
Conservation Biology and Reproduction in a Time of Developmental Plasticity



{kind=link}
{kind=link}
Abstract
1. Introduction
2. Conservation and Adaptations
3. Contemporary Evolution and Developmental Plasticity
Plasticity under Extreme Conditions
4. Developmental Plasticity Is a Continuing Process
5. Reproductive Forecasting and Development
6. Wildlife Conservation, Captive Breeding, and Mammalian Sex Ratios
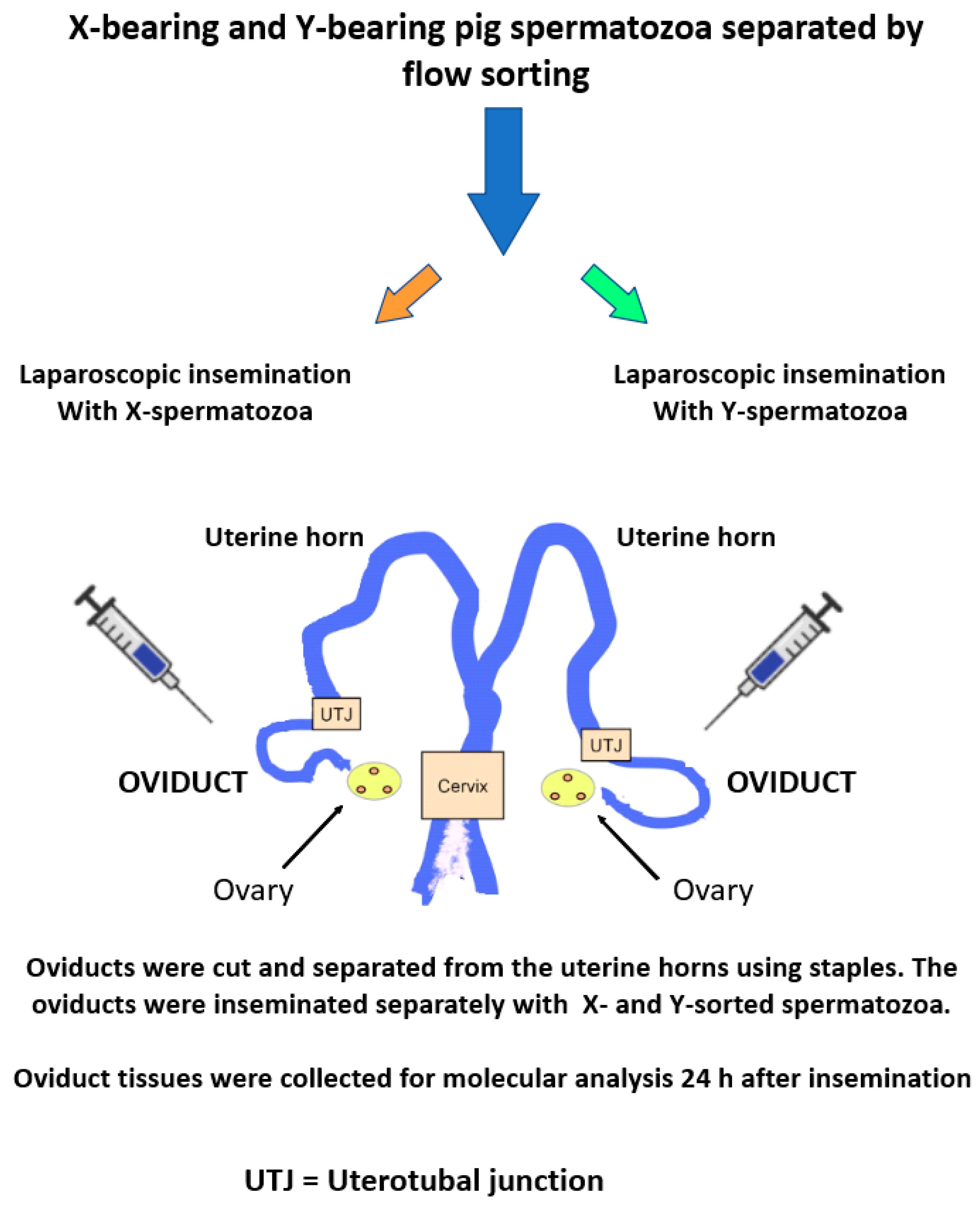
Oviductal Cells Can Discriminate between X- and Y-Chromosome-Bearing Spermatozoa
7. Impacts of Gamete Cryopreservation on Offspring Development and Survival
8. ART and Human Infertility Treatment
9. Stem Cells, Conservation and Ethics
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McFarlane, L.; Wilhelm, D. Non-coding RNAs in mammalian sexual development. Sex. Dev. 2010, 3, 302–316. [Google Scholar] [CrossRef] [PubMed]
- Turner, B.M. Lamarck and the nucleosome: Evolution and environment across 200 years. Front. Life Sci. 2013, 7, 2–11. [Google Scholar] [CrossRef][Green Version]
- Turner, B.M. Nucleosome signalling; An evolving concept. Biochim. Biophys. Acta 2014, 1839, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Bateson, P.; Gluckman, P.; Hanson, M. The biology of developmental plasticity and the Predictive Adaptive Response hypothesis. J. Physiol. 2014, 592, 2357–2368. [Google Scholar] [CrossRef] [PubMed]
- Conde, D.A.; Flesness, N.; Colchero, F.; Jones, O.R.; Scheuerlein, A. An emerging role of zoos to conserve biodiversity. Science 2011, 331, 1390–1391. [Google Scholar] [CrossRef]
- Zeng, Y.; Jiang, Z.; Li, C. Genetic variability in relocated Pere David’s deer (Elaphurus davidianus) populations—Implications to reintroduction program. Conserv. Genet. 2007, 8, 1051–1059. [Google Scholar] [CrossRef]
- Santymire, R.M.; Livieri, T.M.; Branvold-Faber, H.; Marinari, P.E. The black-footed ferret: On the brink of recovery? In Reproductive Sciences in Animal Conservation; Holt, W.V., Brown, J.L., Comizzoli, P., Eds.; Springer: New York, NY, USA, 2014; pp. 119–134. [Google Scholar]
- Howard, J.G.; Lynch, C.; Santymire, R.M.; Marinari, P.E.; Wildt, D.E. Recovery of gene diversity using long-term cryopreserved spermatozoa and artificial insemination in the endangered black-footed ferret: Black-footed ferret gene restoration. Anim. Conserv. 2016, 19, 102–111. [Google Scholar] [CrossRef]
- Farquharson, K.A.; Hogg, C.J.; Grueber, C.E. A meta-analysis of birth-origin effects on reproduction in diverse captive environments. Nat. Commun. 2018, 9, 1055. [Google Scholar] [CrossRef]
- Araki, H.; Cooper, B.; Blouin, M.S. Genetic effects of captive breeding cause a rapid, cumulative fitness decline in the wild. Science 2007, 318, 100–103. [Google Scholar] [CrossRef]
- Yan, H.; Bonasio, R.; Simola, D.F.; Liebig, J.; Berger, S.L.; Reinberg, D. DNA methylation in social insects: How epigenetics can control behavior and longevity. Annu. Rev. Entomol. 2015, 60, 435–452. [Google Scholar] [CrossRef]
- Weiner, S.A.; Toth, A.L. Epigenetics in social insects: A new direction for understanding the evolution of castes. Genet. Res. Int. 2012, 2012, 609810. [Google Scholar] [CrossRef] [PubMed]
- Weiner, S.A.; Galbraith, D.A.; Adams, D.C.; Valenzuela, N.; Noll, F.B.; Grozinger, C.M.; Toth, A.L. A survey of DNA methylation across social insect species, life stages, and castes reveals abundant and caste-associated methylation in a primitively social wasp. Naturwissenschaften 2013, 100, 795–799. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Lamm, M.S.; Rutherford, K.; Black, M.A.; Godwin, J.R.; Gemmell, N.J. Large-scale transcriptome sequencing reveals novel expression patterns for key sex-related genes in a sex-changing fish. Biol. Sex Differ. 2015, 6, 26. [Google Scholar] [CrossRef] [PubMed]
- Todd, E.V.; Ortega-Recalde, O.; Liu, H.; Lamm, M.S.; Rutherford, K.M.; Cross, H.; Black, M.A.; Kardailsky, O.; Graves, J.A.M.; Hore, T.A.; et al. Stress, novel sex genes, and epigenetic reprogramming orchestrate socially controlled sex change. Sci. Adv. 2019, 5, eaaw7006. [Google Scholar] [CrossRef] [PubMed]
- Richard, G.; Le Trionnaire, G.; Danchin, E.; Sentis, A. Epigenetics and insect polyphenism: Mechanisms and climate change impacts. Curr. Opin. Insect Sci. 2019, 35, 138–145. [Google Scholar] [CrossRef]
- Podrabsky, J.E.; Tingaud-Sequeira, A.; Cerda, J. Metabolic dormancy and responses to environmental desiccation in fish embryos. In Dormancy and Resistance in Harsh Environments; Lubzens, E., Cerda, J., Clark, M., Eds.; Topics in Current Genetics; Springer: Heidelberg, Germany, 2010; Volume 21, pp. 203–226. [Google Scholar]
- Hand, S.C.; Moore, D.S.; Patil, Y. Challenges during diapause and anhydrobiosis: Mitochondrial bioenergetics and desiccation tolerance. IUBMB Life 2018, 70, 1251–1259. [Google Scholar] [CrossRef]
- Zajic, D.E.; Nicholson, J.P.; Podrabsky, J.E. No water, no problem: Stage-specific metabolic responses to dehydration stress in annual killifish embryos. J. Exp. Biol. 2020, 223, jeb231985. [Google Scholar] [CrossRef]
- Podrabsky, J.E.; Carpenter, J.F.; Hand, S.C. Survival of water stress by embryos of an annual killifish: Dehydration avoidance and protein structure of the egg envelope. Am. Zool. 1999, 39, 68A. [Google Scholar]
- Tingaud-Sequeira, A.; Zapater, C.; Chauvigne, F.; Otero, D.; Cerda, J. Adaptive plasticity of killifish (Fundulus heteroclitus) embryos: Dehydration-stimulated development and differential aquaporin-3 expression. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R1041–R1052. [Google Scholar] [CrossRef]
- Jorquera, M.A.; Graether, S.P.; Maruyama, F. Editorial: Bioprospecting and biotechnology of extremophiles. Front. Bioeng. Biotechnol. 2019, 7, 204. [Google Scholar] [CrossRef]
- Willoughby, J.R.; Harder, A.M.; Tennessen, J.A.; Scribner, K.T.; Christie, M.R. Rapid genetic adaptation to a novel environment despite a genome-wide reduction in genetic diversity. Mol. Ecol. 2018, 27, 4041–4051. [Google Scholar] [CrossRef] [PubMed]
- Christie, M.R.; Marine, M.L.; French, R.A.; Blouin, M.S. Genetic adaptation to captivity can occur in a single generation. Proc. Natl. Acad. Sci. USA 2012, 109, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.P.; Reznick, D.N.; Kinnison, M.T.; Bryant, M.J.; Weese, D.J.; Rasanen, K.; Millar, N.P.; Hendry, A.P. Adaptive changes in life history and survival following a new guppy introduction. Am. Nat. 2009, 174, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, C.A.; Hendry, A.P.; Kinnison, M.T. Contemporary evolution meets conservation biology in practice. Trends Ecol. Evol. 2003, 18, 94–101. [Google Scholar] [CrossRef]
- Nichelmann, M. Perinatal epigenetic temperature adaptation in avian species: Comparison of turkey and Muscovy duck. J. Therm. Biol. 2004, 29, 613–619. [Google Scholar] [CrossRef]
- Bronson, F.H. Climate change and seasonal reproduction in mammals. Philos. Trans. R. Soc. Ser. B Biol. Sci. 2009, 364, 3331–3340. [Google Scholar] [CrossRef]
- Kearney, B.D.; Byrne, P.G.; Reina, R.D. Larval tolerance to salinity in three species of Australian anuran: An indication of saline specialisation in Litoria aurea. PLoS ONE 2012, 7, e43427. [Google Scholar] [CrossRef]
- Kearney, B.D.; Byrne, P.G.; Reina, R.D. Short- and long- term consequences of developmental saline stress: Impacts on anuran respiration and behaviour. R. Soc. Open Sci. 2016, 3, 150640. [Google Scholar] [CrossRef]
- Richter-Boix, A.; Tejedo, M.; Rezende, E.L. Evolution and plasticity of anuran larval development in response to desiccation. A comparative analysis. Ecol. Evol. 2011, 1, 15–25. [Google Scholar] [CrossRef]
- Duarte, H.; Tejedo, M.; Katzenberger, M.; Marangoni, F.; Baldo, D.; Beltrán, J.F.; Martí, D.A.; Richter-Boix, A.; Gonzalez-Voyer, A. Can amphibians take the heat? Vulnerability to climate warming in subtropical and temperate larval amphibian communities. Glob. Chang. Biol. 2012, 18, 412–421. [Google Scholar] [CrossRef]
- O’Brien, D.M.; Silla, A.J.; Byrne, P.G. Nest site selection in a terrestrial breeding frog: Interrelationships between nest moisture, pH and male advertisement. Anim. Behav. 2020, 169, 57–64. [Google Scholar] [CrossRef]
- Gould, J.; Clulow, J.; Clulow, S. High clutch failure rate due to unpredictable rainfall for an ephemeral pool-breeding frog. Oecologia 2022, 198, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.S.; Ozanne, S.E.; Sinclair, K.D. Effect of the early-life nutritional environment on fecundity and fertility of mammals. Philos. Trans. R. Soc. B 2009, 364, 3419–3427. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Barker, D.J.; Gelow, J.; Thornburg, K.; Osmond, C.; Kajantie, E.; Eriksson, J.G. The early origins of chronic heart failure: Impaired placental growth and initiation of insulin resistance in childhood. Eur. J. Heart Fail. 2010, 12, 819–825. [Google Scholar] [CrossRef]
- Kwong, W.Y.; Wild, A.E.; Roberts, P.; Willis, A.C.; Fleming, T.P. Maternal undernutrition during the preimplantation period of rat development causes blastocyst abnormalities and programming of postnatal hypertension. Development 2000, 127, 4195–4202. [Google Scholar] [CrossRef]
- Pembrey, M.E.; Bygren, L.O.; Kaati, G.; Edvinsson, S.; Northstone, K.; Sjostrom, M.; Golding, J. Sex-specific, male-line transgenerational responses in humans. Eur. J. Hum. Genet. 2006, 14, 159–166. [Google Scholar] [CrossRef]
- Anway, M.D.; Leathers, C.; Skinner, M.K. Endocrine disruptor vinclozolin induced epigenetic transgenerational adult-onset disease. Endocrinology 2006, 147, 5515–5523. [Google Scholar] [CrossRef]
- Anway, M.D.; Memon, M.A.; Uzumcu, M.; Skinner, M.K. Transgenerational effect of the endocrine disruptor vinclozolin on male spermatogenesis. J. Androl. 2006, 27, 868–879. [Google Scholar] [CrossRef]
- Warburton, E.M.; Khokhlova, I.S.; van der Mescht, L.; Downs, C.J.; Dlugosz, E.M.; Krasnov, B.R. Effects of maternal and grandmaternal flea infestation on offspring quality and quantity in a desert rodent: Evidence for parasite-mediated transgenerational phenotypic plasticity. Int. J. Parasitol. 2019, 49, 481–488. [Google Scholar] [CrossRef]
- Clutton-Brock, T.H.; Iason, G.R. Sex ratio variation in mammals. Q. Rev. Biol. 1986, 61, 339–374. [Google Scholar] [CrossRef]
- Graczyk, M.; Cwiertnia, P.; Borowska, A.; Barczak, E.; Szwaczkowski, T. Inbreeding and offspring sex ratio in the pygmy hippopotamus (Cheoropsis liberiensis) population kept in zoological gardens. Folia Biol. 2015, 63, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Zschokke, S. Distorted sex ratio at birth in the captive pygmy hippopotamus, Hexaprotodon liberiensis. J. Mammal. 2002, 83, 674–681. [Google Scholar] [CrossRef]
- Trivers, R.L.; Willard, D.E. Natural-selection of parental ability to vary sex-ratio of offspring. Science 1973, 179, 90–92. [Google Scholar] [CrossRef]
- Saragusty, J.; Hermes, R.; Goeritz, F.; Schmitt, D.L.; Hildebrandt, T.B. Skewed birth sex ratio and premature mortality in elephants. Anim. Reprod. Sci. 2009, 115, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, M.D.; Tedeschi, J.N.; Garcia-Gonzalez, F.; Firman, R.C. Exposure to male-dominated environments during development influences sperm sex ratios at sexual maturity. Evol. Lett. 2019, 3, 392–402. [Google Scholar] [CrossRef]
- Toni, P.; Forsyth, D.M.; Festa-Bianchet, M. Determinants of offspring sex in kangaroos: A test of multiple hypotheses. Behav. Ecol. 2021, 32, 297–305. [Google Scholar] [CrossRef]
- Hagen, R.; Ortmann, S.; Elliger, A.; Arnold, J. Evidence for a male-biased sex ratio in the offspring of a large herbivore: The role of environmental conditions in the sex ratio variation. Ecol. Evol. 2022, 12, e8938. [Google Scholar] [CrossRef]
- Rosenfeld, C.S.; Roberts, R.M. Maternal diet and other factors affecting offspring sex ratio: A review. Biol. Reprod. 2004, 71, 1063–1070. [Google Scholar] [CrossRef]
- Consuegra, S.; Lopez, C.M.R. Epigenetic-induced alterations in sex-ratios in response to climate change: An epigenetic trap? Bioessays 2016, 38, 950–958. [Google Scholar] [CrossRef]
- Rorie, R.W. Effect of timing of artificial insemination on sex ratio. Theriogenology 1999, 52, 1273–1280. [Google Scholar] [CrossRef]
- James, W.H. Further evidence that mammalian sex ratios at birth are partially controlled by parental hormone levels around the time of conception. Hum. Reprod. 2004, 19, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Cameron, E.Z.; Edwards, A.M.; Parsley, L.M. Developmental sexual dimorphism and the evolution of mechanisms for adjustment of sex ratios in mammals. Ann. N. Y. Acad. Sci. 2017, 1389, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Holt, W.V.; Fazeli, A. Do sperm possess a molecular passport? Mechanistic insights into sperm selection in the female reproductive tract. Mol. Hum. Reprod. 2015, 21, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Holt, W.V.; Fazeli, A. Sperm selection in the female mammalian reproductive tract. Focus on the oviduct: Hypotheses, mechanisms, and new opportunities. Theriogenology 2016, 85, 105–112. [Google Scholar] [CrossRef]
- Thurston, L.M.; Holt, W.V.; Fazeli, A. Battle of the Sexes: How the Selection of Spermatozoa in the Female Reproductive Tract Manipulates the Sex Ratio of Offspring; Academic Press: London, UK, 2017; pp. 251–267. [Google Scholar]
- Tollner, T.L.; Venners, S.A.; Hollox, E.J.; Yudin, A.I.; Liu, X.; Tang, G.; Xing, H.; Kays, R.J.; Lau, T.; Overstreet, J.W.; et al. A common mutation in the defensin DEFB126 causes impaired sperm function and subfertility. Sci. Transl. Med. 2011, 3, 92ra65. [Google Scholar] [CrossRef]
- Qu, Y.; Chen, Q.; Guo, S.; Ma, C.; Lu, Y.; Shi, J.; Liu, S.; Zhou, T.; Noda, T.; Qian, J.; et al. Cooperation-based sperm clusters mediate sperm oviduct entry and fertilization. Protein Cell 2021, 12, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Schoeller, S.F.; Holt, W.V.; Keaveny, E.E. Collective dynamics of sperm cells. Philos. Trans. R. Soc. Ser. B Biol. Sci. 2020, 375, 20190384. [Google Scholar] [CrossRef] [PubMed]
- Gadella, B.M.; Harrison, R.A. The capacitating agent bicarbonate induces protein kinase A-dependent changes in phospholipid transbilayer behavior in the sperm plasma membrane. Development 2000, 127, 2407–2420. [Google Scholar] [CrossRef]
- Okamura, N.; Tajima, Y.; Soejima, A.; Masuda, H.; Sugita, Y. Sodium bicarbonate in seminal plasma stimulates the motility of mammalian spermatozoa through direct activation of adenylate cyclase. J. Biol. Chem. 1985, 260, 9699–9705. [Google Scholar] [CrossRef]
- Nishimura, H.; Kim, E.; Nakanishi, T.; Baba, T. Possible function of the adam1a/adam2 fertilin complex in the appearance of ADAM3 on the sperm surface. J. Biol. Chem. 2004, 279, 34957–34962. [Google Scholar] [CrossRef]
- Yamaguchi, R.; Muro, Y.; Isotani, A.; Tokuhiro, K.; Takumi, K.; Adham, I.; Ikawa, M.; Okabe, M. Disruption of ADAM3 impairs the migration of sperm into oviduct in mouse. Biol. Reprod. 2009, 81, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Holtzmann, I.; Wolf, J.P.; Ziyyat, A. Sperm reservoir in mice: Involvement of ADAMs. Gynecol. Obstet. Fertil. 2011, 39, 630–632. [Google Scholar] [CrossRef] [PubMed]
- Machado, S.A.; Sharif, M.; Wang, H.; Bovin, N.; Miller, D.J. Release of porcine sperm from oviduct cells is stimulated by progesterone and requires CatSper. Sci. Rep. 2019, 9, 19546. [Google Scholar] [CrossRef] [PubMed]
- Vieira, L.A.; Diana, A.; Soriano-Ubeda, C.; Matas, C. Selection of boar sperm by reproductive biofluids as chemoattractants. Animals 2021, 11, 53. [Google Scholar] [CrossRef]
- Giojalas, C.L.; Guidobaldi, A.H. Getting to and away from the egg, an interplay between several sperm transport mechanisms and a complex oviduct physiology. Mol. Cell. Endocrinol. 2020, 518, 110954. [Google Scholar] [CrossRef]
- Bahat, A.; Tur-Kaspa, I.; Gakamsky, A.; Giojalas, L.C.; Breitbart, H.; Eisenbach, M. Thermotaxis of mammalian sperm cells: A potential navigation mechanism in the female genital tract. Nat. Med. 2003, 9, 149–150. [Google Scholar] [CrossRef]
- Perez-Cerezales, S.; Ramos-Ibeas, P.; Salvador Acuna, O.; Aviles, M.; Coy, P.; Rizos, D.; Gutierrez-Adan, A. The oviduct: From sperm selection to the epigenetic landscape of the embryo. Biol. Reprod. 2018, 98, 262–276. [Google Scholar] [CrossRef]
- Coy, P.; Lloyd, R.; Romar, R.; Satake, N.; Matas, C.; Gadea, J.; Holt, W.V. Effects of porcine pre-ovulatory oviductal fluid on boar sperm function. Theriogenology 2010, 74, 632–642. [Google Scholar] [CrossRef]
- Coy, P.; Avilés, M. What controls polyspermy in mammals, the oviduct or the oocyte? Biol. Rev. 2010, 85, 593–605. [Google Scholar] [CrossRef]
- Schjenken, J.E.; Robertson, S.A. The female response to seminal fluid. Physiol. Rev. 2020, 100, 1077–1117. [Google Scholar] [CrossRef]
- Bromfield, J.J.; Schjenken, J.E.; Chin, P.Y.; Care, A.S.; Jasper, M.J.; Robertson, S.A. Maternal tract factors contribute to paternal seminal fluid impact on metabolic phenotype in offspring. Proc. Natl. Acad. Sci. USA 2014, 111, 2200–2205. [Google Scholar] [CrossRef] [PubMed]
- Georgiou, A.S.; Sostaric, E.; Wong, C.H.; Snijders, A.P.; Wright, P.C.; Moore, H.D.; Fazeli, A. Gametes alter the oviductal secretory proteome. Mol. Cell. Proteom. 2005, 4, 1785–1796. [Google Scholar] [CrossRef] [PubMed]
- Georgiou, A.S.; Sostaric, E.; Snijders, A.P.L.; Wright, P.C.; Fazeli, A. Cell surface protein trafficking in oviductal cells in response to spermatozoa. In Proceedings of the 7th Siena Meeting from Genome to Proteome: Back to the Future, Siena, Italy, 3–7 September 2006. [Google Scholar]
- Georgiou, A.S.; Snijders, A.P.; Sostaric, E.; Aflatoonian, R.; Vazquez, J.L.; Vazquez, J.M.; Roca, J.; Martinez, E.A.; Wright, P.C.; Fazeli, A. Modulation of the oviductal environment by gametes. J. Proteome Res. 2007, 6, 4656–4666. [Google Scholar] [CrossRef] [PubMed]
- Holt, W.V.; Fazeli, A. Sperm storage in the female reproductive tract. Annu. Rev. Anim. Biosci. 2016, 4, 291–310. [Google Scholar] [CrossRef] [PubMed]
- Almiñana, C.; Caballero, I.; Heath, P.R.; Maleki-Dizaji, S.; Parrilla, I.; Cuello, C.; Gil, M.A.; Vazquez, J.L.; Vazquez, J.M.; Roca, J.; et al. The battle of the sexes starts in the oviduct: Modulation of oviductal transcriptome by X and Y-bearing spermatozoa. BMC Genom. 2014, 15, 293. [Google Scholar] [CrossRef]
- Rath, D.; Barcikowski, S.; de Graaf, S.; Garrels, W.; Grossfeld, R.; Klein, S.; Knabe, W.; Knorr, C.; Kues, W.; Meyer, H.; et al. Sex selection of sperm in farm animals: Status report and developmental prospects. Reproduction 2013, 145, R15–R30. [Google Scholar] [CrossRef]
- Rahman, M.S.; Pang, M.G. New Biological Insights on X and Y Chromosome-Bearing Spermatozoa. Front. Cell Dev. Biol. 2020, 7, 388. [Google Scholar] [CrossRef]
- Comizzoli, P.; Holt, W.V. Recent Progress in Spermatology Contributing to the Knowledge and Conservation of Rare and Endangered Species. Annu. Rev. Anim. Biosci. 2022, 10, 469–490. [Google Scholar] [CrossRef]
- Strand, J.; Thomsen, H.; Jensen, J.B.; Marcussen, C.; Nicolajsen, T.B.; Skriver, M.B.; Sogaard, I.M.; Ezaz, T.; Purup, S.; Callesen, H.; et al. Biobanking in amphibian and reptilian conservation and management: Opportunities and challenges. Conserv. Genet. Resour. 2020, 12, 709–725. [Google Scholar] [CrossRef]
- Breithoff, E.; Harrison, R. From ark to bank: Extinction, proxies and biocapitals in ex-situ biodiversity conservation practices. Int. J. Herit. Stud. 2020, 26, 37–55. [Google Scholar] [CrossRef]
- Martin-Wintle, M.S.; Kersey, D.C.; Wintle, N.J.P.; Aitken-Palmer, C.; Owen, M.A.; Swaisgood, R.R. Comprehensive breeding techniques for the giant panda. In Reproductive Sciences in Animal Conservation, 2nd ed.; Comizzoli, P., Brown, J.L., Holt, W.V., Eds.; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2019; Volume 1200, pp. 275–308. [Google Scholar]
- Santymire, R. Implementing the use of a biobank in the endangered black-footed ferret (Mustela nigripes). Reprod. Fertil. Dev. 2016, 28, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, M.L.; Bradshaw, C.J.A.; Goldsworthy, S.D.; Sunnucks, P. Lower reproductive success in hybrid fur seal males indicates fitness costs to hybridization. Mol. Ecol. 2007, 16, 3187–3197. [Google Scholar] [CrossRef] [PubMed]
- Knipler, M.L.; Dowton, M.; Mikac, K.M. Genome-wide snps detect hybridisation of marsupial gliders (Petaurus breviceps breviceps × Petaurus norfolcensis) in the wild. Genes 2021, 12, 1327. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, E.H.C.; Gomes, A.J.B.; Costa, A.F.; Emin-Lima, R.; Bonvicino, C.R.; Viana, M.C.; Reis, L.M.A.; Vidal, M.D.; Cavalcanti, M.V.G.; Attademo, F.L.N.; et al. Karyotypical confirmation of natural hybridization between two manatee species, Trichechus manatus and Trichechus inunguis. Life 2022, 12, 616. [Google Scholar] [CrossRef] [PubMed]
- Fadakar, D.; Malekian, M.; Hemami, M.R.; Lerp, H.; Rezaei, H.R.; Bärmann, E.V. Repeated hybridization of two closely related gazelle species (Gazella bennettii and Gazella subgutturosa) in central Iran. Ecol. Evol. 2020, 10, 11372–11386. [Google Scholar] [CrossRef]
- Rodger, J.C.; Clulow, J. Resetting the paradigm of reproductive science and conservation. Anim. Reprod. Sci. 2021, 106911. [Google Scholar] [CrossRef]
- Gooley, R.M.; Hogg, C.J.; Fox, S.; Pemberton, D.; Belov, K.; Grueber, C.E. Inbreeding depression in one of the last DFTD-free wild populations of Tasmanian devils. Peerj 2020, 8, e9220. [Google Scholar] [CrossRef]
- Hogg, C.J.; McLennan, E.A.; Wise, P.; Lee, A.V.; Pemberton, D.; Fox, S.; Belov, K.; Grueber, C.E. Preserving the demographic and genetic integrity of a single source population during multiple translocations. Biol. Conserv. 2020, 241, 108318. [Google Scholar] [CrossRef]
- Brandies, P.A.; Wright, B.R.; Hogg, C.J.; Grueber, C.E.; Belov, K. Characterization of reproductive gene diversity in the endangered Tasmanian devil. Mol. Ecol. Resour. 2020, 21, 721–732. [Google Scholar] [CrossRef]
- Farquharson, K.A.; Gooley, R.M.; Fox, S.; Huxtable, S.J.; Belov, K.; Pemberton, D.; Hogg, C.J.; Grueber, C.E. Are any populations ‘safe’? Unexpected reproductive decline in a population of Tasmanian devils free of devil facial tumour disease. Wildl. Res. 2018, 45, 31–37. [Google Scholar] [CrossRef]
- Poo, S.; Bogisich, A.; Mack, M.; Lynn, B.K.; Devan-Song, A. Post-release comparisons of amphibian growth reveal challenges with sperm cryopreservation as a conservation tool. Conserv. Sci. Pract. 2022, 4, e572. [Google Scholar] [CrossRef]
- Poo, S.; Hinkson, K.M. Amphibian conservation using assisted reproductive technologies: Cryopreserved sperm affects offspring morphology, but not behavior, in a toad. Glob. Ecol. Conserv. 2020, 21, e00809. [Google Scholar] [CrossRef]
- Kouba, A.J.; Lloyd, R.E.; Houck, M.L.; Silla, A.J.; Calatayud, N.; Trudeau, V.L.; Clulow, J.; Molinia, F.; Langhorne, C.; Vance, C.; et al. Emerging trends for biobanking amphibian genetic resources: The hope, reality and challenges for the next decade. Biol. Conserv. 2013, 164, 10–21. [Google Scholar] [CrossRef]
- Silla, A.J.; Byrne, P.G. The role of reproductive technologies in amphibian conservation breeding programs. Annu. Rev. Anim. Biosci. 2019, 7, 499–519. [Google Scholar] [CrossRef] [PubMed]
- Pearl, E.; Morrow, S.; Noble, A.; Lerebours, A.; Horb, M.; Guille, M. An optimized method for cryogenic storage of Xenopus sperm to maximise the effectiveness of research using genetically altered frogs. Theriogenology 2017, 92, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Upton, R.; Clulow, S.; Calatayud, N.E.; Colyvas, K.; Seeto, R.G.Y.; Wong, L.A.M.; Mahony, M.J.; Clulow, J. Generation of reproductively mature offspring from the endangered green and golden bell frog Litoria aurea using cryopreserved spermatozoa. Reprod. Fertil. Dev. 2021, 33, 562. [Google Scholar] [CrossRef]
- Nusbaumer, D.; da Cunha, L.M.; Wedekind, C. Sperm cryopreservation reduces offspring growth. Proc. R. Soc. Biol. Sci. Ser. B 2019, 286, 20191644. [Google Scholar] [CrossRef]
- Ciereszko, A.; Dietrich, G.J.; Nynca, J.; Dobosz, S.; Zalewski, T. Cryopreservation of rainbow trout semen using a glucose-methanol extender. Aquaculture 2014, 420, 275–281. [Google Scholar] [CrossRef]
- Fernandez-Diez, C.; Gonzalez-Rojo, S.; Montfort, J.; Le Cam, A.; Bobe, J.; Robles, V.; Perez-Cerezales, S.; Herraez, M.P. Inhibition of zygotic DNA repair: Transcriptome analysis of the offspring in trout (Oncorhynchus mykiss). Reproduction 2015, 149, 101–111. [Google Scholar] [CrossRef]
- Gosalvez, J.; Holt, W.V.; Johnston, S.D. Sperm DNA fragmentation and its role in wildlife conservation. Adv. Exp. Med. Biol. 2014, 753, 357–384. [Google Scholar] [CrossRef]
- Kopeika, J.; Kopeika, E.; Zhang, T.; Rawson, D.M.; Holt, W.V. Effect of DNA repair inhibitor (3-aminobenzamide) on genetic stability of loach (Misgurnus fossilis) embryos derived from cryopreserved sperm. Theriogenology 2004, 61, 1661–1673. [Google Scholar] [CrossRef]
- De Mello, F.; Garcia, J.S.; Godoy, L.C.; Depince, A.; Labbe, C.; Streit, D.P., Jr. The effect of cryoprotectant agents on DNA methylation patterns and progeny development in the spermatozoa of Colossoma macropomum. Gen. Comp. Endocrinol. 2017, 245, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Depince, A.; Gabory, A.; Dziewulska, K.; Le Bail, P.-Y.; Jammes, H.; Labbe, C. DNA methylation stability in fish spermatozoa upon external constraint: Impact of fish hormonal stimulation and sperm cryopreservation. Mol. Reprod. Dev. 2020, 87, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Marinovic, Z.; Scekic, I.; Lujic, J.; Urbanyi, B.; Horvath, A. The effects of cryopreservation and cold storage on sperm subpopulation structure of common carp (Cyprinus carpio L.). Cryobiology 2021, 99, 88–94. [Google Scholar] [CrossRef]
- Wyns, C.; De Geyter, C.; Calhaz-Jorge, C.; Kupka, M.S.; Motrenko, T.; Smeenk, J.; Bergh, C.; Tandler-Schneider, A.; Rugescu, I.A.; Vidakovic, S.; et al. ART in Europe, 2017: Results generated from European registries by ESHRE. Hum. Reprod. Open 2021, 2021, hoab026. [Google Scholar] [CrossRef]
- Schroeder, M.; Badini, G.; Sferruzzi-Perri, A.N.; Albrecht, C. The consequences of assisted reproduction technologies on the offspring health throughout life: A placental contribution. Front. Cell Dev. Biol. 2022, 10, 906240. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.; Ghosh, J.; Coutifaris, C.; Sapienza, C.; Mainigi, M. Epigenetic changes and assisted reproductive technologies. Epigenetics 2020, 15, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Palomares, A.; Rodriguez-Wallberg, K.A. Update on the epigenomic implication of embryo cryopreservation methods applied in assisted reproductive technologies with potential long-term health effects. Front. Cell Dev. Biol. 2022, 10, 881550. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, A.; Mochida, K.; Matoba, S.; Inoue, K.; Hama, D.; Kadota, M.; Hiraiwa, N.; Yoshiki, A.; Ogura, A. Development of assisted reproductive technologies for Mus spretus. Biol. Reprod. 2021, 104, 234–243. [Google Scholar] [CrossRef]
- Kaneko, T. Simple gamete preservation and artificial reproduction of mammals using micro-insemination techniques. Reprod. Med. Biol. 2015, 14, 99–105. [Google Scholar] [CrossRef]
- Mayer, I. The role of reproductive sciences in the preservation and breeding of commercial and threatened teleost fishes. In Reproductive Sciences in Animal Conservation, 2nd ed.; Comizzoli, P., Brown, J.L., Holt, W.V., Eds.; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2019; Volume 1200, pp. 187–224. [Google Scholar]
- Rivers, N.; Daly, J.; Temple-Smith, P. New directions in assisted breeding techniques for fish conservation. Reprod. Fertil. Dev. 2020, 32, 807–821. [Google Scholar] [CrossRef] [PubMed]
- Higaki, S.; Eto, Y.; Kawakami, Y.; Yamaha, E.; Kagawa, N.; Kuwayama, M.; Nagano, M.; Katagiri, S.; Takahashi, Y. Production of fertile zebrafish (Danio rerio) possessing germ cells (gametes) originated from primordial germ cells recovered from vitrified embryos. Reproduction 2010, 139, 733–740. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kawakami, Y.; Goto-Kazeto, R.; Saito, T.; Fujimoto, T.; Higaki, S.; Takahashi, Y.; Arai, K.; Yamaha, E. Generation of germ-line chimera zebrafish using primordial germ cells isolated from cultured blastomeres and cryopreserved embryoids. Int. J. Dev. Biol. 2010, 54, 1493–1501. [Google Scholar] [CrossRef]
- Smith, D.L. Transplantation of the nuclei of primordial germ cells into enucleated eggs of Rana pipiens. Proc. Natl. Acad. Sci. USA 1965, 54, 101–107. [Google Scholar] [CrossRef]
- Lesimple, M.; Dournon, C.; Labrousse, M.; Houillon, C. Production of fertile salamanders by transfer of germ cell nuclei into eggs. Development 1987, 100, 471–477. [Google Scholar] [CrossRef]
- Porras-Gomez, T.J.; Villagran-SantaCruz, M.; Moreno-Mendoza, N. Biology of primordial germ cells in vertebrates with emphasis in urodeles amphibians. Mol. Reprod. Dev. 2021, 88, 773–792. [Google Scholar] [CrossRef] [PubMed]
- Lacerda, S.; Costa, G.; Campos-Junior, P.; Segatelli, T.; Yazawa, R.; Takeuchi, Y.; Morita, T.; Yoshizaki, G.; França, L. Germ cell transplantation as a potential biotechnological approach to fish reproduction. Fish Physiol. Biochem. 2013, 39, 3–11. [Google Scholar] [CrossRef]
- Franek, R.; Marinovic, Z.; Lujic, J.; Urbanyi, B.; Fucikova, M.; Kaspar, V.; Psenicka, M.; Horvath, A. Cryopreservation and transplantation of common carp spermatogonia. PLoS ONE 2019, 14, e0205481. [Google Scholar] [CrossRef]
- Tran, K.T.D.; Valli-Pulaski, H.; Colvin, A.; Orwig, K.E. Male fertility preservation and restoration strategies for patients undergoing gonadotoxic therapies. Biol. Reprod. 2022, 107, 382–405. [Google Scholar] [CrossRef]
- Praxedes, É.A.; Borges, A.A.; Santos, M.V.O.; Pereira, A.F. Use of somatic cell banks in the conservation of wild felids. Zoo Biol. 2018, 37, 258–263. [Google Scholar] [CrossRef]
- Wu, X.; Goodyear, S.M.; Abramowitz, L.K.; Bartolomei, M.S.; Tobias, J.W.; Avarbock, M.R.; Brinster, R.L. Fertile offspring derived from mouse spermatogonial stem cells cryopreserved for more than 14 years. Hum. Reprod. 2012, 27, 1249–1259. [Google Scholar] [CrossRef] [PubMed]
- Valdivia, M.; Bravo, Z.; Reyes, J.; Gonzales, G. Rescue and conservation of male adult alpacas (Vicugna pacos) based on spermatogonial stem cell biotechnology using atomized Black Maca as a supplement of cryopreservation medium. Front. Vet. Sci. 2021, 8, 597964. [Google Scholar] [CrossRef] [PubMed]
- Shetty, G.; Mitchell, J.M.; Lam, T.N.A.; Phan, T.T.; Zhang, J.; Tailor, R.C.; Peters, K.A.; Penedo, M.C.; Hanna, C.B.; Clark, A.T.; et al. Postpubertal spermatogonial stem cell transplantation restores functional sperm production in rhesus monkeys irradiated before and after puberty. Andrology 2021, 9, 1603–1616. [Google Scholar] [CrossRef] [PubMed]
- Andrae, C.S.; Oliveira, E.C.S.; Ferraz, M.A.M.M.; Nagashima, J.B. Cryopreservation of grey wolf (Canis lupus) testicular tissue. Cryobiology 2021, 100, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Tasai, M.; Takeda, K.; Nirasawa, K.; Tagami, T. Production of functional gametes from cryopreserved primordial germ cells of the Japanese Quail. J. Reprod. Dev. 2013, 59, 580–587. [Google Scholar] [CrossRef]
- Liu, J.; Cheng, K.M.; Silversides, F.G. Recovery of fertility from adult ovarian tissue transplanted into week-old Japanese quail chicks. Reprod. Fertil. Dev. 2015, 27, 281–284. [Google Scholar] [CrossRef]
- Pimentel, M.M.L.; Santos, F.A.D.; Macêdo, L.B.D.; Brito, P.D.D.; Lima, G.L.; Barreto Junior, R.A.; Bezerra, M.B. Rescue of caprine fetal ovaries, vitrification and follicular development after xenotransplantation in two immunodeficient mice models. Anim. Reprod. 2020, 17, e20190115. [Google Scholar] [CrossRef]
- Mastromonaco, G.F.; Gonzalez-Grajales, L.A.; Filice, M.; Comizzoli, P. Somatic cells, stem cells, and induced pluripotent stem cells: How do they now contribute to conservation? In Reproductive Sciences in Animal Conservation: Progress and Prospects; Holt, W.V., Brown, J.L., Comizzoli, P., Eds.; Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 753, pp. 385–427. [Google Scholar]
- Wiedemann, C.; Hribal, R.; Ringleb, J.; Bertelsen, M.F.; Rasmusen, K.; Andersen, C.Y.; Kristensen, S.G.; Jewgenow, K. Preservation of primordial follicles from lions by slow freezing and xenotransplantation of ovarian cortex into an immunodeficient mouse. Reprod. Domest. Anim. 2012, 47, 300–304. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Stanton, M.M.; Tzatzalos, E.; Donne, M.; Kolundzic, N.; Helgason, I.; Ilic, D. Prospects for the use of induced pluripotent stem cells in animal conservation and environmental protection. Stem Cells Transl. Med. 2019, 8, 7–13. [Google Scholar] [CrossRef]
- Ben-Nun, I.F.; Montague, S.C.; Houck, M.L.; Tran, H.T.; Garitaonandia, I.; Leonardo, T.R.; Wang, Y.C.; Charter, S.J.; Laurent, L.C.; Ryder, O.A.; et al. Induced pluripotent stem cells from highly endangered species. Nat. Methods 2011, 8, 829–831. [Google Scholar] [CrossRef] [PubMed]
- Dicks, N.; Bordignon, V.; Mastromonaco, G.F. Chapter 11—Induced pluripotent stem cells in species conservation: Advantages, applications, and the road ahead. In iPSCs from Diverse Species; Birbrair, A., Ed.; Academic Press: London, UK, 2021; pp. 221–245. [Google Scholar]
- Ge, W.; Cheng, S.-F.; Dyce, P.W.; De Felici, M.; Shen, W. Skin-derived stem cells as a source of primordial germ cell- and oocyte-like cells. Cell Death Dis. 2016, 7, e2471. [Google Scholar] [CrossRef] [PubMed]
- Boroda, A.V. Marine mammal cell cultures: To obtain, to apply, and to preserve. Mar. Environ. Res. 2017, 129, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Linher, K.; Dyce, P.; Li, J.L. Primordial germ cell-like cells differentiated in vitro from skin-derived stem cells. PLoS ONE 2009, 4, e8263. [Google Scholar] [CrossRef] [PubMed]
- Dyce, P.W.; Shen, W.; Huynh, E.; Shao, H.; Villagomez, D.A.F.; Kidder, G.M.; King, W.A.; Li, J.L. Analysis of oocyte-like cells differentiated from porcine fetal skin-derived stem cells. Stem Cells Dev. 2011, 20, 809–819. [Google Scholar] [CrossRef]
- Topping, N.E.; Valenzuela, N. Turtle nest-site choice, anthropogenic challenges, and evolutionary potential for adaptation. Front. Ecol. Evol. 2021, 9, 808621. [Google Scholar] [CrossRef]
- Holleley, C.E.; O’Meally, D.; Sarre, S.D.; Graves, J.A.M.; Ezaz, T.; Matsubara, K.; Azad, B.; Zhang, X.W.; Georges, A. Sex reversal triggers the rapid transition from genetic to temperature-dependent sex. Nature 2015, 523, 79–82. [Google Scholar] [CrossRef]
- Senigaglia, V.; Christiansen, F.; Bejder, L.; Sprogis, K.R.; Cantor, M. Human food provisioning impacts the social environment, home range and fitness of a marine top predator. Anim. Behav. 2022, 187, 291–304. [Google Scholar] [CrossRef]
- Simmet, K.; Wolf, E.; Zakhartchenko, V. Manipulating the epigenome in nuclear transfer cloning: Where, when and how. Int. J. Mol. Sci. 2021, 22, 236. [Google Scholar] [CrossRef]
- Hill, J.R. Incidence of abnormal offspring from cloning and other assisted reproductive technologies. Annu. Rev. Anim. Biosci. 2014, 2, 307–321. [Google Scholar] [CrossRef]
- Loi, P.; Saragusty, J.; Ptak, G. Cloning the mammoth: A complicated task or just a dream? In Reproductive Sciences in Animal Conservation: Progress and Prospects; Holt, W.V., Brown, J.L., Comizzoli, P., Eds.; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2014; Volume 753, pp. 489–502. [Google Scholar]
- Minteer, B.A. The Fall of the Wild: Extinction, De-Extinction, and the Ethics of Conservation; Columbia University Press: New York, NY, USA, 2018. [Google Scholar]
- Jans, V.; Dondorp, W.; Bonduelle, M.; de Die, C.; Mertes, H.; Pennings, G.; de Wert, G. Follow-up in the field of reproductive medicine: An ethical exploration. Reprod. BioMed. Online 2020, 41, 1144–1150. [Google Scholar] [CrossRef] [PubMed]
- Sandler, R.L.; Moses, L.; Wisely, S.M. An ethical analysis of cloning for genetic rescue: Case study of the black-footed ferret. Biol. Conserv. 2021, 257, 109118. [Google Scholar] [CrossRef]
- Landi, M.; Everitt, J.; Berridge, B. Bioethical, reproducibility, and translational challenges of animal models. ILAR J. 2021, 62, 60–65. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holt, W.V.; Comizzoli, P. Conservation Biology and Reproduction in a Time of Developmental Plasticity. Biomolecules 2022, 12, 1297. https://doi.org/10.3390/biom12091297
Holt WV, Comizzoli P. Conservation Biology and Reproduction in a Time of Developmental Plasticity. Biomolecules. 2022; 12(9):1297. https://doi.org/10.3390/biom12091297
Chicago/Turabian StyleHolt, William V., and Pierre Comizzoli. 2022. "Conservation Biology and Reproduction in a Time of Developmental Plasticity" Biomolecules 12, no. 9: 1297. https://doi.org/10.3390/biom12091297
APA StyleHolt, W. V., & Comizzoli, P. (2022). Conservation Biology and Reproduction in a Time of Developmental Plasticity. Biomolecules, 12(9), 1297. https://doi.org/10.3390/biom12091297