
C-C Motif Chemokine Ligand 2 Enhances Macrophage Chemotaxis, Osteogenesis, and Angiogenesis during the Inflammatory Phase of Bone Regeneration

 , , ,
, , ,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rabbit MSCs
2.2. Preparation of Rabbit CCL2 Plasmid
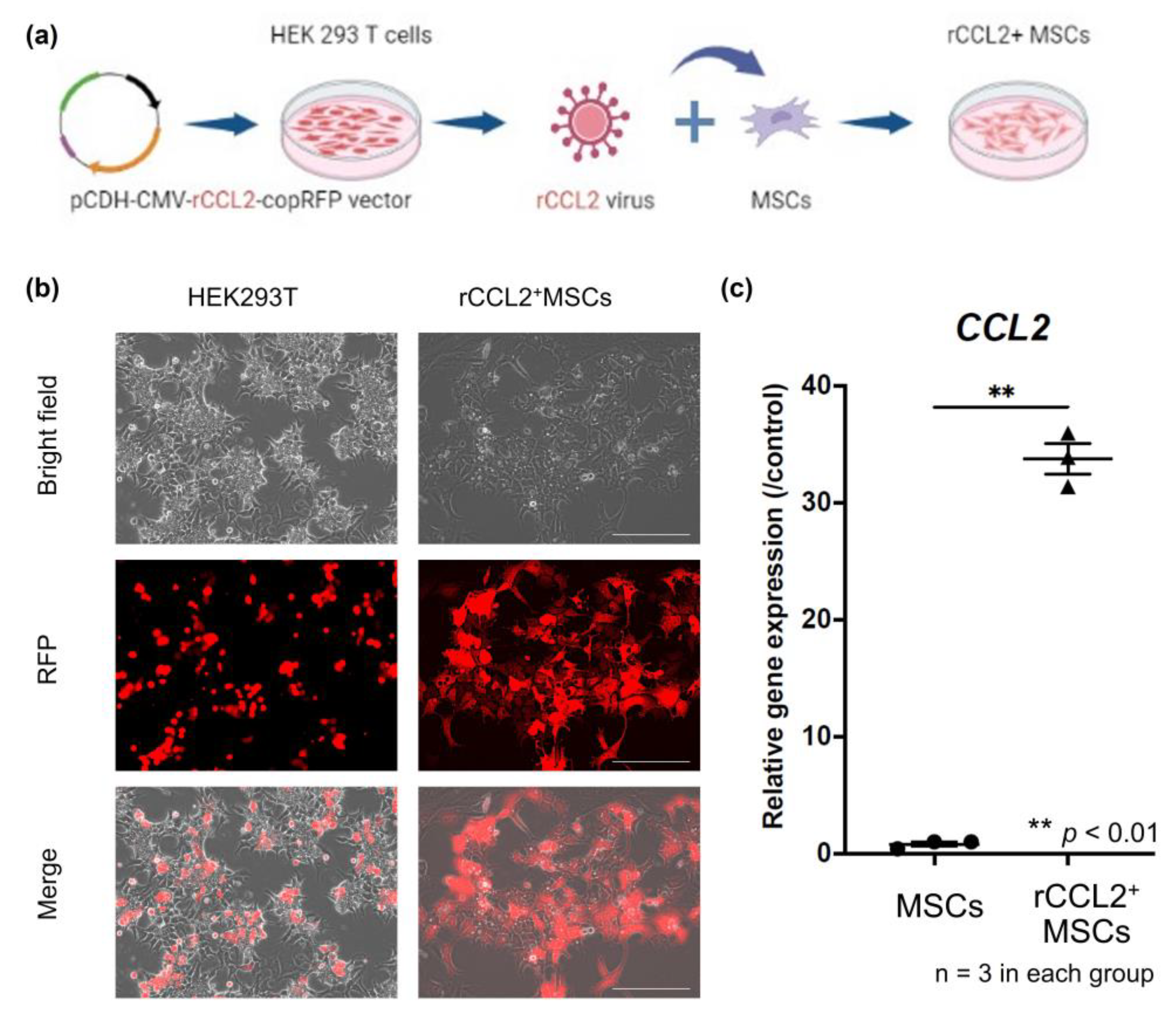
2.3. Establishment of Genetically Modified Rabbit MSCs
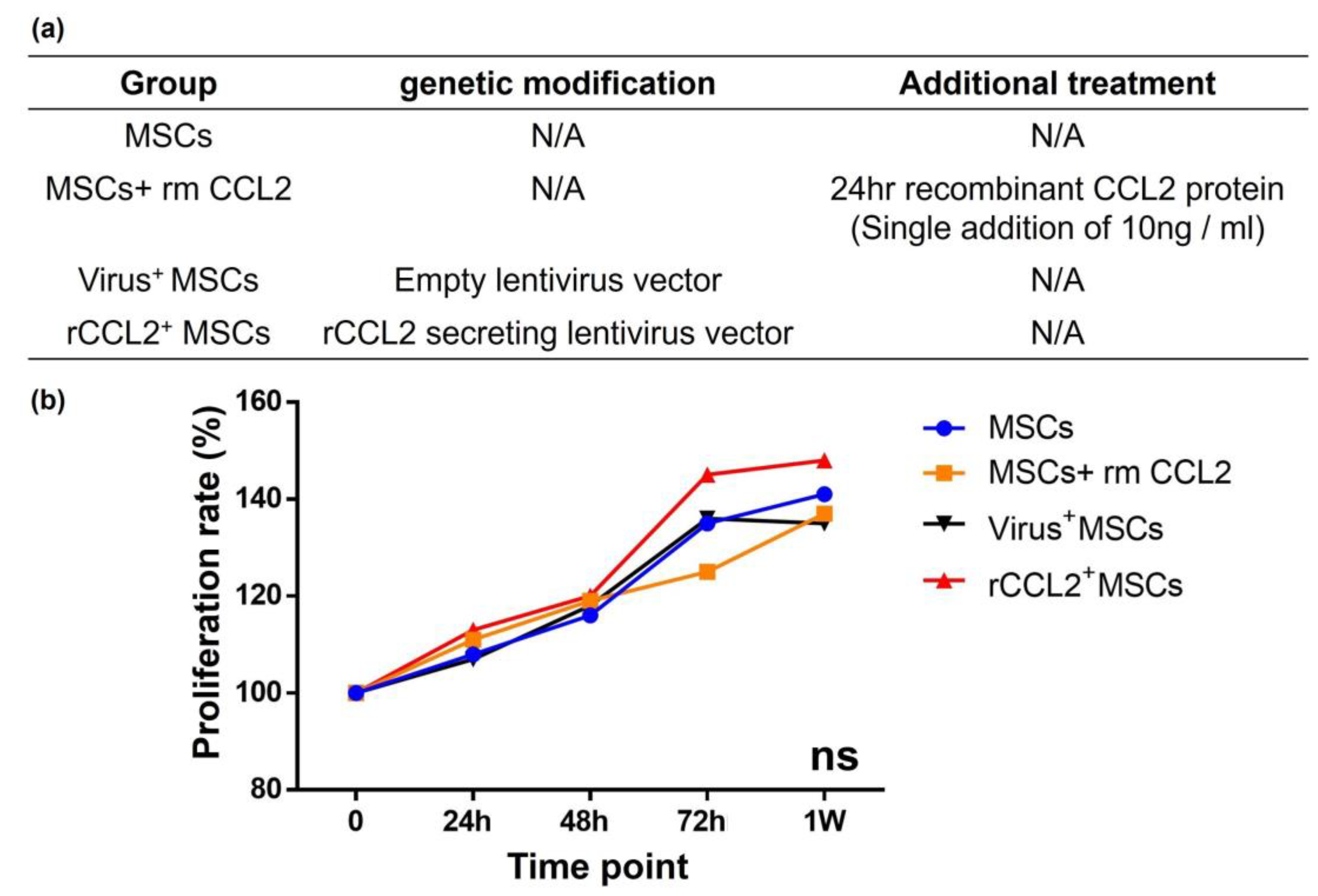
2.4. Cell Cultures and Groups
2.5. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
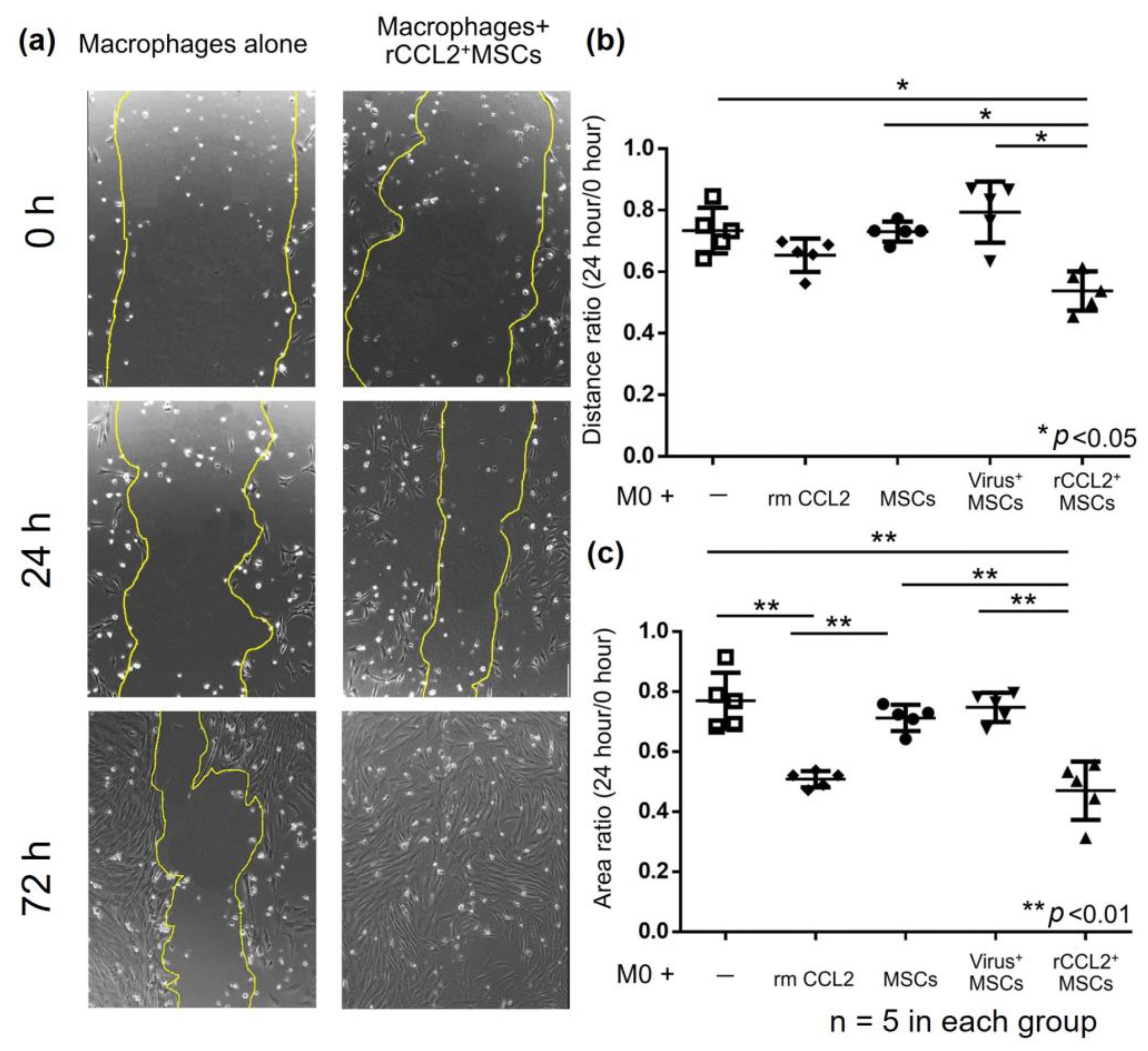
2.6. Migration Assay
2.7. Cell Proliferation Assay
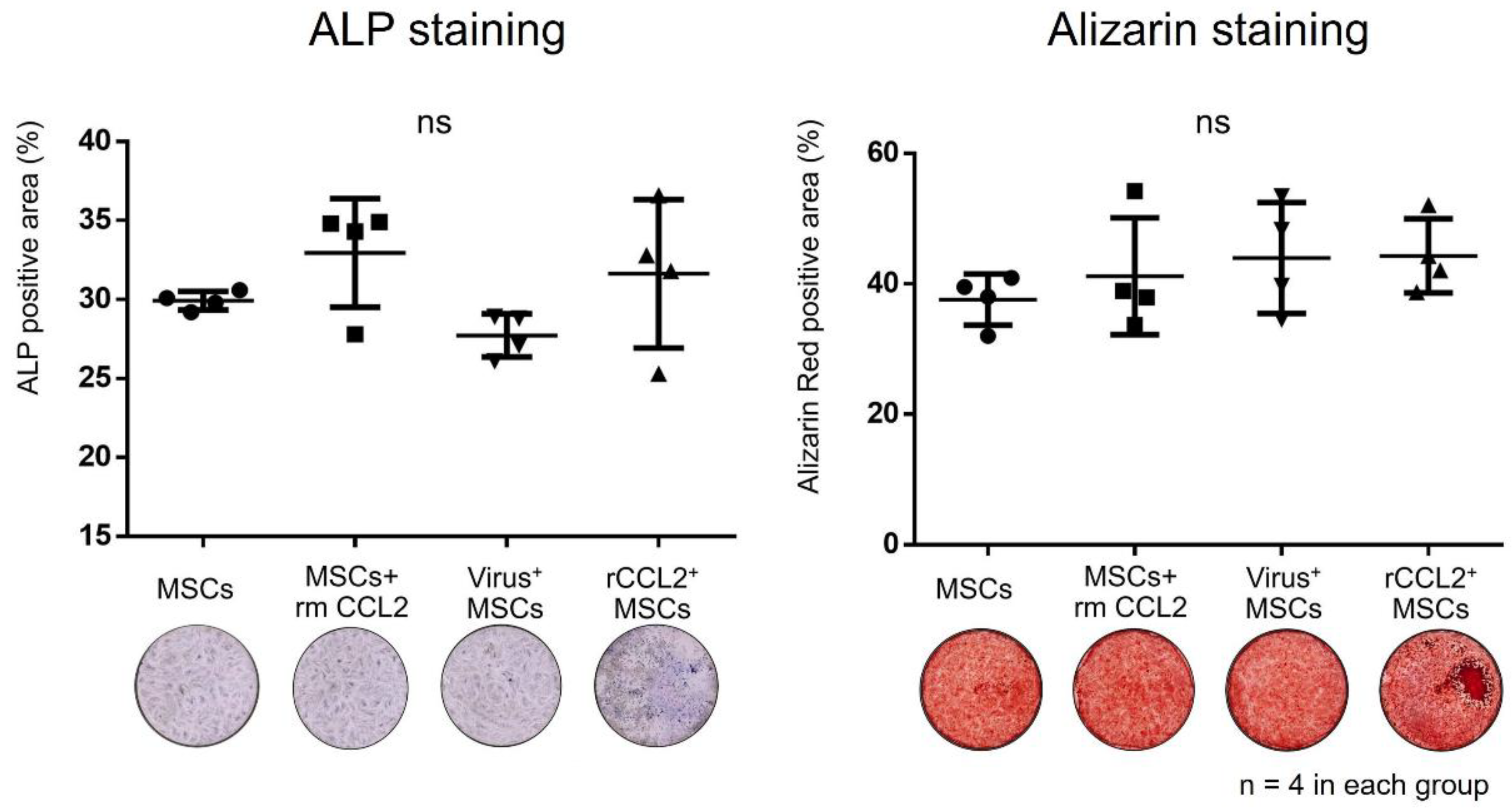
2.8. Osteogenic Differentiation Assay
2.9. Tube Formation Assay
2.10. Statistical Analysis
3. Results
3.1. CCL2 Gene Expression
3.2. Cell Proliferation
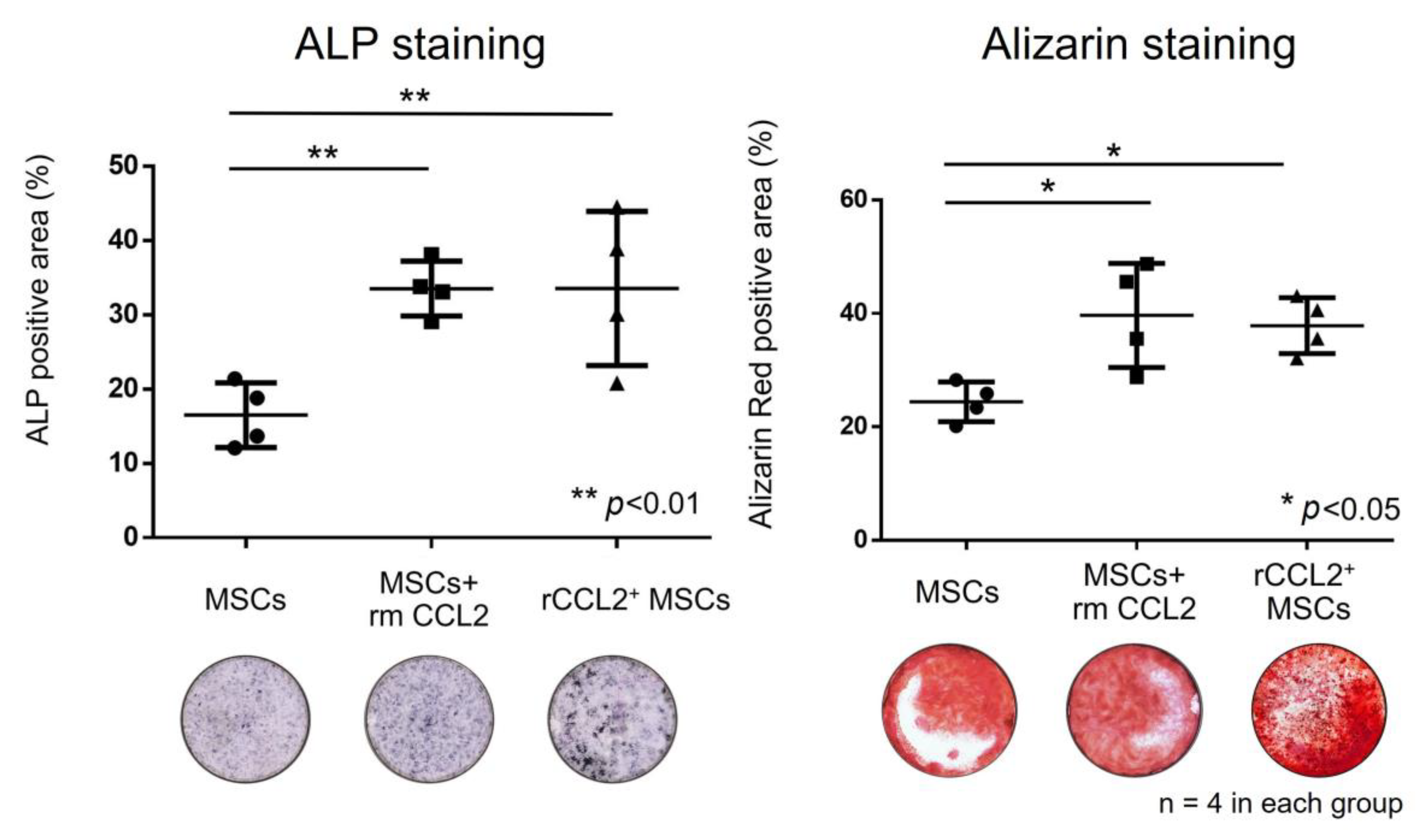
3.3. Osteogenic Differentiation (Monoculture)
3.4. Migration Assay
3.5. Osteogenic Differentiation (Co-Culture)
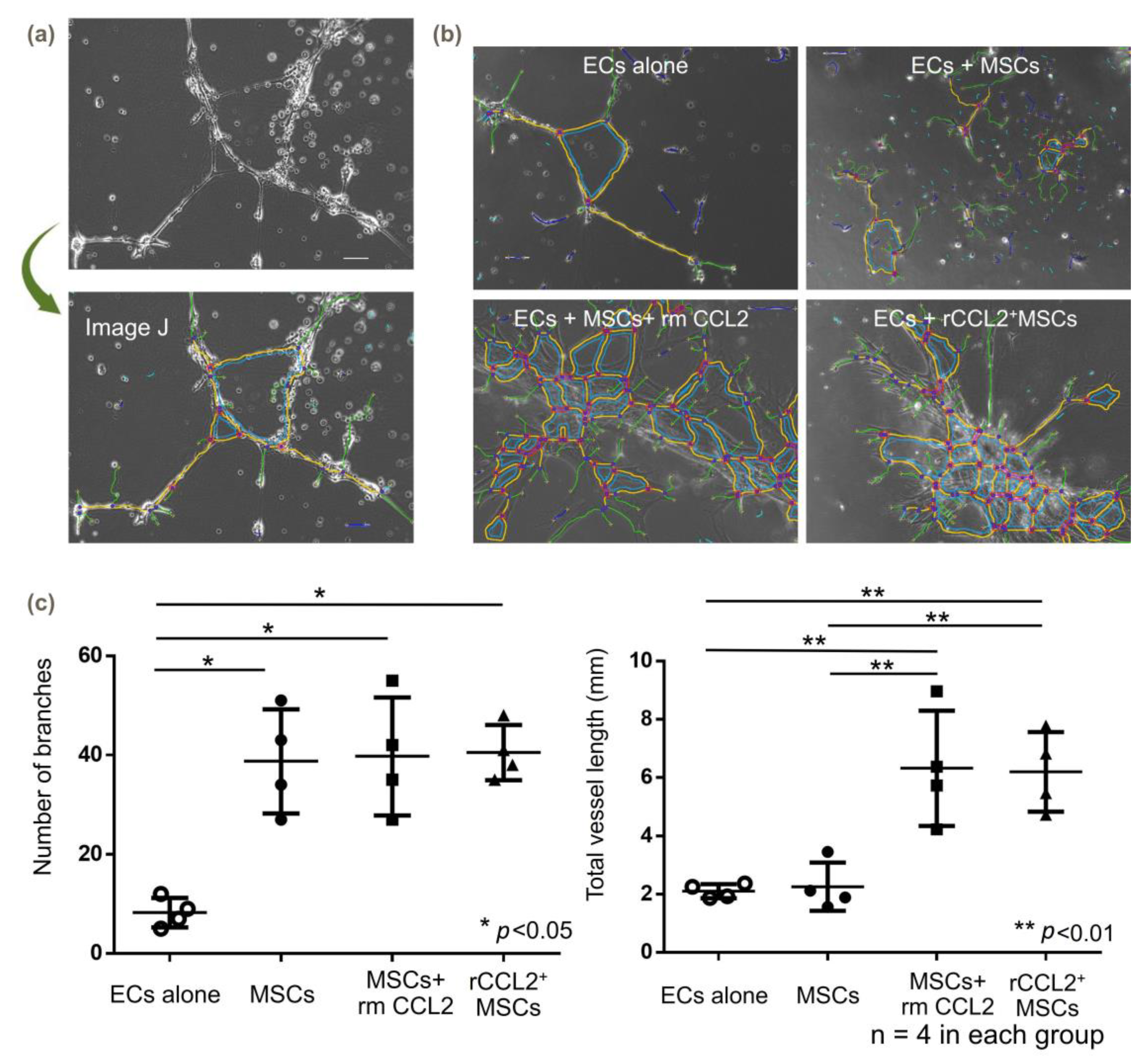
3.6. Angiogenesis (Tube Formation Assay)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Onggo, J.R.; Nambiar, M.; Onggo, J.D.; Tay, G.; Singh, P.J.; Babazadeh, S. Outcome of tantalum rod insertion in the treatment of osteonecrosis of the femoral head with minimum follow-up of 1 year: A meta-analysis and systematic review. J. Hip Preserv. Surg. 2020, 7, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.H.; Jones, L.C.; Chen, C.H.; Cheng, E.Y.; Cui, Q.; Drescher, W.; Fukushima, W.; Gangji, V.; Goodman, S.B.; Ha, Y.C.; et al. Etiologic Classification Criteria of ARCO on Femoral Head Osteonecrosis Part 1: Glucocorticoid-Associated Osteonecrosis. J. Arthroplasty 2019, 34, 163–168.e161. [Google Scholar] [CrossRef]
- Maruyama, M.; Lin, T.; Pan, C.C.; Moeinzadeh, S.; Takagi, M.; Yang, Y.P.; Goodman, S.B. Cell-Based and Scaffold-Based Therapies for Joint Preservation in Early-Stage Osteonecrosis of the Femoral Head: A Review of Basic Research. JBJS Rev. 2019, 7, e5. [Google Scholar] [CrossRef] [PubMed]
- Gangji, V.; Hauzeur, J.P.; Schoutens, A.; Hinsenkamp, M.; Appelboom, T.; Egrise, D. Abnormalities in the replicative capacity of osteoblastic cells in the proximal femur of patients with osteonecrosis of the femoral head. J. Rheumatol. 2003, 30, 348–351. [Google Scholar]
- Mont, M.A.; Hungerford, D.S. Non-traumatic avascular necrosis of the femoral head. J. Bone Joint Surg. Am. 1995, 77, 459–474. [Google Scholar] [CrossRef] [PubMed]
- Ito, H. Chemokines in mesenchymal stem cell therapy for bone repair: A novel concept of recruiting mesenchymal stem cells and the possible cell sources. Mod. Rheumatol. 2011, 21, 113–121. [Google Scholar] [CrossRef]
- Otsuru, S.; Tamai, K.; Yamazaki, T.; Yoshikawa, H.; Kaneda, Y. Circulating bone marrow-derived osteoblast progenitor cells are recruited to the bone-forming site by the CXCR4/stromal cell-derived factor-1 pathway. Stem Cells 2008, 26, 223–234. [Google Scholar] [CrossRef]
- Pajarinen, J.; Lin, T.; Gibon, E.; Kohno, Y.; Maruyama, M.; Nathan, K.; Lu, L.; Yao, Z.; Goodman, S.B. Mesenchymal stem cell-macrophage crosstalk and bone healing. Biomaterials 2019, 196, 80–89. [Google Scholar] [CrossRef]
- Bouland, C.; Philippart, P.; Dequanter, D.; Corrillon, F.; Loeb, I.; Bron, D.; Lagneaux, L.; Meuleman, N. Cross-Talk Between Mesenchymal Stromal Cells (MSCs) and Endothelial Progenitor Cells (EPCs) in Bone Regeneration. Front. Cell Dev. Biol. 2021, 9, 674084. [Google Scholar] [CrossRef]
- Hao, Q.; Vadgama, J.V.; Wang, P. CCL2/CCR2 signaling in cancer pathogenesis. Cell Commun. Signal 2020, 18, 82. [Google Scholar] [CrossRef]
- Toya, M.; Zhang, N.; Tsubosaka, M.; Kushioka, J.; Gao, Q.; Li, X.; Chow, S.K.; Goodman, S.B. CCL2 promotes osteogenesis by facilitating macrophage migration during acute inflammation. Front. Cell Dev. Biol. 2023, 11, 1213641. [Google Scholar] [CrossRef]
- Jiang, Y.; Graves, D.T. Periodontal pathogens stimulate CC-chemokine production by mononuclear and bone-derived cells. J. Periodontol. 1999, 70, 1472–1478. [Google Scholar] [CrossRef]
- Stamatovic, S.M.; Keep, R.F.; Mostarica-Stojkovic, M.; Andjelkovic, A.V. CCL2 regulates angiogenesis via activation of Ets-1 transcription factor. J. Immunol. 2006, 177, 2651–2661. [Google Scholar] [CrossRef]
- Ridiandries, A.; Tan, J.T.; Bursill, C.A. The Role of CC-Chemokines in the Regulation of Angiogenesis. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Lo, C.W.; Utsunomiya, T.; Maruyama, M.; Huang, E.; Rhee, C.; Gao, Q.; Yao, Z.; Goodman, S.B. PDGF-BB and IL-4 co-overexpression is a potential strategy to enhance mesenchymal stem cell-based bone regeneration. Stem Cell Res. Ther. 2021, 12, 40. [Google Scholar] [CrossRef] [PubMed]
- Longhin, E.M.; El Yamani, N.; Runden-Pran, E.; Dusinska, M. The alamar blue assay in the context of safety testing of nanomaterials. Front. Toxicol. 2022, 4, 981701. [Google Scholar] [CrossRef]
- Blatt, S.; Thiem, D.G.E.; Kyyak, S.; Pabst, A.; Al-Nawas, B.; Kammerer, P.W. Possible Implications for Improved Osteogenesis? The Combination of Platelet-Rich Fibrin With Different Bone Substitute Materials. Front. Bioeng. Biotechnol. 2021, 9, 640053. [Google Scholar] [CrossRef] [PubMed]
- Kauanova, S.; Urazbayev, A.; Vorobjev, I. The Frequent Sampling of Wound Scratch Assay Reveals the “Opportunity” Window for Quantitative Evaluation of Cell Motility-Impeding Drugs. Front. Cell Dev. Biol. 2021, 9, 640972. [Google Scholar] [CrossRef]
- Bankhead, P.; Loughrey, M.B.; Fernandez, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef] [PubMed]
- Gregory, C.A.; Gunn, W.G.; Peister, A.; Prockop, D.J. An Alizarin red-based assay of mineralization by adherent cells in culture: Comparison with cetylpyridinium chloride extraction. Anal. Biochem. 2004, 329, 77–84. [Google Scholar] [CrossRef]
- Li, F.; Gou, X.; Xu, D.; Han, D.; Hou, K.; Fang, W.; Li, Y. Improvement of tube formation model of cell: Application for acute hypoxia in in vitro study of angiogenesis. Microvasc. Res. 2022, 140, 104297. [Google Scholar] [CrossRef] [PubMed]
- Kang, I.; Lee, B.C.; Choi, S.W.; Lee, J.Y.; Kim, J.J.; Kim, B.E.; Kim, D.H.; Lee, S.E.; Shin, N.; Seo, Y.; et al. Donor-dependent variation of human umbilical cord blood mesenchymal stem cells in response to hypoxic preconditioning and amelioration of limb ischemia. Exp. Mol. Med. 2018, 50, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, M.; Hayakawa, N.; Suzuki, M.; Mihara, M. IL-6/sIL-6R trans-signalling, but not TNF-alpha induced angiogenesis in a HUVEC and synovial cell co-culture system. Rheumatol. Int. 2009, 29, 1449–1454. [Google Scholar] [CrossRef] [PubMed]
- Carpentier, G.; Berndt, S.; Ferratge, S.; Rasband, W.; Cuendet, M.; Uzan, G.; Albanese, P. Angiogenesis Analyzer for ImageJ—A comparative morphometric analysis of “Endothelial Tube Formation Assay” and “Fibrin Bead Assay”. Sci. Rep. 2020, 10, 11568. [Google Scholar] [CrossRef]
- Singh, S.; Anshita, D.; Ravichandiran, V. MCP-1: Function, regulation, and involvement in disease. Int. Immunopharmacol. 2021, 101, 107598. [Google Scholar] [CrossRef]
- Zhu, S.; Liu, M.; Bennett, S.; Wang, Z.; Pfleger, K.D.G.; Xu, J. The molecular structure and role of CCL2 (MCP-1) and C-C chemokine receptor CCR2 in skeletal biology and diseases. J. Cell Physiol. 2021, 236, 7211–7222. [Google Scholar] [CrossRef]
- Robinson, W.H.; Lepus, C.M.; Wang, Q.; Raghu, H.; Mao, R.; Lindstrom, T.M.; Sokolove, J. Low-grade inflammation as a key mediator of the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 580–592. [Google Scholar] [CrossRef]
- Appleton, C.T.; Usmani, S.E.; Pest, M.A.; Pitelka, V.; Mort, J.S.; Beier, F. Reduction in disease progression by inhibition of transforming growth factor alpha-CCL2 signaling in experimental posttraumatic osteoarthritis. Arthritis Rheumatol. 2015, 67, 2691–2701. [Google Scholar] [CrossRef]
- Eraltan, H.; Cacina, C.; Kahraman, O.T.; Kurt, O.; Aydogan, H.Y.; Uyar, M.; Can, A.; Cakmakoglu, B. MCP-1 and CCR2 gene variants and the risk for osteoporosis and osteopenia. Genet. Test. Mol. Biomarkers 2012, 16, 229–233. [Google Scholar] [CrossRef]
- Rana, A.K.; Li, Y.; Dang, Q.; Yang, F. Monocytes in rheumatoid arthritis: Circulating precursors of macrophages and osteoclasts and, their heterogeneity and plasticity role in RA pathogenesis. Int. Immunopharmacol. 2018, 65, 348–359. [Google Scholar] [CrossRef]
- Graves, D.T.; Jiang, Y.; Valente, A.J. Regulated expression of MCP-1 by osteoblastic cells in vitro and in vivo. Histol. Histopathol. 1999, 14, 1347–1354. [Google Scholar] [CrossRef]
- Vasicek, J.; Balazi, A.; Tirpakova, M.; Svoradova, A.; Ondruska, L.; Parkanyi, V.; Chrenek, P. Secretome Analysis of Rabbit and Human Mesenchymal Stem and Endothelial Progenitor Cells: A Comparative Study. Int. J. Mol. Sci. 2021, 22, 12283. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, D.; Wang, H.; Chen, K.; Wang, S.; Xu, J.; Ji, P. Exosomes from adipose-derived stem cells regulate M1/M2 macrophage phenotypic polarization to promote bone healing via miR-451a/MIF. Stem Cell Res. Ther. 2022, 13, 149. [Google Scholar] [CrossRef] [PubMed]
- Sierra-Filardi, E.; Nieto, C.; Dominguez-Soto, A.; Barroso, R.; Sanchez-Mateos, P.; Puig-Kroger, A.; Lopez-Bravo, M.; Joven, J.; Ardavin, C.; Rodriguez-Fernandez, J.L.; et al. CCL2 shapes macrophage polarization by GM-CSF and M-CSF: Identification of CCL2/CCR2-dependent gene expression profile. J. Immunol. 2014, 192, 3858–3867. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, M.; Sul, O.J.; Choi, E.K.; Kim, J.E.; Suh, J.H.; Choi, H.S. MCP-1 deficiency enhances browning of adipose tissue via increased M2 polarization. J. Endocrinol. 2019, 242, 91–101. [Google Scholar] [CrossRef]
- Shahrara, S.; Amin, M.A.; Woods, J.M.; Haines, G.K.; Koch, A.E. Chemokine receptor expression and in vivo signaling pathways in the joints of rats with adjuvant-induced arthritis. Arthritis Rheum. 2003, 48, 3568–3583. [Google Scholar] [CrossRef]
- Odegaard, J.I.; Ricardo-Gonzalez, R.R.; Goforth, M.H.; Morel, C.R.; Subramanian, V.; Mukundan, L.; Red Eagle, A.; Vats, D.; Brombacher, F.; Ferrante, A.W.; et al. Macrophage-specific PPARgamma controls alternative activation and improves insulin resistance. Nature 2007, 447, 1116–1120. [Google Scholar] [CrossRef]
- Seebach, C.; Henrich, D.; Wilhelm, K.; Barker, J.H.; Marzi, I. Endothelial progenitor cells improve directly and indirectly early vascularization of mesenchymal stem cell-driven bone regeneration in a critical bone defect in rats. Cell Transplant. 2012, 21, 1667–1677. [Google Scholar] [CrossRef]
- Xu, L.; Zhou, J.; Liu, J.; Liu, Y.; Wang, L.; Jiang, R.; Diao, Z.; Yan, G.; Peault, B.; Sun, H.; et al. Different Angiogenic Potentials of Mesenchymal Stem Cells Derived from Umbilical Artery, Umbilical Vein, and Wharton’s Jelly. Stem Cells Int. 2017, 2017, 3175748. [Google Scholar] [CrossRef]
- Arderiu, G.; Pena, E.; Aledo, R.; Juan-Babot, O.; Badimon, L. Tissue factor regulates microvessel formation and stabilization by induction of chemokine (C-C motif) ligand 2 expression. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2607–2615. [Google Scholar] [CrossRef]
- Yu, Y.R.; Mao, L.; Piantadosi, C.A.; Gunn, M.D. CCR2 deficiency, dysregulation of Notch signaling, and spontaneous pulmonary arterial hypertension. Am. J. Respir. Cell Mol. Biol. 2013, 48, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Ehling, J.; Bartneck, M.; Wei, X.; Gremse, F.; Fech, V.; Mockel, D.; Baeck, C.; Hittatiya, K.; Eulberg, D.; Luedde, T.; et al. CCL2-dependent infiltrating macrophages promote angiogenesis in progressive liver fibrosis. Gut 2014, 63, 1960–1971. [Google Scholar] [CrossRef] [PubMed]
- Galvez, B.G.; Genis, L.; Matias-Roman, S.; Oblander, S.A.; Tryggvason, K.; Apte, S.S.; Arroyo, A.G. Membrane type 1-matrix metalloproteinase is regulated by chemokines monocyte-chemoattractant protein-1/ccl2 and interleukin-8/CXCL8 in endothelial cells during angiogenesis. J. Biol. Chem. 2005, 280, 1292–1298. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shinohara, I.; Tsubosaka, M.; Toya, M.; Lee, M.L.; Kushioka, J.; Murayama, M.; Gao, Q.; Li, X.; Zhang, N.; Chow, S.K.-H.; et al. C-C Motif Chemokine Ligand 2 Enhances Macrophage Chemotaxis, Osteogenesis, and Angiogenesis during the Inflammatory Phase of Bone Regeneration. Biomolecules 2023, 13, 1665. https://doi.org/10.3390/biom13111665
Shinohara I, Tsubosaka M, Toya M, Lee ML, Kushioka J, Murayama M, Gao Q, Li X, Zhang N, Chow SK-H, et al. C-C Motif Chemokine Ligand 2 Enhances Macrophage Chemotaxis, Osteogenesis, and Angiogenesis during the Inflammatory Phase of Bone Regeneration. Biomolecules. 2023; 13(11):1665. https://doi.org/10.3390/biom13111665
Chicago/Turabian StyleShinohara, Issei, Masanori Tsubosaka, Masakazu Toya, Max L. Lee, Junichi Kushioka, Masatoshi Murayama, Qi Gao, Xueping Li, Ning Zhang, Simon Kwoon-Ho Chow, and et al. 2023. "C-C Motif Chemokine Ligand 2 Enhances Macrophage Chemotaxis, Osteogenesis, and Angiogenesis during the Inflammatory Phase of Bone Regeneration" Biomolecules 13, no. 11: 1665. https://doi.org/10.3390/biom13111665
APA StyleShinohara, I., Tsubosaka, M., Toya, M., Lee, M. L., Kushioka, J., Murayama, M., Gao, Q., Li, X., Zhang, N., Chow, S. K. -H., Matsumoto, T., Kuroda, R., & Goodman, S. B. (2023). C-C Motif Chemokine Ligand 2 Enhances Macrophage Chemotaxis, Osteogenesis, and Angiogenesis during the Inflammatory Phase of Bone Regeneration. Biomolecules, 13(11), 1665. https://doi.org/10.3390/biom13111665