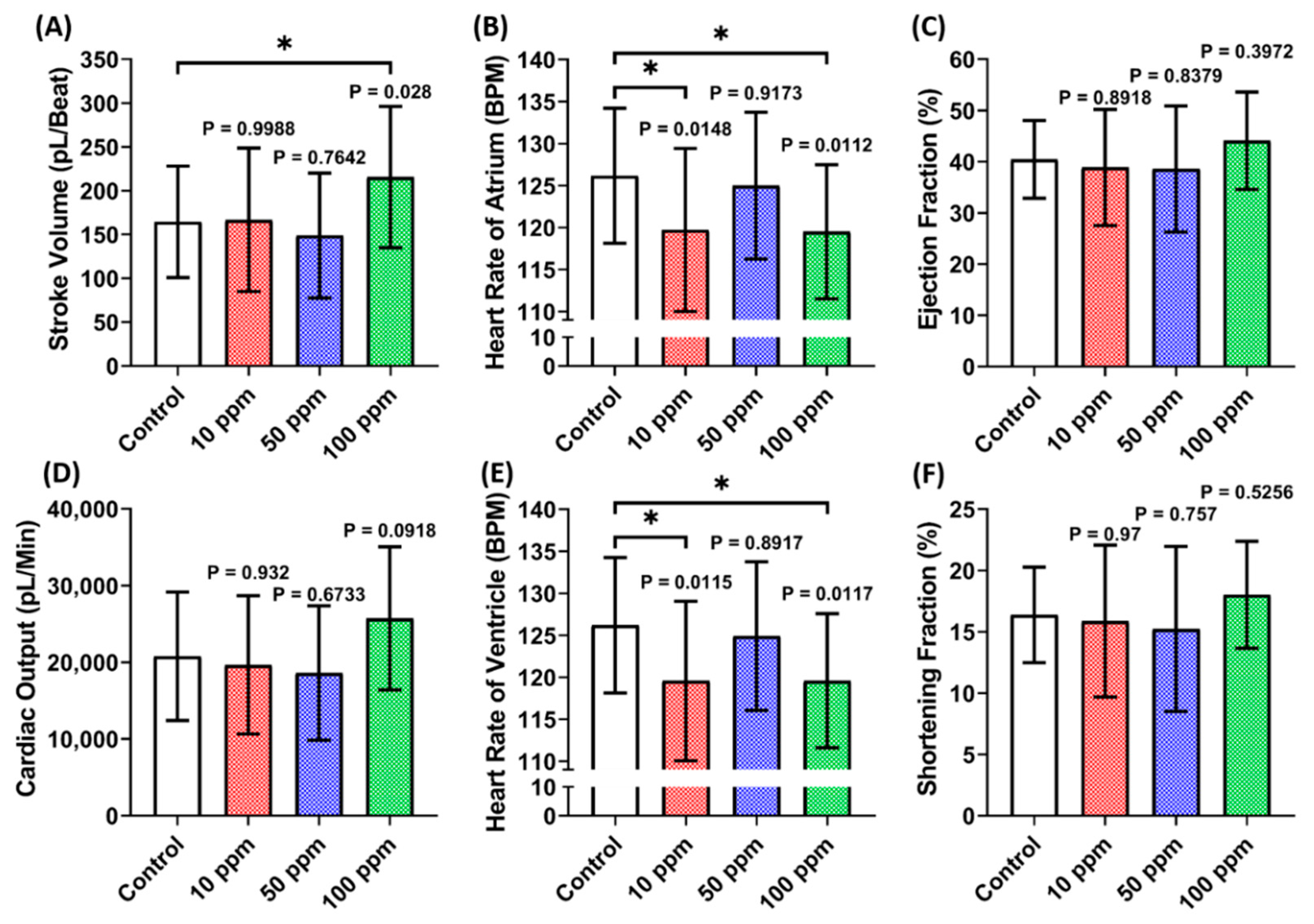
3.2. Cardiac Performance Assessment of p-TSA in Zebrafish
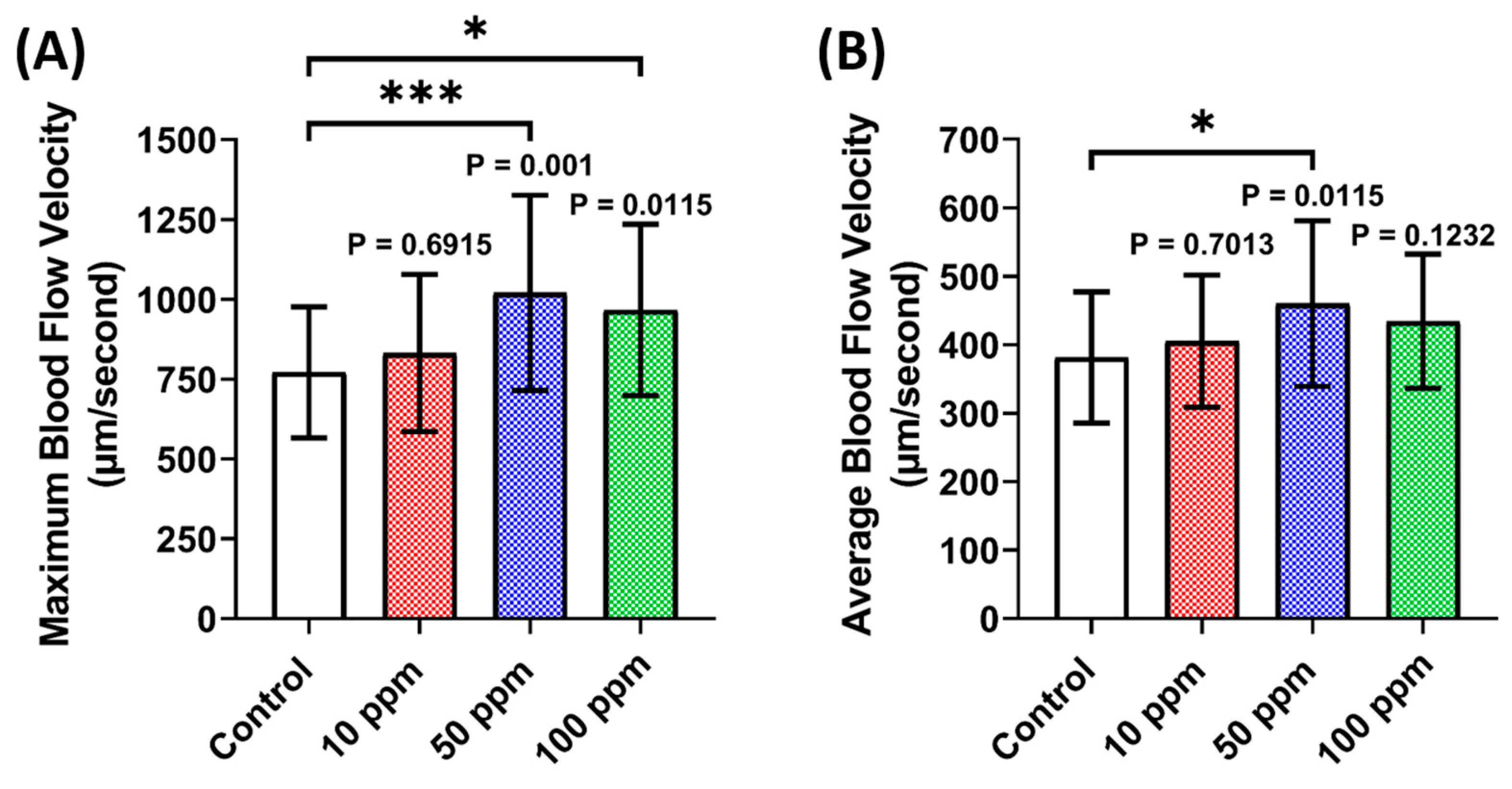
3.3. Vascular Performance Assessment of p-TSA in Zebrafish
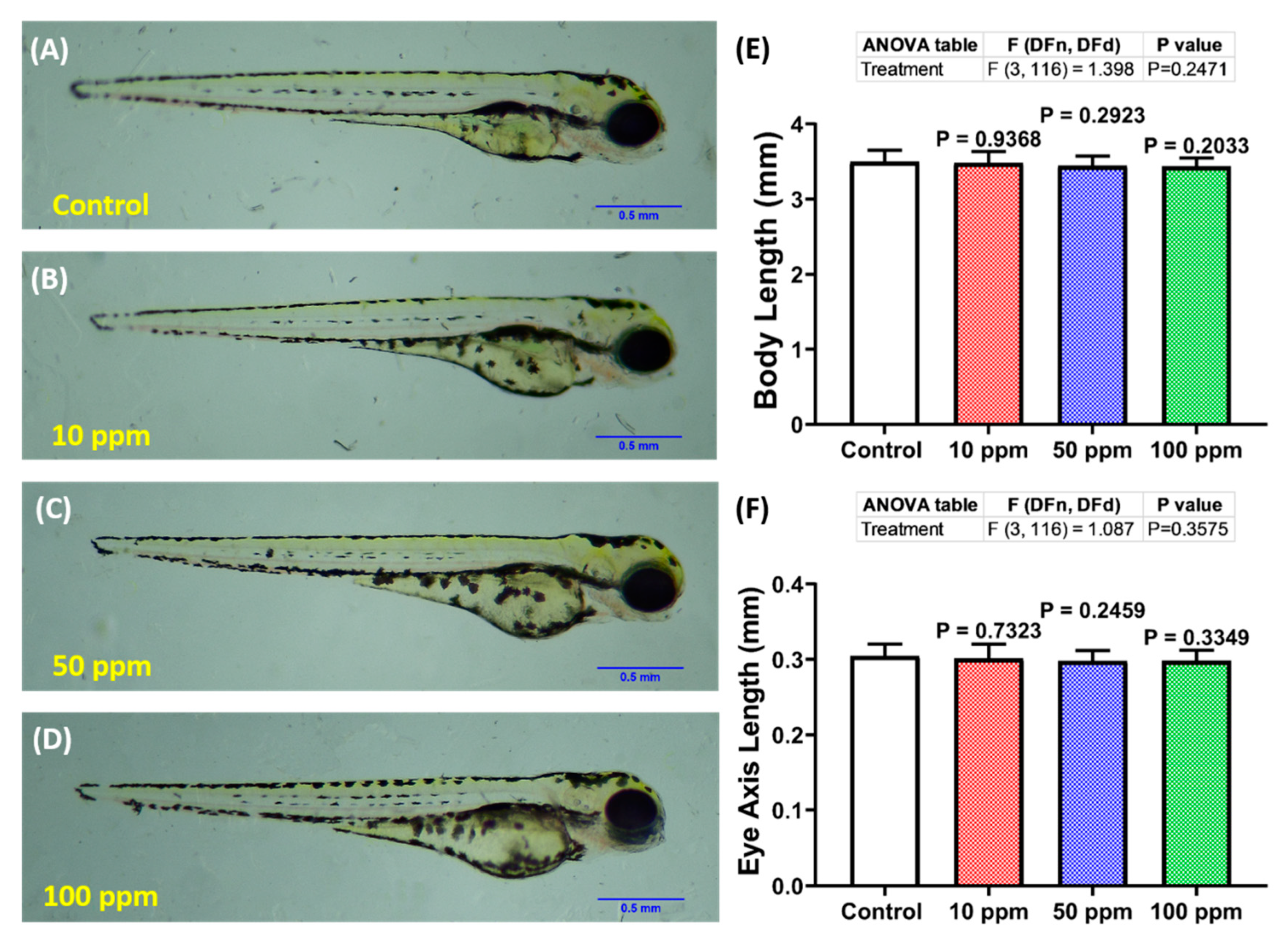
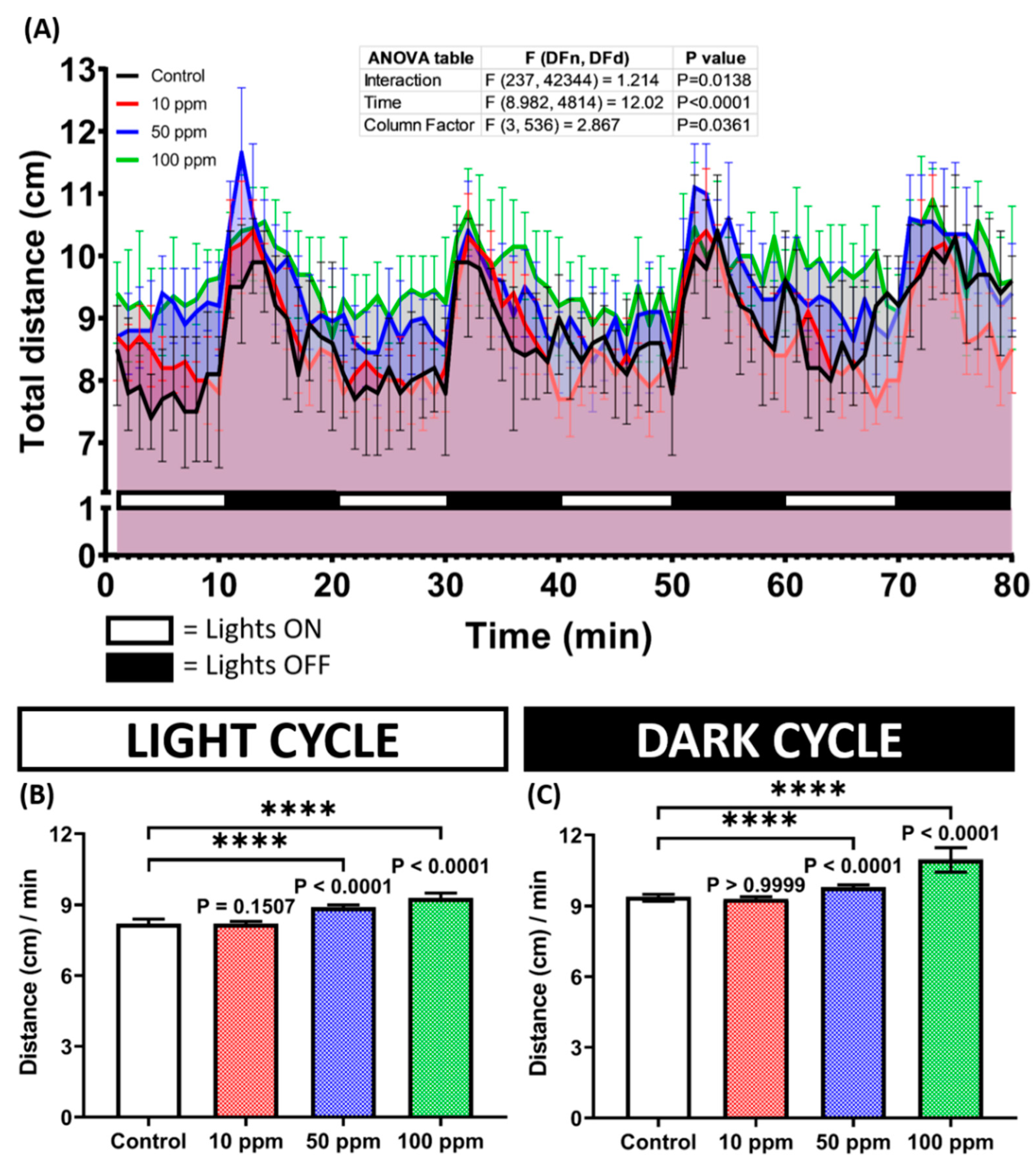
3.4. Photomotor Response and Morphology Assessment of p-TSA in Zebrafish
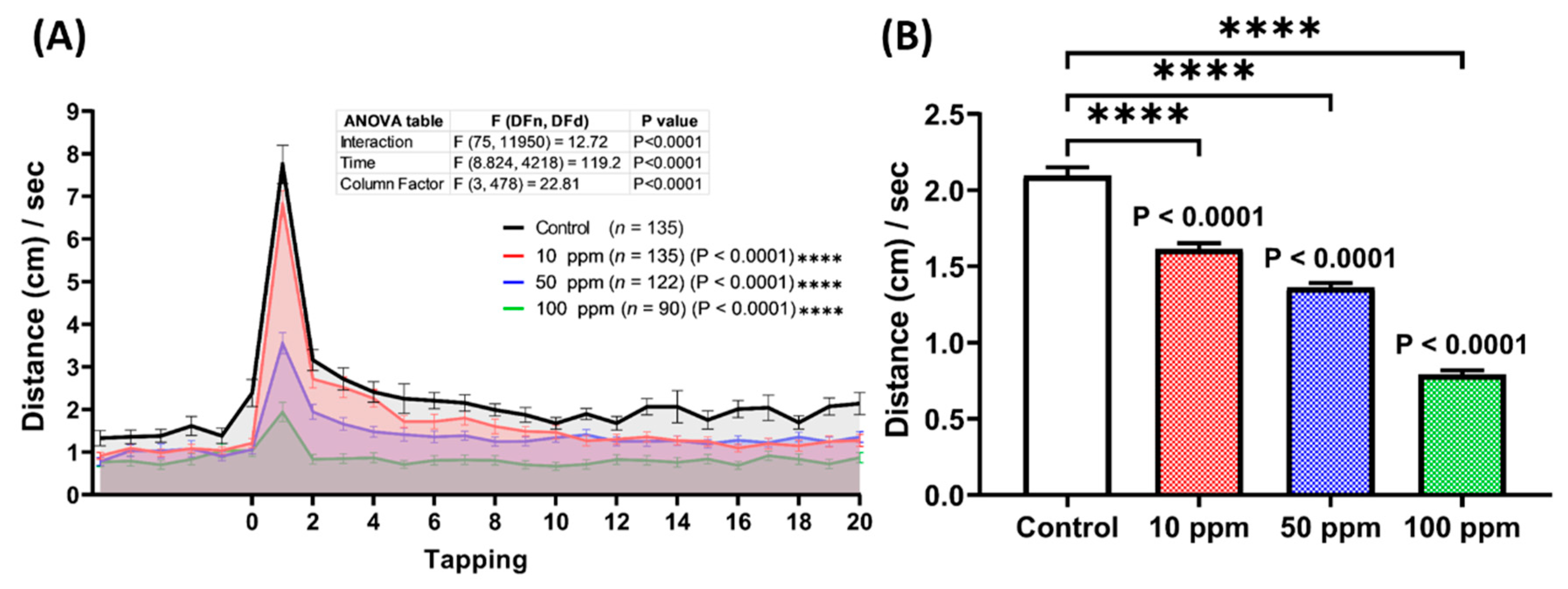
3.5. Startle Reflex Assessment of p-TSA in Zebrafish
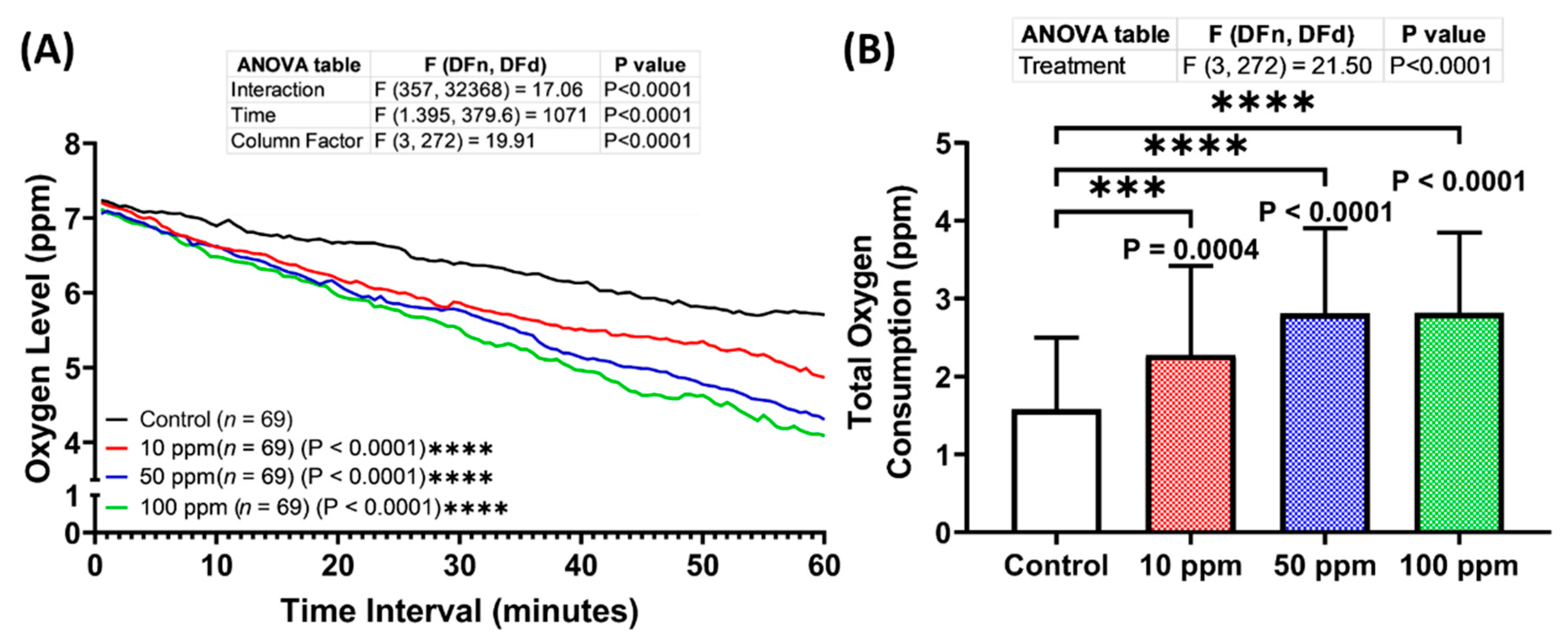
3.6. Respiratory Rate Assessment of p-TSA in Zebrafish
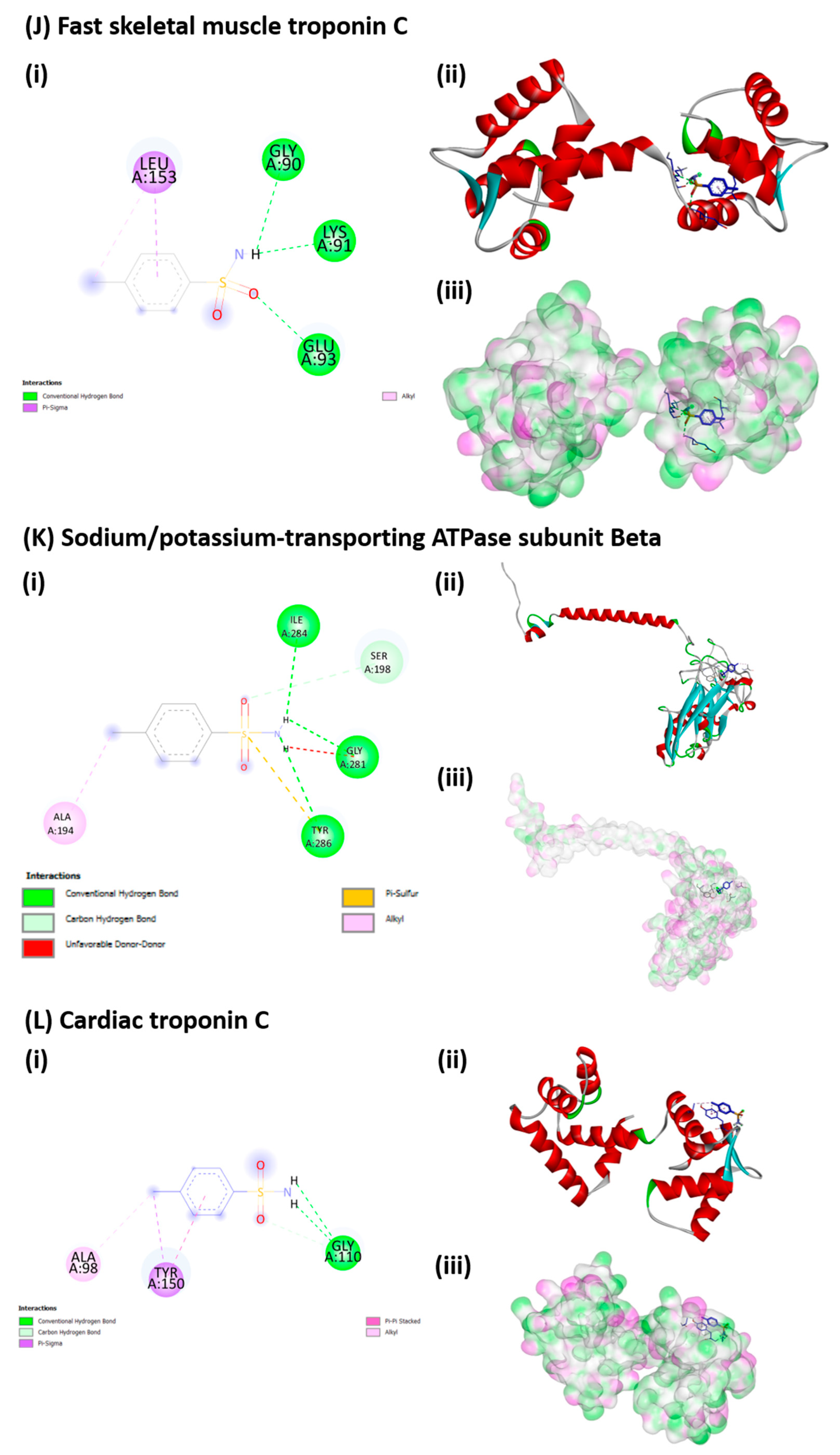
3.7. Molecular Docking of p-TSA to Its Target Protein
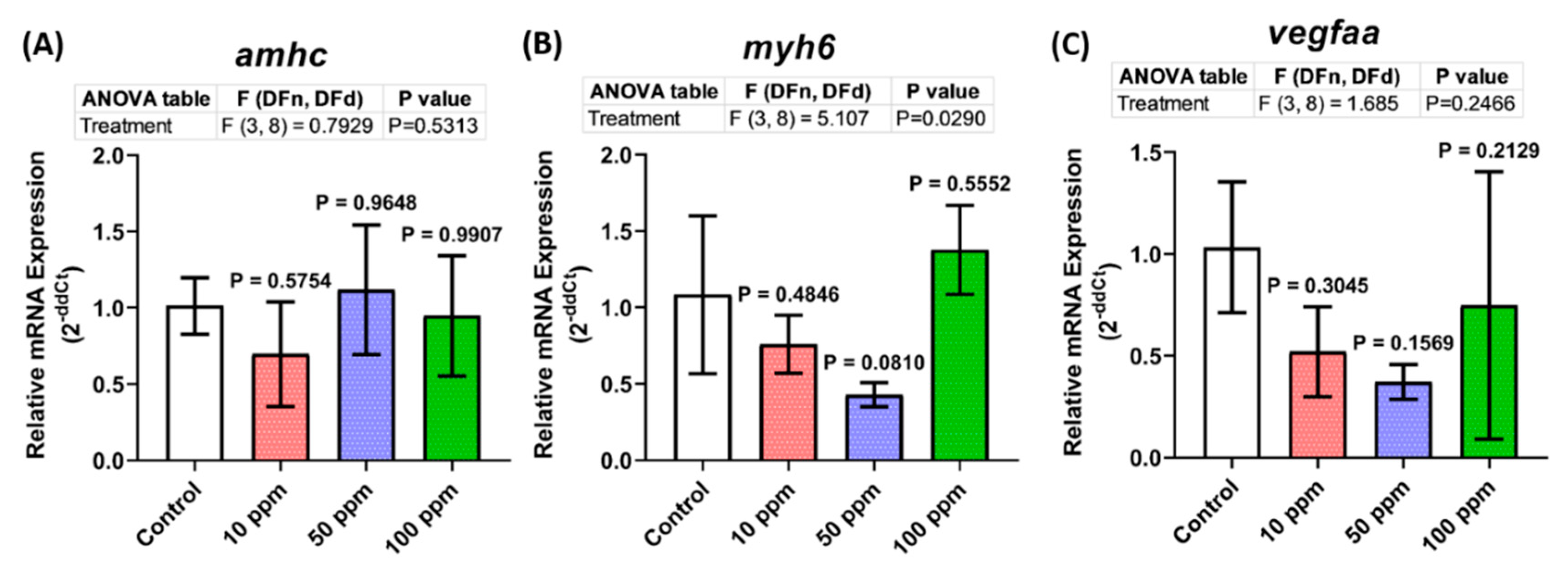
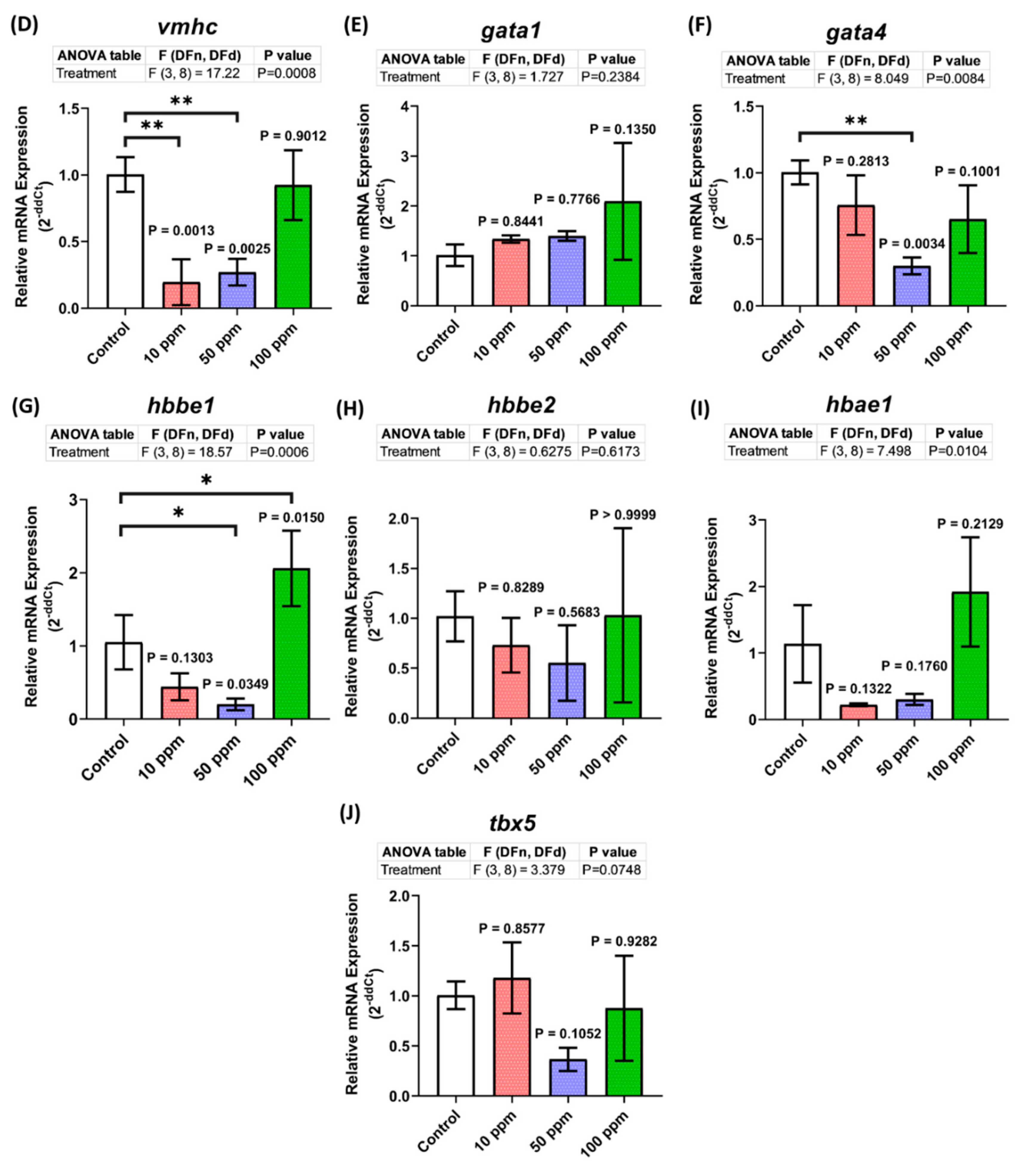
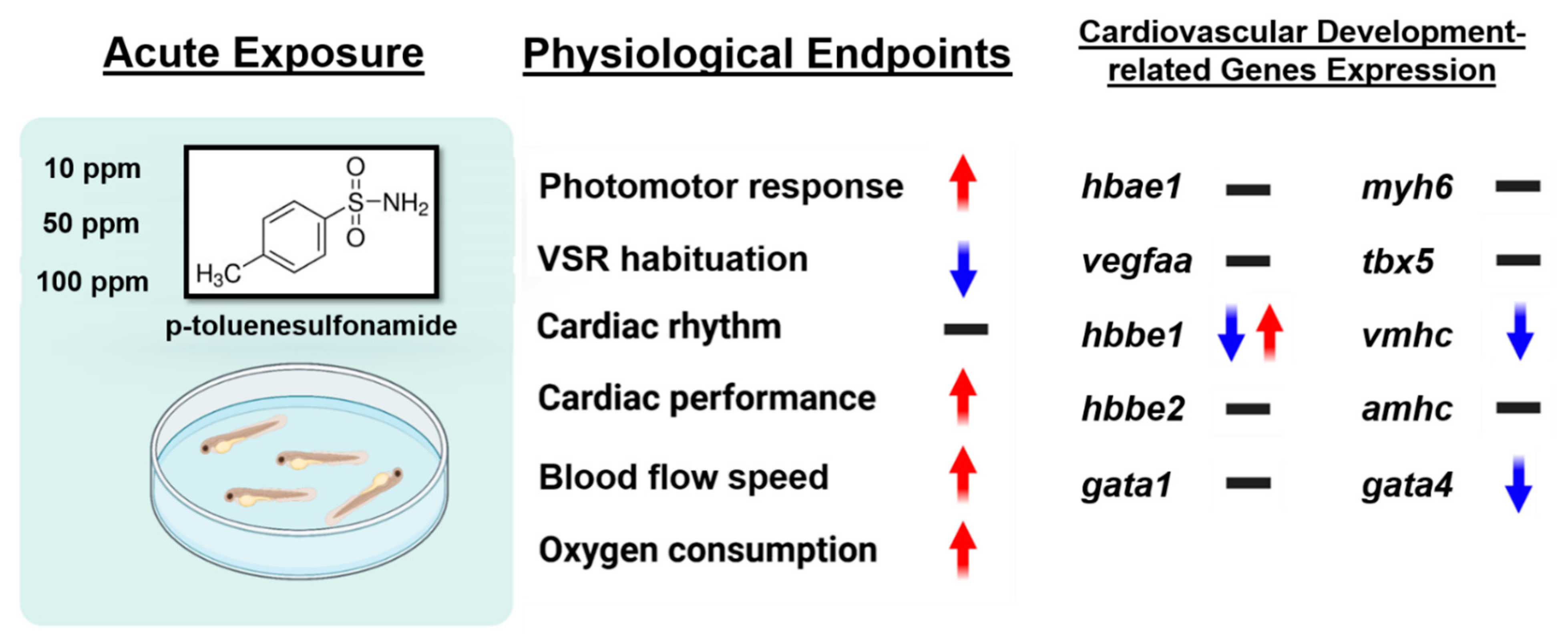
3.8. Cardiovascular Development-Related Genes Expression
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequences of the Forward Primer (5′-3′) | Sequences of the Reversed Primer (5′-3′) |
| myh6 | F: CACCAGCAGACACTGGATG | R: GCTCCAAGTCCATTCTGAC |
| txb5 | F: ATTCGCCGATAACAAATGG | R: CGCCTTGACGATGTGGAT |
| vmhc | F: GAAGAGGCAGAGGCATCACT | R: AATTGCGTTTGCTCTGCTCC |
| amhc | F: AAGCCACTACCGCCTCTCTA | R: TTTGAGGCAAGGTCGTCCAA |
| gata4 | F: TCCAGGCGGGTGGGTTTATC | R:TGTCTGGTTCAGTCTTGATGGGTC |
| vegfaa | F: AAAAGAGTGCGTGCAAGACC | R: GACGTTTCGTGTCTCTGTCG |
| hbbe1 | F: GCTCTGGCAAGGTGTCTCAT | R: TTCTTCACTGCCAGCTCCAG |
| hbbe2 | F: ACTATGAGGAGGCTGGAC | R: CGGCGTAGGTGTTCTTG |
| hbae1 | F: CCAGGATGTTGATTGTCTAC | R: CAGTCTTGCCGTGTTTC |
| gata1 | F: GAGACTGACCTACTGCCATCG | R: TCCCAGAATTGACTGAGATGAG |
References
- Yang, S.; Cha, J.; Carlson, K. Quantitative determination of trace concentrations of tetracycline and sulfonamide antibiotics in surface water using solid-phase extraction and liquid chromatography/ion trap tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2004, 18, 2131–2145. [Google Scholar] [CrossRef]
- Shimizu, A.; Takada, H.; Koike, T.; Takeshita, A.; Saha, M.; Nakada, N.; Murata, A.; Suzuki, T.; Suzuki, S.; Chiem, N.H. Ubiquitous occurrence of sulfonamides in tropical Asian waters. Sci. Total Environ. 2013, 452, 108–115. [Google Scholar] [CrossRef]
- Chen, K.; Zhou, J. Occurrence and behavior of antibiotics in water and sediments from the Huangpu River, Shanghai, China. Chemosphere 2014, 95, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-Q.; Ying, G.-G.; Pan, C.-G.; Liu, Y.-S.; Zhao, J.-L. Comprehensive evaluation of antibiotics emission and fate in the river basins of China: Source analysis, multimedia modeling, and linkage to bacterial resistance. Environ. Sci. Technol. 2015, 49, 6772–6782. [Google Scholar] [CrossRef] [PubMed]
- Daniel, A.B.; Aruldhas, D.; Joe, I.H.; Sasi, B.A. Experimental and computational approach on p-toluenesulfonamide and its derivatives. J. Mol. Struct. 2020, 1218, 128503. [Google Scholar] [CrossRef]
- Yuan, R.-X.; Xiong, R.-G.; Chen, Z.-F.; Zhang, P.; Ju, H.-X.; Dai, Z.; Guo, Z.-J.; Fun, H.-K.; You, X.-Z. Crystal structure of zinc (II) 2-sulfanilamidopyrimidine: A widely used topical burn drug. J. Chem. Soc. Dalton Trans. 2001, 6, 774–776. [Google Scholar] [CrossRef]
- Maren, T.H. Relations between structure and biological activity of sulfonamides. Annu. Rev. Pharmacol. Toxicol. 1976, 16, 309–327. [Google Scholar] [CrossRef] [PubMed]
- Tunc, T. Synthesis and antimicrobial study of new benzimidazole Schiff bases bearing p-toluene sulfonamide moiety. J. Chil. Chem. Soc. 2019, 64, 4590–4592. [Google Scholar] [CrossRef]
- Wolff, M.E. Burger’s medicinal chemistry and drug discovery. Am. J. Ther. 1996, 3, 608. [Google Scholar] [CrossRef]
- Abbate, F.; Casini, A.; Owa, T.; Scozzafava, A.; Supuran, C.T. Carbonic anhydrase inhibitors: E7070, a sulfonamide anticancer agent, potently inhibits cytosolic isozymes I and II, and transmembrane, tumor-associated isozyme IX. Bioorg. Med. Chem. Lett. 2004, 14, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Akurathi, V.; Dubois, L.; Celen, S.; Lieuwes, N.G.; Chitneni, S.K.; Cleynhens, B.J.; Innocenti, A.; Supuran, C.T.; Verbruggen, A.M.; Lambin, P. Development and biological evaluation of 99mTc-sulfonamide derivatives for in vivo visualization of CA IX as surrogate tumor hypoxia markers. Eur. J. Med. Chem. 2014, 71, 374–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owa, T.; Nagasu, T. Novel sulphonamide derivatives for the treatment of cancer. Expert Opin. Ther. Pat. 2000, 10, 1725–1740. [Google Scholar] [CrossRef]
- Ghorab, M.M.; Ragab, F.A.; Heiba, H.I.; Agha, H.M. Synthesis of Some Novel Sulfonamides Containing Biologically Active Alkanoic Acid, Acetamide, Thiazole, and Pyrrole Moieties of Expected Antitumor and Radiosensitizing Activities. J. Basic Appl. Chem. 2011, 1, 8–14. [Google Scholar]
- Ghorab, M.M.; Ragab, F.A.; Heiba, H.I.; Arafa, R.K.; El-Hossary, E.M. Docking study, in vitro anticancer screening and radiosensitizing evaluation of some new fluorine-containing quinoline and pyrimidoquinoline derivatives bearing a sulfonamide moiety. Med. Chem. Res. 2011, 20, 388–400. [Google Scholar] [CrossRef]
- Al-Said, M.S.; Ghorab, M.M.; Al-Qasoumi, S.I.; El-Hossary, E.M.; Noaman, E. Synthesis and in vitro anticancer screening of some novel 4-[2-amino-3-cyano-4-substituted-5,6,7,8-tetrahydroquinolin-1-(4H)-yl] benzenesulfonamides. Eur. J. Med. Chem. 2010, 45, 3011–3018. [Google Scholar] [CrossRef] [PubMed]
- Lal, J.; Gupta, S.K.; Thavaselvam, D.; Agarwal, D.D. Biological activity, design, synthesis and structure activity relationship of some novel derivatives of curcumin containing sulfonamides. Eur. J. Med. Chem. 2013, 64, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Gawin, R.; De Clercq, E.; Naesens, L.; Koszytkowska-Stawińska, M. Synthesis and antiviral evaluation of acyclic azanucleosides developed from sulfanilamide as a lead structure. Bioorg. Med. Chem. 2008, 16, 8379–8389. [Google Scholar] [CrossRef] [PubMed]
- Chandna, N.; Kumar, S.; Kaushik, P.; Kaushik, D.; Roy, S.; Gupta, G.; Jachak, S.; Kapoor, J.; Sharma, P. Synthesis of novel celecoxib analogues by bioisosteric replacement of sulfonamide as potent anti-inflammatory agents and cyclooxygenase inhibitors. Bioorg. Med. Chem. 2013, 21, 4581–4590. [Google Scholar] [CrossRef] [PubMed]
- Thornber, C. Isosterism and molecular modification in drug design. Chem. Soc. Rev. 1979, 8, 563–580. [Google Scholar] [CrossRef]
- Supuran, C.T.; Scozzafava, A.; Mastrolorenzo, A. Bacterial proteases: Current therapeutic use and future prospects for the development of new antibiotics. Expert Opin. Ther. Pat. 2001, 11, 221–259. [Google Scholar] [CrossRef]
- Jana, U.; Maiti, S.; Biswas, S. An efficient FeCl3-catalyzed amidation reaction of secondary benzylic and allylic alcohols with carboxamides or p-toluenesulfonamide. Tetrahedron Lett. 2008, 49, 858–862. [Google Scholar] [CrossRef]
- Meinertz, J.R.; Stehly, G.R.; Gingerich, W.H.; Greseth, S.L. Performance of a proposed determinative method for p-TSA in rainbow trout fillet tissue and bridging the proposed method with a method for total chloramine-T residues in rainbow trout fillet tissue. J. AOAC Int. 2001, 84, 1332–1336. [Google Scholar] [CrossRef] [Green Version]
- Richter, D.; Dünnbier, U.; Massmann, G.; Pekdeger, A. Quantitative determination of three sulfonamides in environmental water samples using liquid chromatography coupled to electrospray tandem mass spectrometry. J. Chromatogr. A 2007, 1157, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Meffe, R.; Kohfahl, C.; Holzbecher, E.; Massmann, G.; Richter, D.; Dünnbier, U.; Pekdeger, A. Modelling the removal of p-TSA (para-toluenesulfonamide) during rapid sand filtration used for drinking water treatment. Water Res. 2010, 44, 205–213. [Google Scholar] [CrossRef]
- Yang, T.-M.; Su, C.-S.; Li, J.-S.; Lu, K.-T.; Yeh, T.-F. Recrystallization and micronization of p-toluenesulfonamide using the rapid expansion of supercritical solution (RESS) process. Crystals 2019, 9, 449. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Gao, Y.; Guan, W.; Huang, L.; Xu, X.; Zhang, C.; Chen, X.; Wu, Y.; Zeng, G.; Zhong, N. Antitumor effect of para-toluenesulfonamide against lung cancer xenograft in a mouse model. J. Thorac. Dis. 2013, 5, 472. [Google Scholar]
- Hsu, J.-L.; Leu, W.-J.; Hsu, L.-C.; Liu, S.-P.; Zhong, N.-S.; Guh, J.-H. Para-toluenesulfonamide induces anti-tumor activity through Akt-dependent and-independent mTOR/p70S6K pathway: Roles of lipid raft and cholesterol contents. Front. Pharmacol. 2018, 9, 1223. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Ying, W.; Yang, H.; Xu, X.; Shao, W.; Guan, Y.; Jiang, M.; Wu, Y.; Zhong, B.; Wang, D. Gemcitabine plus cisplatin chemotherapy with concurrent para-toluenesulfonamide local injection therapy for peripherally advanced nonsmall cell lung cancer larger than 3 cm in the greatest dimension. Anti-Cancer Drugs 2009, 20, 838–844. [Google Scholar] [CrossRef]
- Richter, D.; Massmann, G.; Dünnbier, U. Behaviour and biodegradation of sulfonamides (p-TSA, o-TSA, BSA) during drinking water treatment. Chemosphere 2008, 71, 1574–1581. [Google Scholar] [CrossRef]
- Lin, T.; Yu, S.; Chen, Y.; Chen, W. Integrated biomarker responses in zebrafish exposed to sulfonamides. Environ. Toxicol. Pharmacol. 2014, 38, 444–452. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlahogianni, T.; Dassenakis, M.; Scoullos, M. Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants. Ecotoxicol. Environ. Saf. 2006, 64, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Van der Oost, R.; Beyer, J.; Vermeulen, N.P. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Yang, C.; Song, G.; Lim, W. A review of the toxicity in fish exposed to antibiotics. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 237, 108840. [Google Scholar] [CrossRef] [PubMed]
- Avdesh, A.; Chen, M.; Martin-Iverson, M.T.; Mondal, A.; Ong, D.; Rainey-Smith, S.; Taddei, K.; Lardelli, M.; Groth, D.M.; Verdile, G. Regular care and maintenance of a zebrafish (Danio rerio) laboratory: An introduction. J. Vis. Exp. 2012, 69, e4196. [Google Scholar] [CrossRef] [Green Version]
- Hua, J.; Vijver, M.G.; Richardson, M.K.; Ahmad, F.; Peijnenburg, W.J. Particle-specific toxic effects of differently shaped zinc oxide nanoparticles to zebrafish embryos (Danio rerio). Environ. Toxicol. Chem. 2014, 33, 2859–2868. [Google Scholar] [CrossRef]
- Saputra, F.; Uapipatanakul, B.; Lee, J.-S.; Hung, S.-M.; Huang, J.-C.; Pang, Y.-C.; Muñoz, J.E.R.; Macabeo, A.P.G.; Chen, K.H.-C.; Hsiao, C.-D. Co-Treatment of Copper Oxide Nanoparticle and Carbofuran Enhances Cardiotoxicity in Zebrafish Embryos. Int. J. Mol. Sci. 2021, 22, 8259. [Google Scholar] [CrossRef] [PubMed]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef] [Green Version]
- Santoso, F.; Sampurna, B.P.; Lai, Y.-H.; Liang, S.-T.; Hao, E.; Chen, J.-R.; Hsiao, C.-D. Development of a simple imagej-based method for dynamic blood flow tracking in zebrafish embryos and its application in drug toxicity evaluation. Inventions 2019, 4, 65. [Google Scholar] [CrossRef] [Green Version]
- Tinevez, J.-Y.; Perry, N.; Schindelin, J.; Hoopes, G.M.; Reynolds, G.D.; Laplantine, E.; Bednarek, S.Y.; Shorte, S.L.; Eliceiri, K.W. TrackMate: An open and extensible platform for single-particle tracking. Methods 2017, 115, 80–90. [Google Scholar] [CrossRef]
- Kristofco, L.A.; Cruz, L.C.; Haddad, S.P.; Behra, M.L.; Chambliss, C.K.; Brooks, B.W. Age matters: Developmental stage of Danio rerio larvae influences photomotor response thresholds to diazinion or diphenhydramine. Aquat. Toxicol. 2016, 170, 344–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siregar, P.; Audira, G.; Feng, L.-Y.; Lee, J.-H.; Santoso, F.; Yu, W.-H.; Lai, Y.-H.; Li, J.-H.; Lin, Y.-T.; Chen, J.-R. Pharmaceutical Assessment Suggests Locomotion Hyperactivity in Zebrafish Triggered by Arecoline Might Be Associated with Multiple Muscarinic Acetylcholine Receptors Activation. Toxins 2021, 13, 259. [Google Scholar] [CrossRef] [PubMed]
- Faria, M.; Prats, E.; Novoa-Luna, K.A.; Bedrossiantz, J.; Gómez-Canela, C.; Gómez-Oliván, L.M.; Raldúa, D. Development of a vibrational startle response assay for screening environmental pollutants and drugs impairing predator avoidance. Sci. Total Environ. 2019, 650, 87–96. [Google Scholar] [CrossRef]
- Bagatto, B.; Pelster, B.; Burggren, W. Growth and metabolism of larval zebrafish: Effects of swim training. J. Exp. Biol. 2001, 204, 4335–4343. [Google Scholar] [CrossRef] [PubMed]
- Prokop, J.; Duff, R.; Ball, H.; Copeland, D.; Londraville, R. Leptin and leptin receptor: Analysis of a structure to function relationship in interaction and evolution from humans to fish. Peptides 2012, 38, 326–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with PyRx. In Chemical Biology; Springer: Berlin/Heidelberg, Germany, 2015; pp. 243–250. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Adams, D.C.; Anthony, C.D. Using randomization techniques to analyse behavioural data. Anim. Behav. 1996, 51, 733–738. [Google Scholar] [CrossRef] [Green Version]
- Privitera, G.J. Student Study Guide with IBM® SPSS® Workbook for Essential Statistics for the Behavioral Sciences; SAGE Publications: New York, NY, USA, 2015. [Google Scholar]
- Salkind, N.J. Statistics for People Who (Think They) Hate Statistics: Excel 2007 Edition; SAGE: New York, NY, USA, 2009. [Google Scholar]
- El-Harbawi, M. Toxicity measurement of imidazolium ionic liquids using acute toxicity test. Procedia Chem. 2014, 9, 40–52. [Google Scholar] [CrossRef] [Green Version]
- Totten, G.; Cerf, J.; Bishop, R.J., Jr.; Webster, G. Recent results of biodegradability and toxicology studies of water-glycol hydraulic fluids. SAE Trans. 1997, 106, 241–246. [Google Scholar]
- USEPA. Technical Overview of Ecological Risk Assessment-Analysis Phase, Ecological Effects Characterization; US Environmental Protection Agency: Washington, DC, USA, 2019.
- Bowen, S.E.; Hannigan, J.H. Developmental toxicity of prenatal exposure to toluene. AAPS J. 2006, 8, E419–E424. [Google Scholar] [CrossRef]
- Pearson, M.A.; Rimsza, M.E.; Hoyme, H.E.; Seaver, L.H. Toluene embryopathy: Delineation of the phenotype and comparison with fetal alcohol syndrome. Pediatrics 1994, 93, 211–215. [Google Scholar] [CrossRef]
- Arnold, G.L.; Kirby, R.S.; Langendoerfer, S.; Wilkins-Haug, L. Toluene embryopathy: Clinical delineation and developmental follow-up. Pediatrics 1994, 93, 216–220. [Google Scholar] [CrossRef]
- Arito, H.; Tsuruta, H.; Nakagaki, K. Acute effects of toluene on circadian rhythms of sleep-wakefulness and brain monoamine metabolism in rats. Toxicology 1984, 33, 291–301. [Google Scholar] [CrossRef]
- Arito, H.; Tsuruta, H.; Nakagaki, K.; Tanaka, S. Partial insomnia, hyperactivity and hyperdipsia induced by repeated administration of toluene in rats: Their relation to brain monoamine metabolism. Toxicology 1985, 37, 99–110. [Google Scholar] [CrossRef]
- Korsak, Z.; Sokal, J.A.; Gorny, R. Toxic effects of combined exposure to toluene and m-xylene in animals. III. Subchronic inhalation study. Int. J. Occup. Med. Environ. Health 1992, 5, 27–33. [Google Scholar]
- Baydas, G.; Ozveren, F.; Akdemir, I.; Tuzcu, M.; Yasar, A. Learning and memory deficits in rats induced by chronic thinner exposure are reversed by melatonin. J. Pineal Res. 2005, 39, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Taş, U.; Ekici, F.; Koç, F.; Sogut, E.; Ayan, M.; Kuloğlu, T.; Arici, A.; Özyurt, B. Acute cardiotoxic effects of high dose toluene: An experimental study. Anatol. J. Cardiol. 2013, 13, 3–8. [Google Scholar]
- Soulage, C.; Perrin, D.; Berenguer, P.; Pequignot, J. Sub-chronic exposure to toluene at 40 ppm alters the monoamine biosynthesis rate in discrete brain areas. Toxicology 2004, 196, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Wada, H. Behavioral approaches to toluene intoxication. Environ. Res. 1993, 62, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Wada, H.; Hosokawa, T.; Saito, K. Repeated toluene exposure and changes of response latency in shock avoidance learning. Neurotoxicol. Teratol. 1988, 10, 387–391. [Google Scholar] [CrossRef]
- Miyake, H. Slow learning in rats due to long-term inhalation of toluene. Neurobehav. Toxicol. Teratol. 1983, 5, 541–548. [Google Scholar]
- Wada, H.; Marushichi, H.; Abe, M.; Kitadate, T.; Kunoh, H.; Iwamoto, T. Effects of toluene administration on delayed matching-to-sample performance in the pigeon. In Neurobehavioral Methods and Effects in Occupational and Environmental Health; Elsevier: Amsterdam, The Netherlands, 1994; pp. 847–856. [Google Scholar]
- Dorman, D.C. An integrative approach to neurotoxicology. Toxicol. Pathol. 2000, 28, 37–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codina, M.; Li, J.; Gutierrez, J.; Kao, J.P.; Du, S.J. Loss of Smyhc1 or Hsp90α1 function results in different effects on myofibril organization in skeletal muscles of zebrafish embryos. PLoS ONE 2010, 5, e8416. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.; Hershberger, W.; Juchau, M. Metabolism of 3, 4-benzpyrene in rainbow trout (Salmo gairdneri). Bull. Environ. Contam. Toxicol. 1974, 12, 481–486. [Google Scholar] [PubMed]
- Payne, J.; Penrose, W. Induction of aryl hydrocarbon (benzo (a) pyrene) hydroxylase in fish by petroleum. Bull. Environ. Contam. Toxicol. 1975, 14, 112–116. [Google Scholar] [CrossRef]
- Thomas, R.; Rice, S. The effect of exposure temperatures on oxygen consumption and opercular breathing rates of pink salmon fry to toluene, naphthalene, and water-soluble fractions of Cook Inlet crude oil and no. 2 fuel oil. In Marine Pollution: Functional Responses; Academic Press, Inc.: Cambridge, MA, USA; New York, NY, USA, 1979. [Google Scholar]
- Berry, W.O.; Brammer, J.; Bee, D.E. Uptake of water-soluble gasoline fractions and their effect on oxygen consumption in aquatic stages of the mosquito (Aedes aegypti (L.)). Environ. Pollut. (1970) 1978, 15, 1–22. [Google Scholar] [CrossRef]
- Smit, H. Some experiments on the oxygen consumption of goldfish (Carassius auratus L.) in relation to swimming speed. Can. J. Zool. 1965, 43, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Spoor, W. A quantitative study of the relationship between the activity and oxygen consumption of the goldfish, and its application to the measurement of respiratory metabolism in fishes. Biol. Bull. 1946, 91, 312–325. [Google Scholar] [CrossRef]
- Kramer, D.L. Dissolved oxygen and fish behavior. Environ. Biol. Fishes 1987, 18, 81–92. [Google Scholar] [CrossRef]
- Katsu-Kimura, Y.; Nakaya, F.; Baba, S.A.; Mogami, Y. Substantial energy expenditure for locomotion in ciliates verified by means of simultaneous measurement of oxygen consumption rate and swimming speed. J. Exp. Biol. 2009, 212, 1819–1824. [Google Scholar] [CrossRef] [Green Version]
- Placzek, J.D.; Boyce, D.A. Orthopaedic Physical Therapy Secrets-E-Book; Elsevier Health Sciences: St. Louis, MO, USA, 2016. [Google Scholar]
- Inoue, M.; Hori, M.; Imai, S.; Berne, R.M. Regulation of Coronary Blood Flow; Springer: Tokyo, Japan, 2013. [Google Scholar]
- Gao, Y. Biology of Vascular Smooth Muscle: Vasoconstriction and Dilatation; Springer Nature: Singapore, 2017. [Google Scholar]
- Zhou, P.; He, A.; Pu, W.T. Regulation of GATA4 transcriptional activity in cardiovascular development and disease. Curr. Top. Dev. Biol. 2012, 100, 143–169. [Google Scholar]
- Tongsong, T.; Wanapirak, C.; Piyamongkol, W.; Sirichotiyakul, S.; Tongprasert, F.; Srisupundit, K.; Luewan, S. Fetal ventricular shortening fraction in hydrops fetalis. Obstet. Gynecol. 2011, 117, 84–91. [Google Scholar] [CrossRef]
- Christian, H.A. Drug treatment of cardiac decompensation. J. Am. Med. Assoc. 1937, 108, 44–46. [Google Scholar] [CrossRef]
- Mei, J.; Zhang, Q.-Y.; Li, Z.; Lin, S.; Gui, J.-F. C1q-like inhibits p53-mediated apoptosis and controls normal hematopoiesis during zebrafish embryogenesis. Dev. Biol. 2008, 319, 273–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickart, M.A.; Klee, E.W.; Nielsen, A.L.; Sivasubbu, S.; Mendenhall, E.M.; Bill, B.R.; Chen, E.; Eckfeldt, C.E.; Knowlton, M.; Robu, M.E. Genome-wide reverse genetics framework to identify novel functions of the vertebrate secretome. PLoS ONE 2006, 1, e104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bass, M. Sudden sniffing death. JAMA 1970, 212, 2075–2079. [Google Scholar] [CrossRef] [PubMed]
- Einav, S.; Amitai, Y.; Reichman, J.; Geber, D. Bradycardia in toluene poisoning. J. Toxicol. Clin. Toxicol. 1997, 35, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, C.F.; Azar, A.; Maxfield, M.E.; Smith Jr, P.E.; Mullin, L.S. Cardiac arrhythmias and aerosol “sniffing”. Arch. Environ. Health Int. J. 1971, 22, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.J.; Samsam, T.E.; Oshiro, W.M.; Bushnell, P.J. Cardiovascular effects of oral toluene exposure in the rat monitored by radiotelemetry. Neurotoxicol. Teratol. 2007, 29, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Taylor, G.J.; Harris, W.S. Glue sniffing causes heart block in mice. Science 1970, 170, 866–868. [Google Scholar] [CrossRef]
- Vidrio, H.; Magos, G.; Lorenzana-Jimenez, M. Electrocardiographic effects of toluene in the anesthetized rat. Arch. Int. Pharmacodyn. Ther. 1986, 279, 121–129. [Google Scholar]
- Ikeda, N.; Takahashi, H.; Umetsu, K.; Suzuki, T. The course of respiration and circulation in ‘toluene-sniffing’. Forensic Sci. Int. 1990, 44, 151–158. [Google Scholar] [CrossRef]
- Yasar, S.; Yildirim, E.; Koklu, M.; Gursoy, E.; Celik, M.; Yuksel, U.C. A case of reversible cardiomyopathy associated with acute toluene exposure. Turk. J. Emerg. Med. 2016, 16, 123–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Wang, Z.; Peng, Y.; Xu, R.; Yan, J.; Xiong, C.; Ma, J.; Zhong, K.; Lu, H. Carboxin can induce cardiotoxicity in zebrafish embryos. Ecotoxicol. Environ. Saf. 2022, 233, 113318. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Ni, T.T.; Hou, J.; Rellinger, E.; Zhong, T.P. Promoter analysis of ventricular myosin heavy chain (vmhc) in zebrafish embryos. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2009, 238, 1760–1767. [Google Scholar]
- Streicher, H.Z.; Gabow, P.A.; Moss, A.H.; Kono, D.; KAEHNY, W.D. Syndromes of toluene sniffing in adults. Ann. Intern. Med. 1981, 94, 758–762. [Google Scholar] [CrossRef] [PubMed]


| Proteins (UniProt) | Grid Box Parameters (x, y, z Coordinates) |
|---|---|
| ATPase GET3 (Q6IQE5) | Center: x = −5.014, y = −5.685, z = 2.564 Dimensions (Å): x = 64.180, y = 73.503, z = 89.405 |
| Calcium-transporting ATPase (A0A286Y8X6) | Center: x = 11.827, y = 14.899, z = 3.753 Dimensions (Å): x = 120.170, y = 138.445, z = 136.836 |
| Cardiac troponin C (Q800V7) | Center: x = 11.827, y = 14.899, z = 3.753 Dimensions (Å): x = 120.170, y = 138.445, z = 136.836 |
| Fast skeletal muscle troponin C (Q918U8) | Center: x = −6.144, y = 1.479, z = 0.281 Dimensions (Å): x = 53.230, y = 36.770, z = 61.960 |
| Myosin XIX (A0A0R4IEQ7) | Center: x = 3.520, y = 0.499, z = −6.061 Dimensions (Å): x= 116.386, y = 76.255, z= 82.202 |
| Phospholipid-transporting ATPase (A0A0R4IC01) | Center: x = 0.368, y = 3.647, z = −0.397 Dimensions (Å): x = 117.422, y = 87.093, z = 112.228 |
| Sodium potassium transporting ATPase subunit alpha (A0A0R4IJ10) | Center: x = −10.402, y = −0.074, z = −5.433 Dimensions (Å): x = 103.253, y = 76.181, z = 109.807 |
| Sodium/potassium-transporting ATPase subunit Beta (Q9DGL3) | Center: x = −7.726, y = 10.487, z = −22.818 Dimensions (Å): x = 69.418, y = 70.062, z= 111.353 |
| Sodium/potassium-transporting ATPase subunit-beta−1-interacting protein 1 (Q6PHL4) | Center: x = 19.534, y = 2.194, z = −12.666 Dimensions (Å): x = 96.393, y = 59.773, z = 85.675 |
| v-type proton ATPase subunit A (A0A286Y8R2) | Center: x = −34.780, y = −3.090, z = −6.351 Dimensions (Å): x = 169.683, y = 84.065, z = 17.886 |
| v-type proton ATPase subunit C (F1QPC9) | Center: x = 1.970, y = −4.212, z = −5.107 Dimensions (Å): x = 92.620, y = 73.399, z = 107.690 |
| v-type proton ATPase subunit H (B0R0V8) | Center: x = −15.095, y = −3.567, z = 3.838 Dimensions (Å): x = 103.359, y = 59.900, z = 88.660 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Young, A.Y.W.; Audira, G.; Saputra, F.; Alos, H.C.; Aventurado, C.A.; Lai, Y.-H.; Vasquez, R.D.; Hsiao, C.-D.; Hung, C.-H. Toxicity Assessment of an Anti-Cancer Drug of p-Toluene Sulfonamide in Zebrafish Larvae Based on Cardiovascular and Locomotion Activities. Biomolecules 2022, 12, 1103. https://doi.org/10.3390/biom12081103
Young AYW, Audira G, Saputra F, Alos HC, Aventurado CA, Lai Y-H, Vasquez RD, Hsiao C-D, Hung C-H. Toxicity Assessment of an Anti-Cancer Drug of p-Toluene Sulfonamide in Zebrafish Larvae Based on Cardiovascular and Locomotion Activities. Biomolecules. 2022; 12(8):1103. https://doi.org/10.3390/biom12081103
Chicago/Turabian StyleYoung, Andrew Yau Wah, Gilbert Audira, Ferry Saputra, Honeymae C. Alos, Charlaine A. Aventurado, Yu-Heng Lai, Ross D. Vasquez, Chung-Der Hsiao, and Chih-Hsin Hung. 2022. "Toxicity Assessment of an Anti-Cancer Drug of p-Toluene Sulfonamide in Zebrafish Larvae Based on Cardiovascular and Locomotion Activities" Biomolecules 12, no. 8: 1103. https://doi.org/10.3390/biom12081103