
Machine Learning Technology for EEG-Forecast of the Blood–Brain Barrier Leakage and the Activation of the Brain’s Drainage System during Isoflurane Anesthesia

 , , , , ,
, , , , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. EEG and EMG Recording
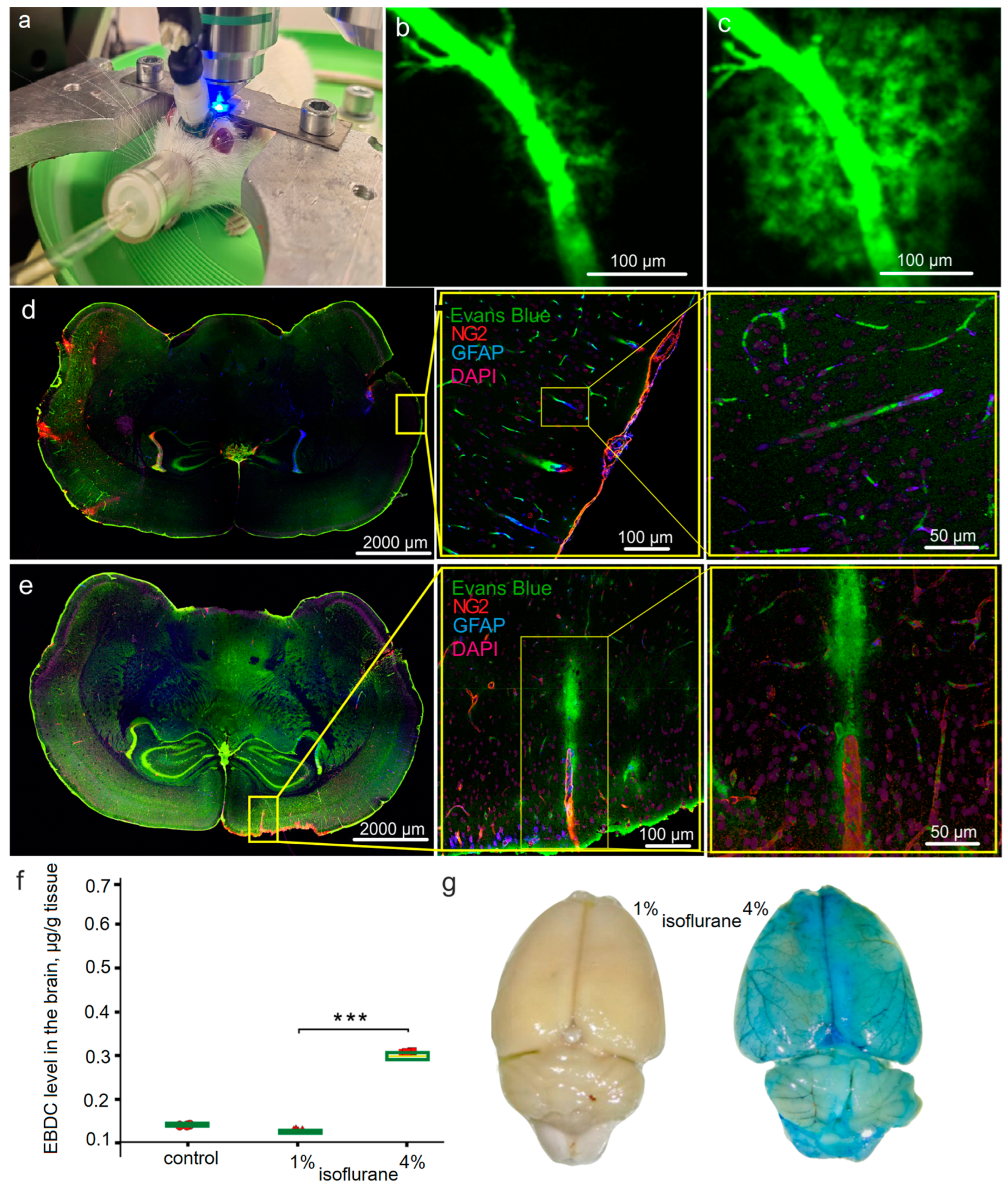
2.3. Optical In Vivo and Ex Vivo Analysis of OBBB
2.4. Spectrofluorometric Assay of EBD Extravasation
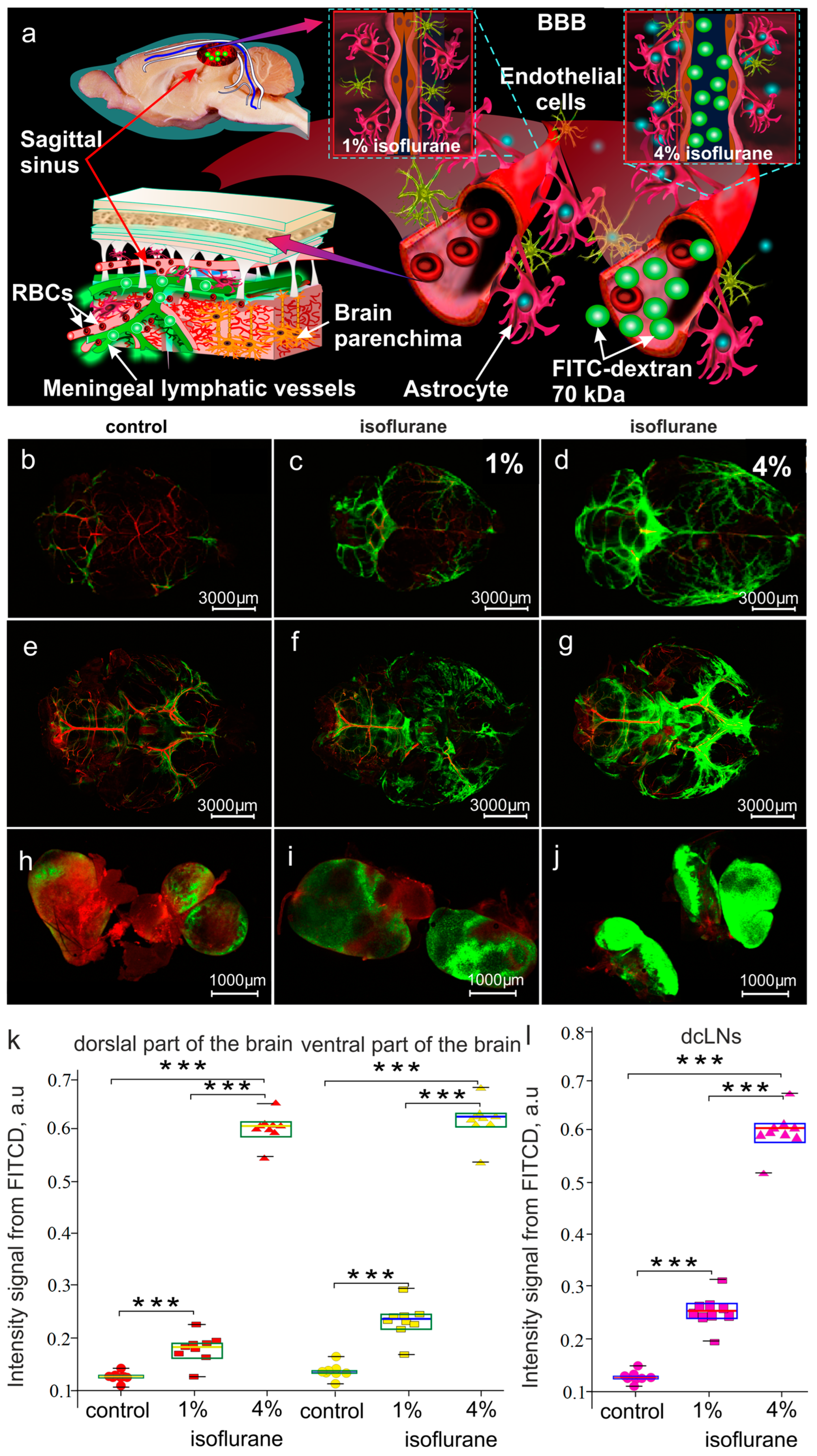
2.5. Optical Monitoring of BDS Activity
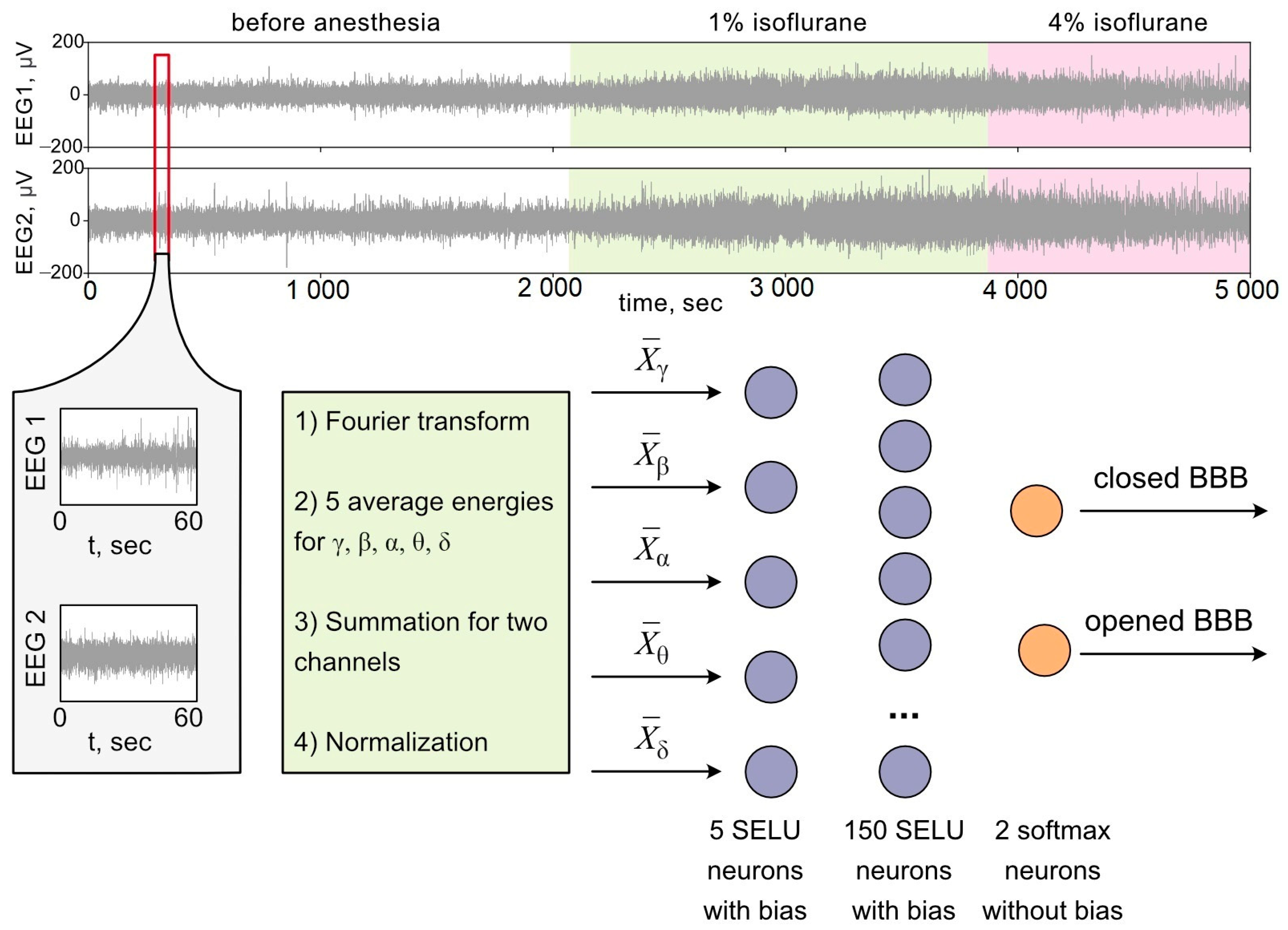
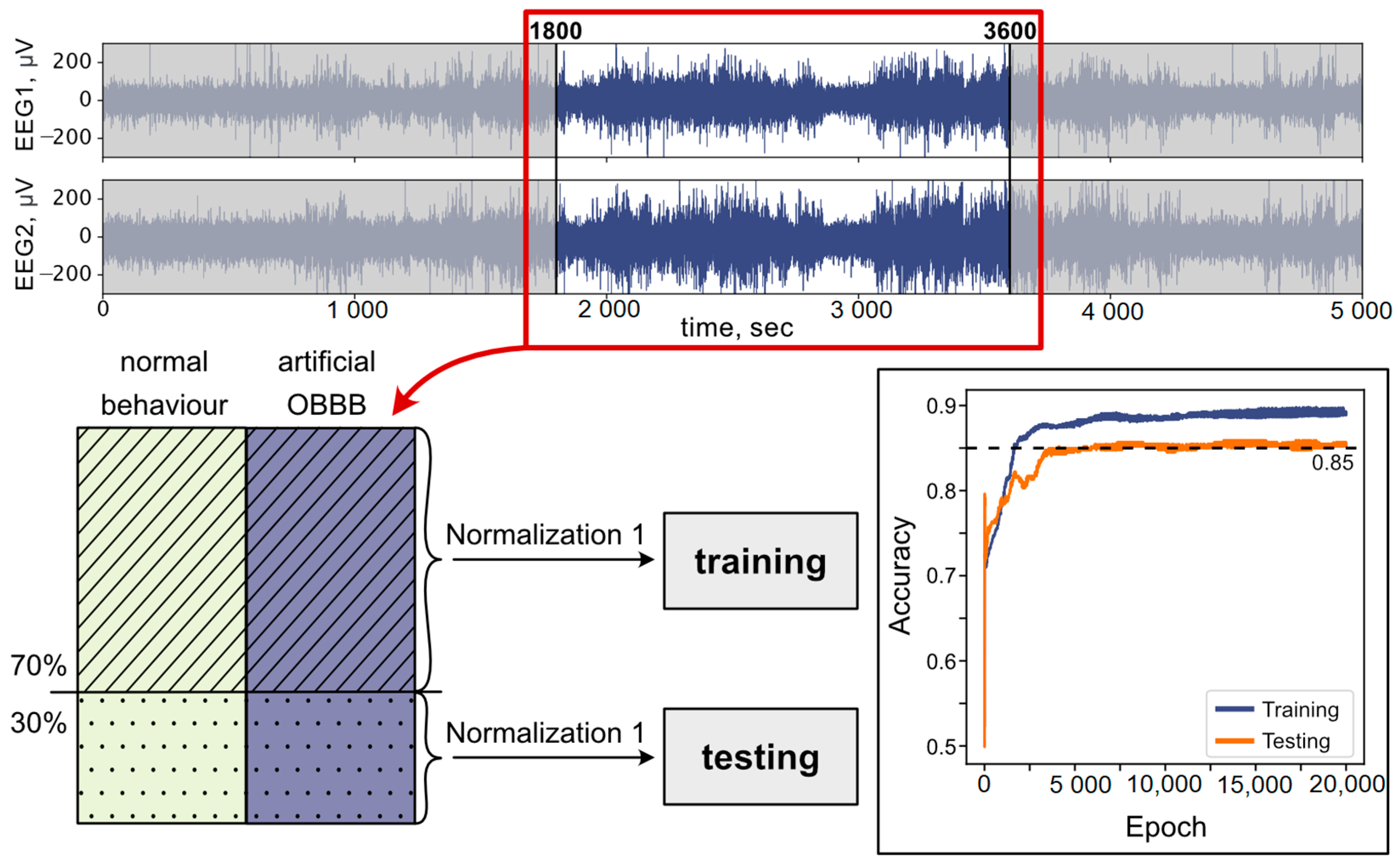
2.6. Artificial Neural Network
2.7. Spectral Analysis
2.8. Statistical Analysis
3. Results
3.1. Effects of Different Doses of Isoflurane Anesthesia on BBB Integrity and BDS Functions
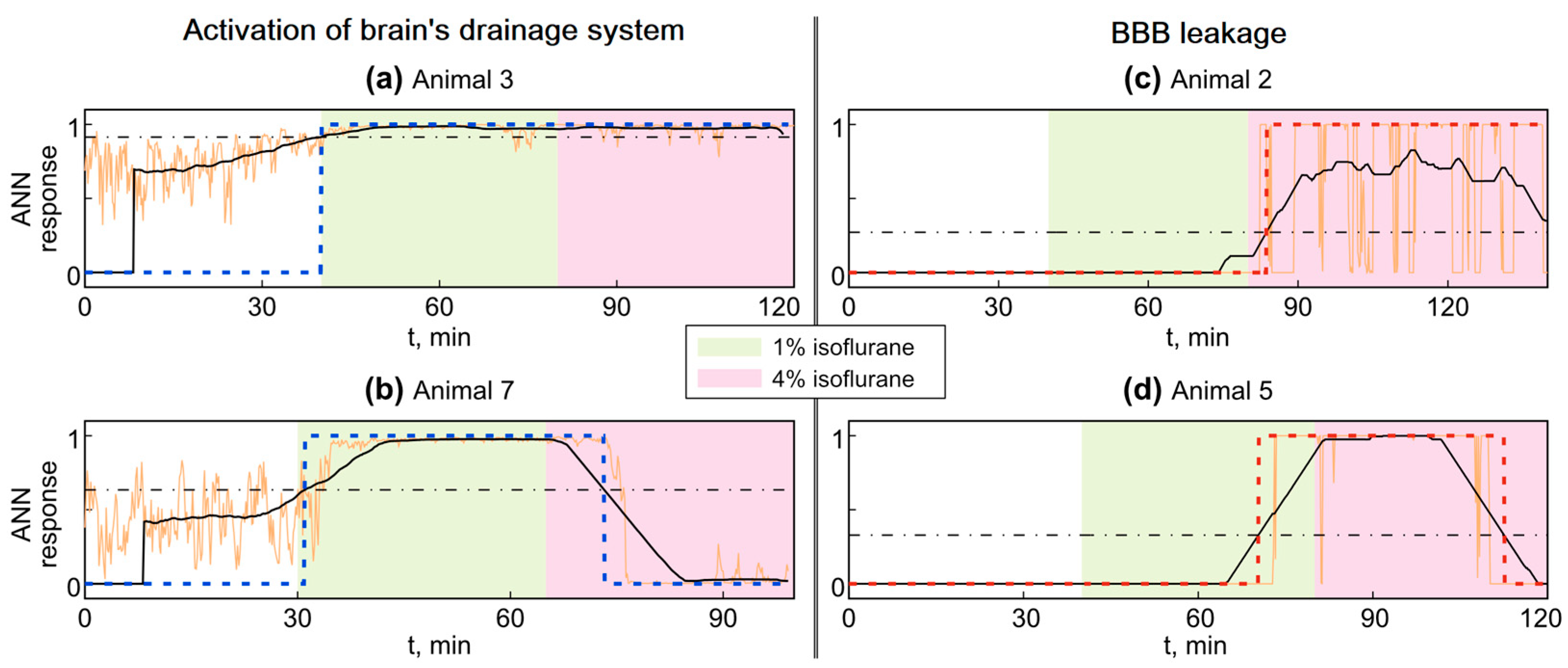
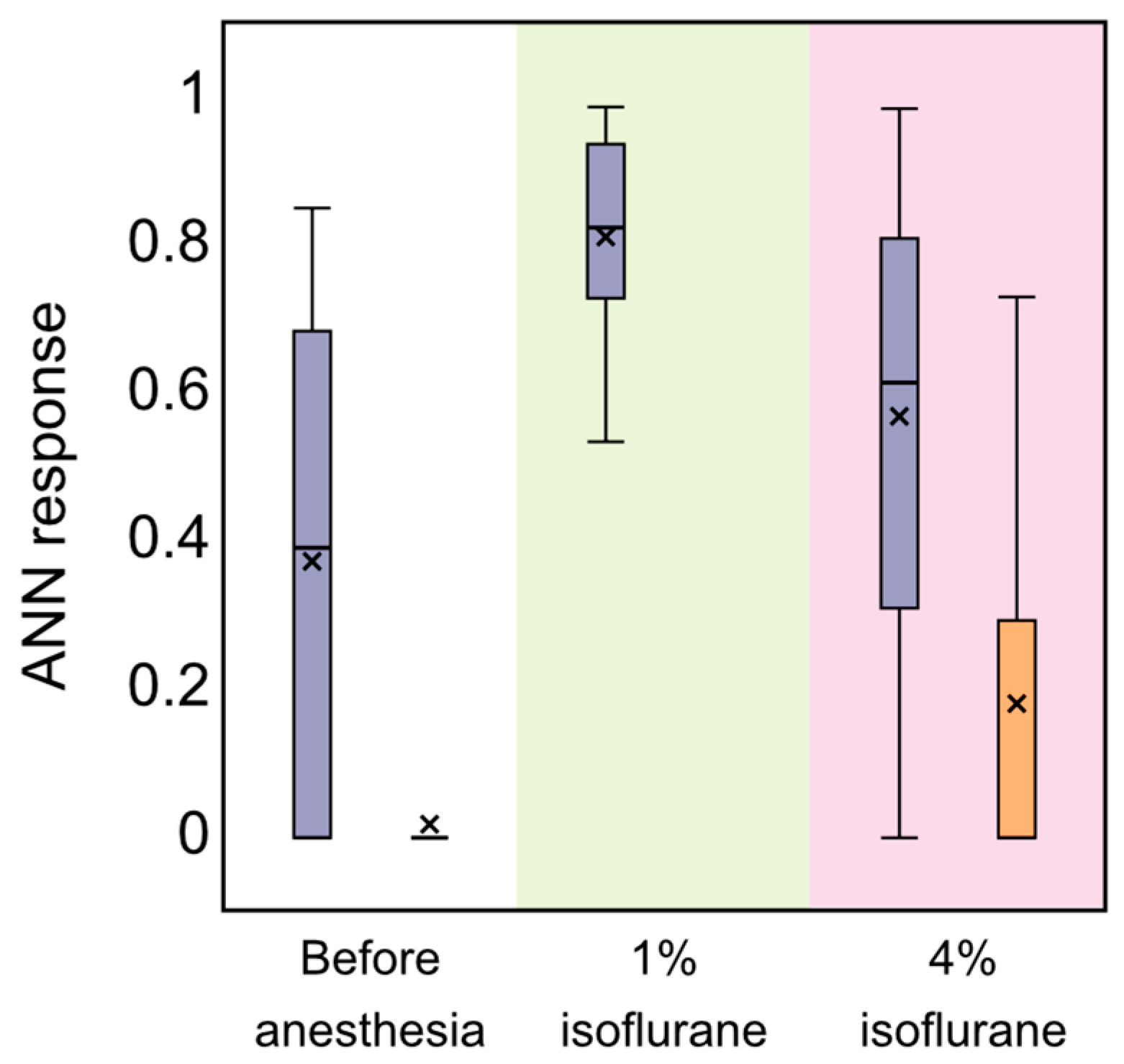
3.2. Analysis of OBBB with ANN
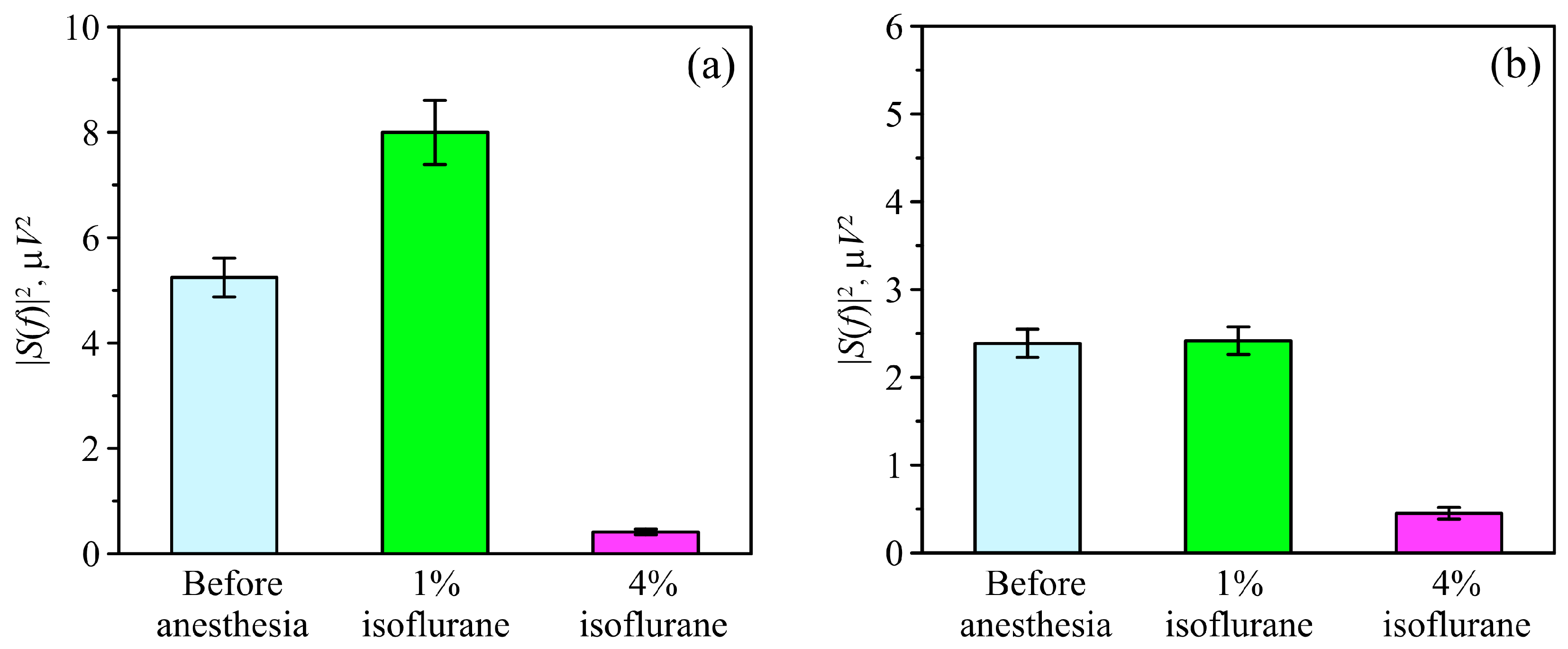
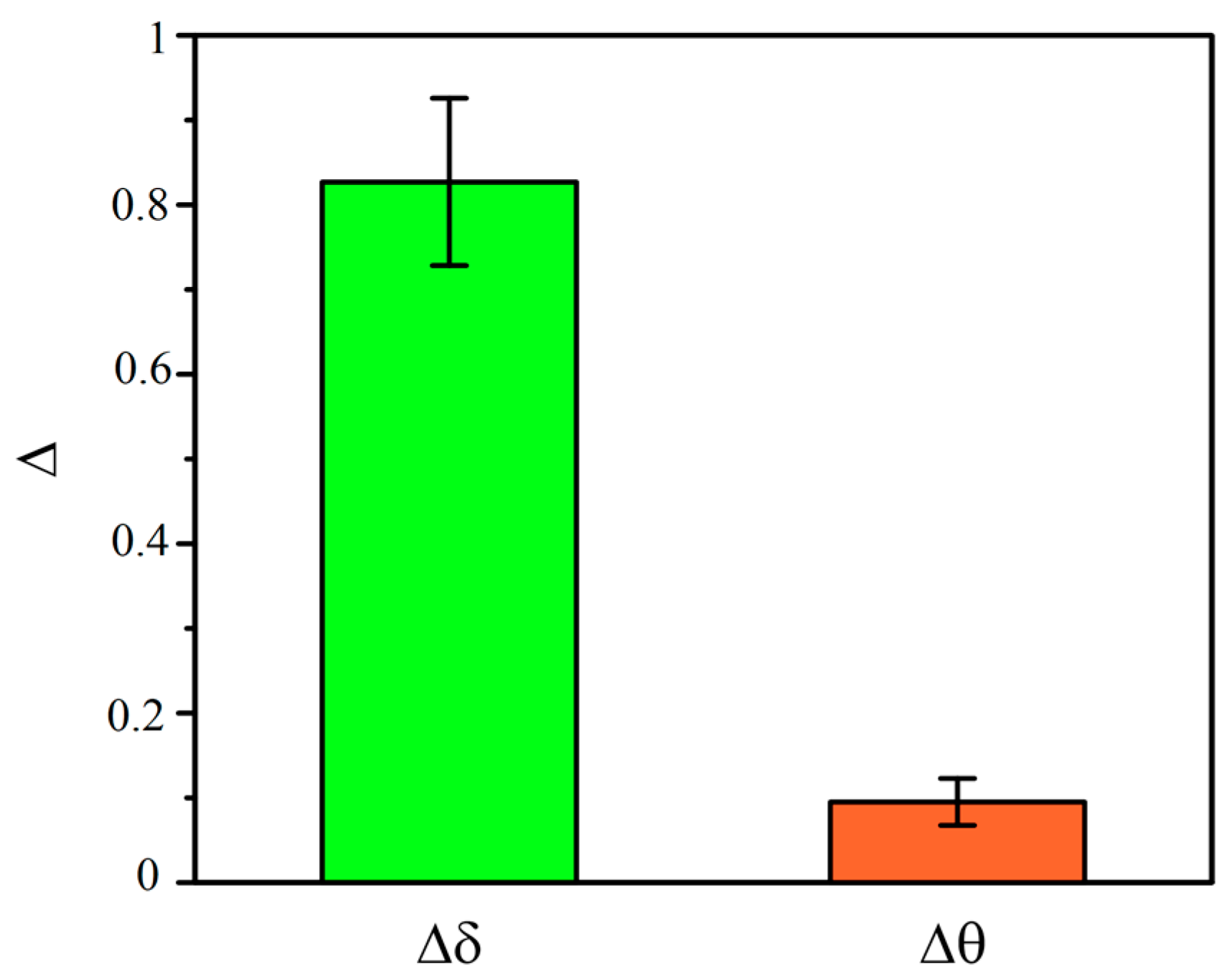
3.3. Power Spectral Analysis of OBBB
4. Discussion
5. Conclusions
6. Suggestions for Future Studies
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, X.; Chen, X. The crosstalk between the blood-brain barrier dysfunction and neuroinflammation after general anaesthesia. Curr. Issues Mol. Biol. 2022, 44, 5700–5717. [Google Scholar] [CrossRef]
- Iqbal, F.; Thompson, A.J.; Riaz, S.; Pehar, M.; Rice, T.; Syed, N.I. Anesthetics: From modes of action to unconsciousness and neurotoxicity. J. Neurophysiol. 2019, 2, 760–787. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Gu, C.; Mandeville, E.T.; Dong, Y.; Esposito, E.; Zhang, Y.; Yang, G.; Shen, Y.; Fu, X.; Lo, E.H. Anesthesia and surgery impair blood-brain barrier and cognitive function in mice. Front. Immunol. 2017, 8, 902. [Google Scholar] [CrossRef] [PubMed]
- Tétrault, S.; Chever, O.; Sik, A.; Amzica, F. Opening of the blood-brain barrier during isoflurane anaesthesia. Eur. J. Neurosci. 2008, 28, 1330–1341. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Ye, X.; Xia, R.; Ran, R.; Wu, Y.; Chen, Q.; Li, H.; Huang, S.; Shu, A.; et al. Anaesthesia-related mortality within 24 h following 9,391,669 anaesthetics in 10 cities in Hubei Province, China: A serial cross-sectional study. Lancet Reg. Health West. Pac. 2023, 37, 100787. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Tellez, N.; Pehar, M.; Visser, F.; Casas-Ortiz, A.; Rice, T.; Syed, N.I. Sevoflurane Exposure in Neonates Perturbs the Expression Patterns of Specific Genes That May Underly the Observed Learning and Memory Deficits. Int. J. Mol. Sci. 2023, 10, 8696. [Google Scholar] [CrossRef]
- Shen, X.; Liu, Y.; Xu, S.; Zhao, Q.; Guo, X.; Shen, R.; Wang, F. Early life exposure to sevoflurane impairs adulthood spatial memory in the rat. Neurotoxicology 2013, 39, 45–56. [Google Scholar] [CrossRef]
- Vutskits, L.; Xie, Z. Lasting impact of general anaesthesia on the brain: Mechanisms and relevance. Nat. Rev. Neurosci. 2016, 17, 705–717. [Google Scholar] [CrossRef]
- Fodale, V.; Tripodi, V.F.; Penna, O.; Fama, F.; Squadrito, F.; Mondello, E.; David, A. An update on anesthetics and impact on the brain. Expert. Opin. Drug Saf. 2017, 16, 997–1008. [Google Scholar] [CrossRef]
- Liu, X.; Ji, J.; Zhao, G.Q. General anesthesia affecting on developing brain: Evidence from animal to clinical research. J. Anesth. 2020, 34, 765–772. [Google Scholar] [CrossRef]
- Zhao, X.; Hao, L.J.; Zhang, Y.T.; Zhang, Y.; Zhang, C. Influence of general anesthetic exposure in developing brain on cognition and the underlying mechanisms. Sheng Li Xue Bao 2019, 71, 749–759. [Google Scholar]
- Fodale, V.; Ritchie, K.; Rasmussen, L.S.; Mandal, P.K. Anesthetics and Alzheimer’s disease: Background and research. J. Alzheimer’s Dis. 2010, 22 (Suppl. S3), 1–3. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Kang, Z.; Li, S.; Zeng, J.; Zhao, J. Sarm1 is Essential for Anesthesia-Induced Neuroinflammation and Cognitive Impairment in Aged Mice. Cell. Mol. Neurobiol. 2022, 42, 1465–1476. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, X.; Li, S.; Li, W.; Teng, Y.; Zhou, Y.; Xiong, H. Carnosol alleviates sevoflurane-induced cognitive dysfunction by mediating NF-kappaB pathway in aged rats. Drug Dev. Res. 2022, 83, 1342–1350. [Google Scholar] [CrossRef]
- Peng, L.; Liu, S.; Xu, J.; Xie, W.; Fang, X.; Xia, T.; Gu, X. Metformin alleviates prolonged isoflurane inhalation induced cognitive decline via reducing neuroinflammation in adult mice. Int. Immunopharmacol. 2022, 109, 108903. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.; Neese, O.; Bieber, D.; Lewis, K.; Ahmadi, L.; Parsons, D.; Canfield, S. The Effects of Propofol on a Human In Vitro Blood-Brain Barrier Model. Front. Cell. Neurosci. 2022, 16, 835649. [Google Scholar] [CrossRef]
- Zhang, Q.; Zheng, M.; Betancourt, C.; Liu, L.; Sitikov, A.; Sladojevic, N.; Zhao, Q.; Zhang, J.; Liao, J.; Wu, R.J.O.M. Increase in Blood-Brain Barrier (BBB) Permeability Is Regulated by MMP3 via the ERK Signaling Pathway. Oxidative Med. Cell. Longev. 2021, 2021, 6655122. [Google Scholar] [CrossRef]
- Spieth, L.; Berghoff, S.A.; Stumpf, S.K.; Winchenbach, J.; Michaelis, T.; Watanabe, T.; Gerndt, N.; Düking, T.; Hofer, S.; Ruhwedel, T. Anesthesia triggers drug delivery to experimental glioma in mice by hijacking caveolar transport. Neuro-Oncol. Adv. 2021, 3, vdab140. [Google Scholar] [CrossRef] [PubMed]
- Zlokovic, B.V. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Zhao, Z.; Montagne, A.; Nelson, A.R.; Zlokovic, B.V. Blood-brain barrier: From physiology to disease and back. Physiol Rev. 2019, 99, 21–78. [Google Scholar] [CrossRef]
- Acharya, N.K.; Goldwaser, E.L.; Forsberg, M.M. Sevoflurane and Isoflurane induce structural changes in brain vascular endothelial cells and increase blood-brain barrier permeability: Possible link to postoperative delirium and cognitive decline. Brain Res. 2015, 1620, 29–41. [Google Scholar] [CrossRef]
- Wu, X.; Lu, Y.; Dong, Y.; Zhang, G.; Zhang, Y.; Xu, Z. The inhalation anesthetic isoflurane increases levels of proinflammatory TNF-alpha, IL-6, and IL-1beta. Neurobiol. Aging 2012, 33, 1364–1378. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Dong, Y.; Xu, Z.; Wang, H.; Miao, C.; Soriano, S.G. Selective anesthesia-induced neuroinflammation in developing mouse brain and cognitive impairment. Anesthesiology 2013, 118, 502–515. [Google Scholar] [CrossRef]
- Tao, G.; Zhang, J.; Zhang, L.; Dong, Y.; Yu, B.; Crosby, G. Sevoflurane induces tau phosphorylation and glycogen synthase kinase 3beta activation in young mice. Anesthesiology 2014, 121, 510–527. [Google Scholar] [CrossRef]
- Rizk, A.; Plitman, E.; Senthil, P.; Venkatraghavan, L.; Chowdhury, T. Effects of Anesthetic Agents on Blood Brain Barrier Integrity: A Systematic Review. Can. J. Neurol. Sci. 2023, 50, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Semyachkina-Glushkovskaya, O.V.; Karavaev, A.S.; Prokhorov, M.D.; Runnova, A.E.; Borovkova, E.I.; Ishbulatov, Y.M.; Hramkov, A.N.; Kulminskiy, D.D.; Semenova, N.I.; Sergeev, K.S.; et al. EEG biomarkers of activation of the lymphatic drainage system of the brain during sleep and opening of the blood-brain barrier. Comput. Struct. Biotechnol. J. 2023, 21, 758–768. [Google Scholar] [CrossRef]
- Hanrahan, G. Artificial Neural Networks in Biological and Environmental Analysis; CRC Press: Boca Raton, FL, USA, 2011; p. 182. [Google Scholar]
- Sadoughi, F.; Ghaderzadeh, M. A hybrid particle swarm and neural network approach for detection of prostate cancer from benign hyperplasia of prostate. Stud. Health Technol. Inf. 2014, 205, 481–485. [Google Scholar]
- Ghaderzadeh, M. Clinical decision support system for early detection of prostate cancer from benign hyperplasia of prostate. Stud. Health Technol. Inform. 2013, 192, 928. [Google Scholar] [PubMed]
- Gheisari, M.; Garavand, A.; Salehnasab, C.; Behmanesh, A.; Aslani, N.; Zadeh, A.H.; Ghaderzadeh, M. Efficient Model for Coronary Artery Disease Diagnosis: A Comparative Study of Several Machine Learning Algorithms. J. Healthc. Eng. 2022, 2022, 5359540. [Google Scholar] [CrossRef]
- Livingstone, D.J. (Ed.) Artificial Neural Networks: Methods and Applications; Humana Press: Totowa, NJ, USA, 2008; p. 202. [Google Scholar]
- Albus, U. Guide for the Care and Use of Laboratory Animals; The National Academies Press: Washington, DC, USA, 2012; p. 246. [Google Scholar]
- Villette, V.; Chavarha, M.; Dimov, I.K.; Dradley, J.; Pradhan, L. Ultrafast two-photon imaging of a high-gain voltage indicator in awake behaving mice. Cell 2019, 179, 1590–1608. [Google Scholar] [CrossRef]
- Koletar, M.M.; Dorr, A.; Brown, M.E.; McLaurin, J.; Stefanovec, B. Refinement of a chronic cranial window implant in the rat for longitudinal in vivo two—Photon fluorescence microscopy of neurovascular function. Sci. Rep. 2019, 9, 5499. [Google Scholar] [CrossRef]
- Wang, H.-L.; Lai, T.W. Optimization of Evans blue quantitation in limited rat tissue samples. Sci. Rep. 2014, 4, 6588. [Google Scholar] [CrossRef] [PubMed]
- Devos, S.L.; Miller, T.M. Direct intraventricular delivery of drugs to the rodent central nervous system. J. Vis. Exp. 2013, 12, e50326. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2019, 9, 676–682. [Google Scholar] [CrossRef]
- Satapathy, S.K.; Loganathan, D. Multimodal Multiclass Machine Learning Model for Automated Sleep Staging Based on Time Series Data. SN Comput. Sci. 2022, 3, 276. [Google Scholar] [CrossRef]
- Majkowski, A.; Kołodziej, M.; Rak, R.J.; Rysz, A. Implementation of ECoG Signal Energy, Entropy and Fractal Dimension for Spike Detection. In Proceedings of the IEEE International Symposium on Medical Measurements and Applications (MeMeA), Rome, Italy, 11–13 June 2018; pp. 1–6. [Google Scholar]
- Sergeev, K.; Runnova, A.; Zhuravlev, M.; Sitnikova, E.; Rutskova, E.; Smirnov, K.; Slepnev, A.; Semenova, N. Simple method for detecting sleep episodes in rats ECoG using machine learning. Chaos Solitons Fractals 2023, 173, 113608. [Google Scholar] [CrossRef]
- Semenova, N.; Segreev, K.; Slepnev, A.; Runnova, A.; Zhuravlev, M.; Blokhina, I.; Dubrovsky, A.; Klimova, M.; Terskov, A.; Semyachkina-Glushkovskaya, O.; et al. Blood-brain barrier permeability changes: Nonlinear analysis of ECoG based on wavelet and machine learning approaches. Eur. Phys. J. Plus 2021, 136, 736. [Google Scholar] [CrossRef]
- GitHub. Available online: https://github.com/NSresearch/ANN-for-BBB-spectra/ (accessed on 29 September 2023).
- Welch, P. The use of fast Fourier transform for the estimation of power spectra: A method based on time averaging over short, modified periodograms. IEEE Trans. Audio Electroacoust. 1967, 15, 70–73. [Google Scholar] [CrossRef]
- Liu, C.W.; Wang, E.Y.; Wang, H.L.; Liao, K.H.; Chen, H.Y.; Chen, H.S.; Lai, T.W. Blood-Brain Barrier Disruption in Preclinical Mouse Models of Stroke Can Be an Experimental Artifact Caused by Craniectomy. eNeuro 2022, 5, 0343-22. [Google Scholar] [CrossRef]
- Semyachkina-Glushkovskaya, O.; Esmat, A.; Bragin, D.; Bragina, O.; Shirokov, A.A.; Navolokin, N.; Yang, Y.; Abdurashitov, A.; Khorovodov, A.; Terskov, A.; et al. Phenomenon of music-induced opening of the blood-brain barrier in healthy mice. Proc. R. Soc. B 2020, 287, 20202337. [Google Scholar] [CrossRef]
- Semyachkina-Glushkovskaya, O.; Khorovodov, A.; Fedosov, I.; Pavlov, A.; Shirokov, A.; Sharif, A.E.; Dubrovsky, A.; Blokhina, I.; Terskov, A.; Navolokin, N.; et al. A Novel Method to Stimulate Lymphatic Clearance of Beta-Amyloid from Mouse Brain Using Noninvasive Music-Induced Opening of the Blood–Brain Barrier with EEG Markers. Appl. Sci. 2021, 11, 10287. [Google Scholar] [CrossRef]
- Hablitz, L.M.; Vinitsky, H.S.; Sun, Q.; Stæger, F.F.; Sigurdsson, B.; Mortensen, K.N.; Lilius, T.O.; Nedergaard, M. Increased glymphatic influx is correlated with high EEG delta power and low heart rate in mice under anesthesia. Sci. Adv. 2019, 5, 5447. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Dong, Y.; Maeda, U.; Moir, R.D.; Xia, W.; Culley, D.J.; Crosby, G.; Tanzi, R.E. The inhalation anesthetic isoflurane induces a vicious cycle of apoptosis and amyloid beta-protein accumulation. J. Neurosci. 2007, 27, 1247–1254. [Google Scholar] [CrossRef] [PubMed]
- Gakuba, C.; Gaberel, T.; Goursaud, S.; Bourges, J.; Di Palma, C.; Quenault, A.; Martinez de Lizarrondo, S.; Vivien, D.; Gauberti, M. General Anesthesia Inhibits the Activity of the “Glymphatic System”. Theranostics 2018, 8, 710–722. [Google Scholar] [CrossRef] [PubMed]
- Semyachkina-Glushkovskaya, O.V.; Postnov, D.E.; Khorovodov, A.P.; Navolokin, N.A.; Kurthz, J.H. Lymphatic Drainage System of the Brain: A New Player in Neuroscience. J. Evol. Biochem. Phys. 2023, 59, 1–19. [Google Scholar] [CrossRef]
- Hladky, S.B.; Barrand, M.A. The glymphatic hypothesis: The theory and the evidence. Fluids Barriers CNS 2022, 19, 9. [Google Scholar] [CrossRef]
- Abbot, N.J.; Pizzo, M.; Preston, J.E.; Janigro, D.; Thorne, R.G. The role of brain barriers in fluid movement in the CNS: Is there a ‘glymphatic system’. Acta Neuropathol. 2018, 135, 387–407. [Google Scholar] [CrossRef]
- Smith, A.; Jin, B.J.; Verkman, A.S. Muddying the water in brain edema? Trends Neurosci. 2015, 38, 331–332. [Google Scholar] [CrossRef]
- Dupont, G.; Schmidt, C.; Yilmaz, E.; Oskouian, R.J.; Macchi, V.; de Caro, R.; Tubbs, R.S. Our current understanding of the lymphatics of the brain and spinal cord. Clin. Anat. 2019, 32, 117–121. [Google Scholar] [CrossRef]
- Hladky, S.B.; Barrand, M.A. Mechanisms of fluid movement into, through and out of the brain: Evaluation of the evidence. Fluids Barriers CNS 2014, 11, 26. [Google Scholar] [CrossRef]
- Weller, R.O.; Galea, I.; Carare, R.O.; Minagar, A. Pathophysiology of the lymphatic drainage of the central nervous system: Implications for pathogenesis and therapy of multiple sclerosis. Pathophysiology 2010, 17, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Haorah, J. How does the brain remove its waste metabolites from within. Int. J. Physiol. Pathophysiol. Pharmacol. 2019, 11, 238–249. [Google Scholar]
- Li, R.; Zhao, M.; Yao, D.; Zhou, X.; Lenahan, C.; Wang, L.; Ou, Y.; He, Y. The role of the astrocyte in subarachnoid hemorrhage and its therapeutic implications. Front. Immunol. 2022, 13, 1008795. [Google Scholar] [CrossRef] [PubMed]
- Pasantes-Morales, H.; Tuz, K. Volume changes in neurons: Hyperexcitability and neuronal death. Contri. Nebhrol. 2006, 152, 221–240. [Google Scholar]
- Hübel, N.; Ullah, G. Anions govern cell volume: A case study of relative astrocytic and neuronal swelling in spreading depolarization. PLoS ONE 2016, 11, e0147060. [Google Scholar] [CrossRef] [PubMed]
- Florence, C.M.; Baillie, L.D.; Mulligan, S.J. Dynamic volume changes in astrocytes are an intrinsic phenomenon mediated by bicarbonate ion flux. PLoS ONE 2012, 7, e51124. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chen, Y.; Feng, W.; Cai, J.; Gao, J.; Ge, F.; Zhou, T.; Wang, Z.; Ding, F.; Marshall, C.; et al. Drainage of senescent astrocytes from brain via meningeal lymphatic routes. Brain Behav. Immun. 2022, 103, 85–96. [Google Scholar] [CrossRef]
- Kuroda, T.; Matsuda, N.; Ishibashi, Y.; Suzuki, I. Detection of astrocytic slow oscillatory activity and response to seizurogenic compounds using planar microelectrode array. Front. Neurosci. 2023, 16, 1050150. [Google Scholar] [CrossRef]
- Ikeda, A.; Takeyama, H.; Bernard, C.; Nakatani, M.; Shimotake, A.; Daifu, M.; Matsuhashi, M.; Kikuchi, T.; Kunieda, T.; Matsumoto, R.; et al. Active direct current (DC) shifts and “Red slow”: Two new concepts for seizure mechanisms and identification of the epileptogenic zone. Neurosci. Res. 2020, 156, 95–101. [Google Scholar] [CrossRef]
- Michinaga, S.; Koyama, Y. Dual Roles of Astrocyte-Derived Factors in Regulation of Blood-Brain Barrier Function after Brain Damage. Int. J. Mol. Sci. 2019, 20, 571. [Google Scholar] [CrossRef]
- Toth, A.E.; Nielsen, M.S. Sortilins in the blood-brain barrier: Impact on barrier integrity. Neural Regen. Res. 2023, 18, 549–550. [Google Scholar] [PubMed]
- Buskila, Y.; Bellot-Saez, A.; Morley, J.W. Generating brain waves, the power of astrocytes. Front. Neurosci. 2019, 13, 1125. [Google Scholar] [CrossRef]
- Henneberger, C.; Papouin, T.; Oliet, S.H.R.; Rusacov, D. Long-term potentiation depends on release of D-serine from astrocytes. Nature 2010, 463, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Takata, N.; Mishima, T.; Hisatsune, C.; Nagai, Y.; Ebisui, E.; Mikoshiba, K.; Hirase, H. Astrocyte calcium signaling transforms cholinergic modulation to cortical plasticity in vivo. J. Neurosci. 2011, 31, 18155–18165. [Google Scholar] [CrossRef]
- Bellesi, M.; Vivo, L.; Tononi, G.; Cirelli, C. Effects of sleep and wake on astrocytes: Clues from molecular and ultrastructural studies. BMC Biol. 2015, 13, 66. [Google Scholar] [CrossRef]
- Rudolph, M.; Pelletier, J.G.; Paré, D.; Destexhe, A. Characterization of synaptic conductances and integrative properties during electrically induced EEG-activated states in neocortical neurons in vivo. J. Neurophysiol. 2005, 94, 2805–2821. [Google Scholar] [CrossRef]
- Kiviniemi, V.; Korhonen, V.; Kortelainen, J.; Rytky, S.; Keinänen, T.; Tuovinen, T.; Isokangas, M.; Sonkajärvi, E.; Siniluoto, T.; Nikkinen, J.; et al. Real-time monitoring of human blood-brain barrier disruption. PLoS ONE 2017, 3, e0174072. [Google Scholar] [CrossRef]
- Lipsman, N.; Meng, Y.; Bethune, A.; Huang, Y.; Lam, B.; Masellis, M.; Herrmann, N.; Heyn, N.; Aubert, I.; Boutet, A.; et al. Blood–brain barrier opening in Alzheimer’s disease using MR-guided focused ultrasound. Nat. Commun. 2018, 9, 2336. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semyachkina-Glushkovskaya, O.; Sergeev, K.; Semenova, N.; Slepnev, A.; Karavaev, A.; Hramkov, A.; Prokhorov, M.; Borovkova, E.; Blokhina, I.; Fedosov, I.; et al. Machine Learning Technology for EEG-Forecast of the Blood–Brain Barrier Leakage and the Activation of the Brain’s Drainage System during Isoflurane Anesthesia. Biomolecules 2023, 13, 1605. https://doi.org/10.3390/biom13111605
Semyachkina-Glushkovskaya O, Sergeev K, Semenova N, Slepnev A, Karavaev A, Hramkov A, Prokhorov M, Borovkova E, Blokhina I, Fedosov I, et al. Machine Learning Technology for EEG-Forecast of the Blood–Brain Barrier Leakage and the Activation of the Brain’s Drainage System during Isoflurane Anesthesia. Biomolecules. 2023; 13(11):1605. https://doi.org/10.3390/biom13111605
Chicago/Turabian StyleSemyachkina-Glushkovskaya, Oxana, Konstantin Sergeev, Nadezhda Semenova, Andrey Slepnev, Anatoly Karavaev, Alexey Hramkov, Mikhail Prokhorov, Ekaterina Borovkova, Inna Blokhina, Ivan Fedosov, and et al. 2023. "Machine Learning Technology for EEG-Forecast of the Blood–Brain Barrier Leakage and the Activation of the Brain’s Drainage System during Isoflurane Anesthesia" Biomolecules 13, no. 11: 1605. https://doi.org/10.3390/biom13111605
APA StyleSemyachkina-Glushkovskaya, O., Sergeev, K., Semenova, N., Slepnev, A., Karavaev, A., Hramkov, A., Prokhorov, M., Borovkova, E., Blokhina, I., Fedosov, I., Shirokov, A., Dubrovsky, A., Terskov, A., Manzhaeva, M., Krupnova, V., Dmitrenko, A., Zlatogorskaya, D., Adushkina, V., Evsukova, A., ... Kurths, J. (2023). Machine Learning Technology for EEG-Forecast of the Blood–Brain Barrier Leakage and the Activation of the Brain’s Drainage System during Isoflurane Anesthesia. Biomolecules, 13(11), 1605. https://doi.org/10.3390/biom13111605