
A High-Throughput Assay for In Vitro Determination of Release Factor-Dependent Peptide Release from a Pretermination Complex by Fluorescence Anisotropy—Application to Nonsense Suppressor Screening and Mechanistic Studies



Abstract
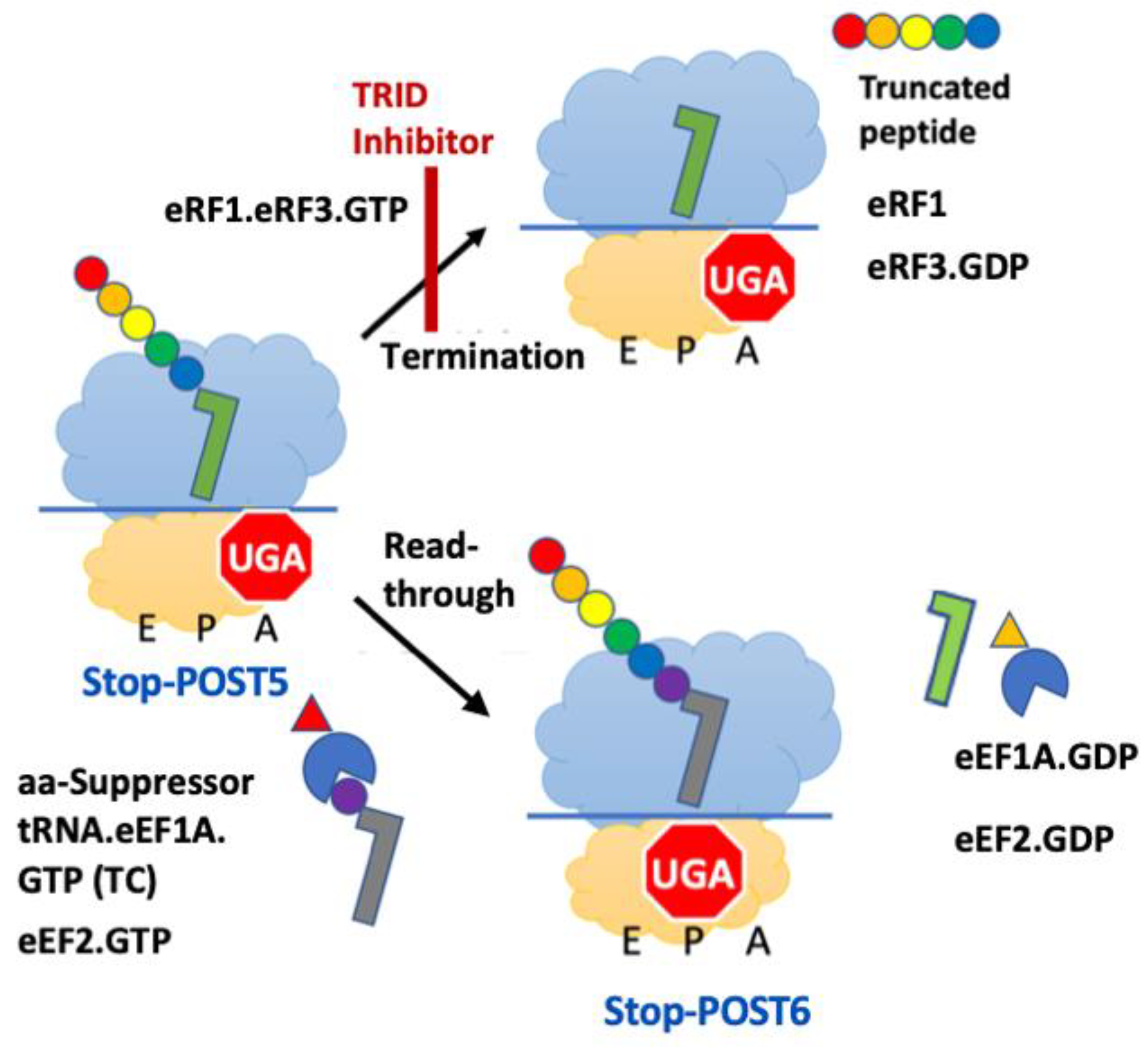
:1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Other Materials
2.1.2. Dye-Labeled [3H]-Lys-tRNALys Formation
2.1.3. Labeled Stop-POST5 Complexes
2.2. Methods
2.2.1. Peptide Release Assays
2.2.2. Thin Layer Electrophoresis
2.2.3. Mass Spectral Analysis of Released Labeled Peptide
3. Results
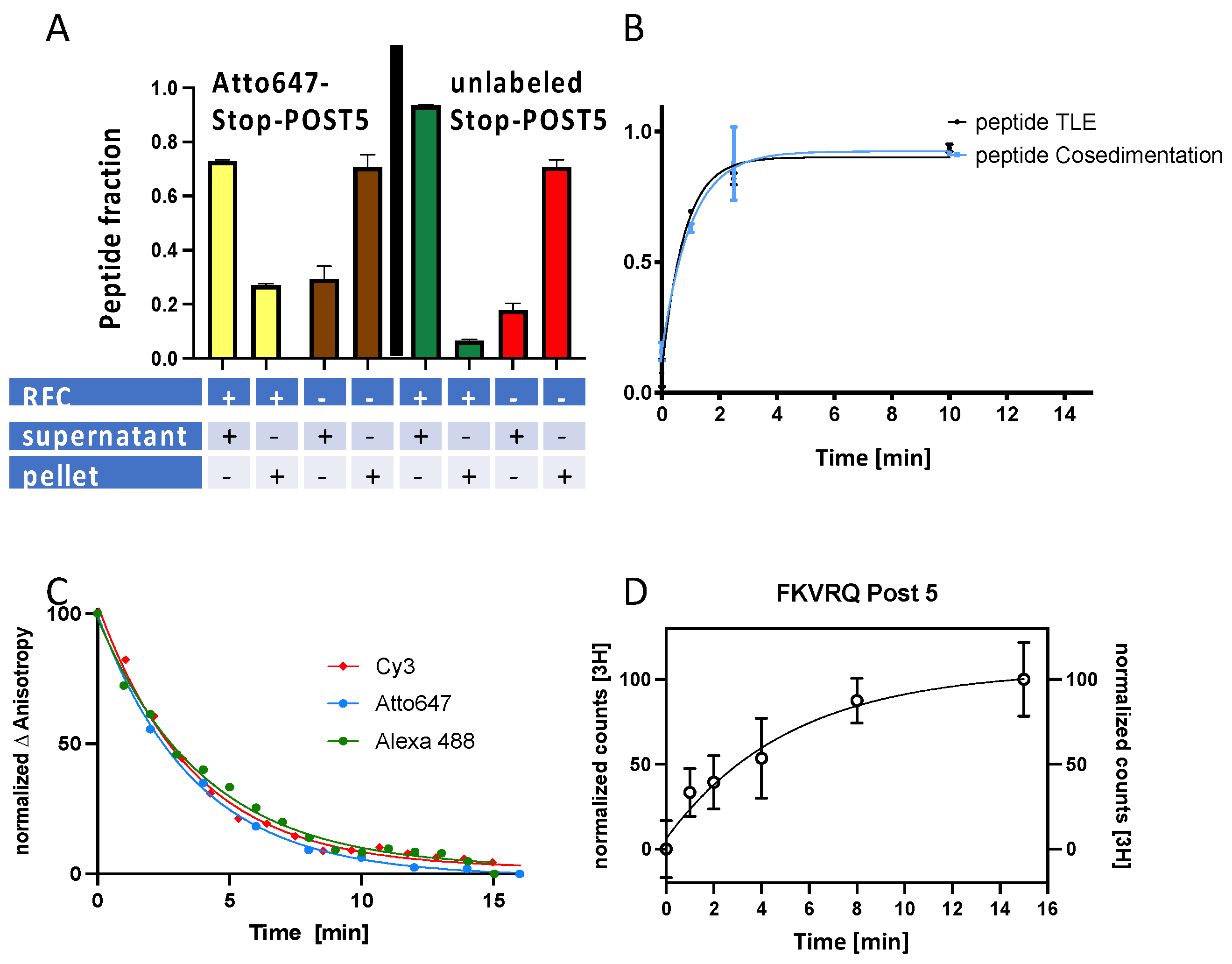
3.1. Incorporation of Fluorescent-Labeled Lys into the Stop-POST5 Complex
3.2. Dye Labeling Has No Substantial Effect on the Stoichiometry and Rates of RFC-Dependent Peptide Release from a Stop-POST5 Complex
3.3. Optimization of the Plate Reader Assay for High-Throughput (HT) Applications
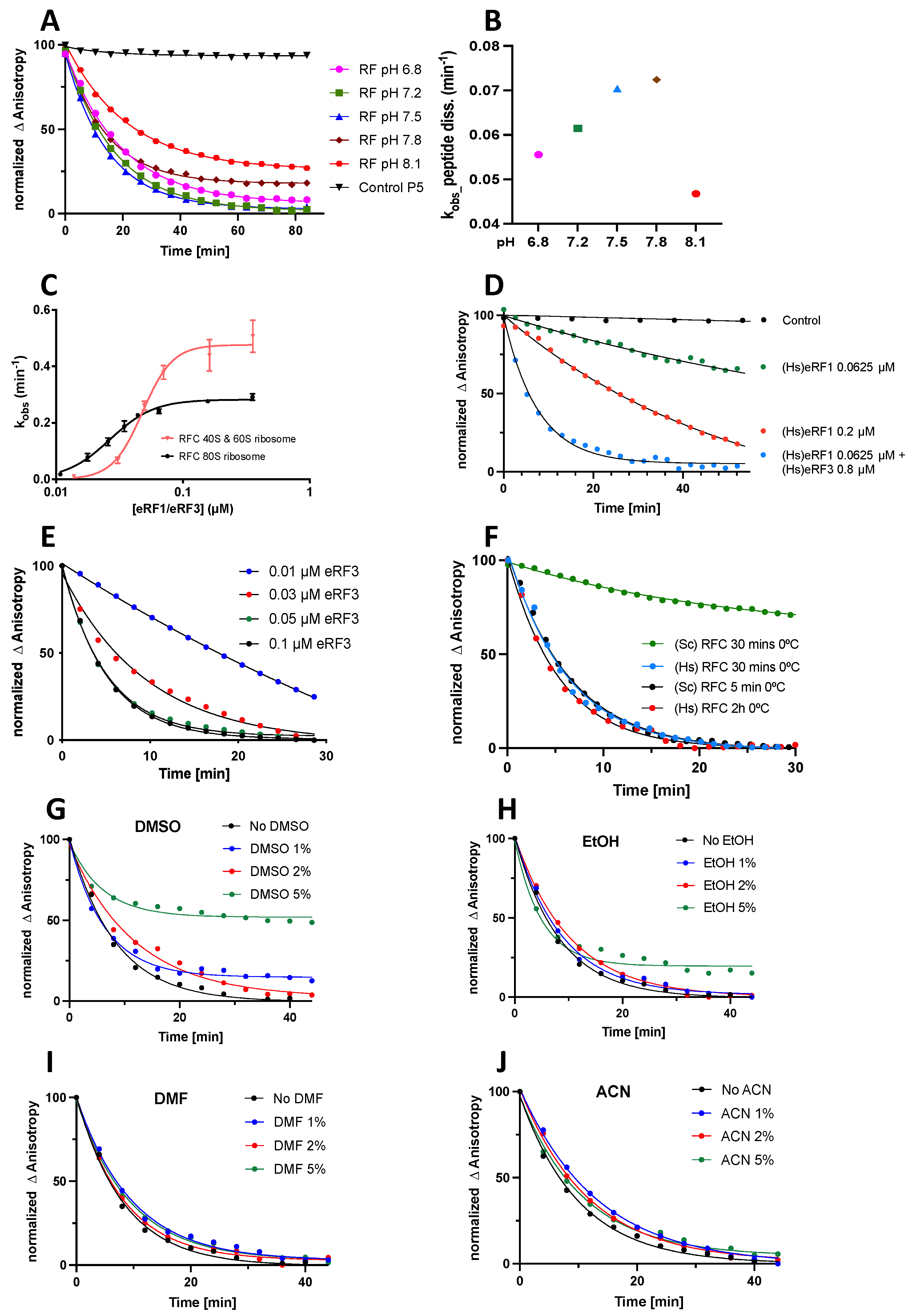
3.3.1. Results
- The pH optimum for the rate of peptide release falls in the range of 7.5–7.8 (Figure 3A,B);
- The apparent reactivity of Stop-POST5 complexes toward peptide release is somewhat higher for complexes prepared from purified 40S and 60S subunits than it is for complexes prepared by KCl treatment of 80S ribosomes, with the following EC50 and Vmax values: 40S + 60S: EC50 0.049 ± 0.003 µM, and Vmax 0.49 ± 0.02 min−1; 80S: EC50 0.029 ± 0.002 µM, and Vmax 0.31 ± 0.02 (Figure 3C);
- Although eRF1 alone can catalyze peptide release, much higher rates were obtained when it was combined with eRF3 (Figure 3D);
- At an eRF1 concentration of 0.03 µM, the addition of 0.05 µM eRF3 essentially enables the formation of an active RFC complex (Figure 3E);
- Yeast and human RFCs have equal reactivity when they are freshly prepared. However, yeast RFC is not cold stable, whereas human RFC is (Figure 3F);
- Several water miscible solvents (DMSO, EtOH, DMF, and ACN) added up to 2% (v/v) in the reaction mixture had little or no effect on the RFC reactivity. This range can be extended to 5% for DMF and ACN (Figure 3G–J).
3.3.2. Current Standard Conditions
3.4. Applications of the High-Throughput Assay
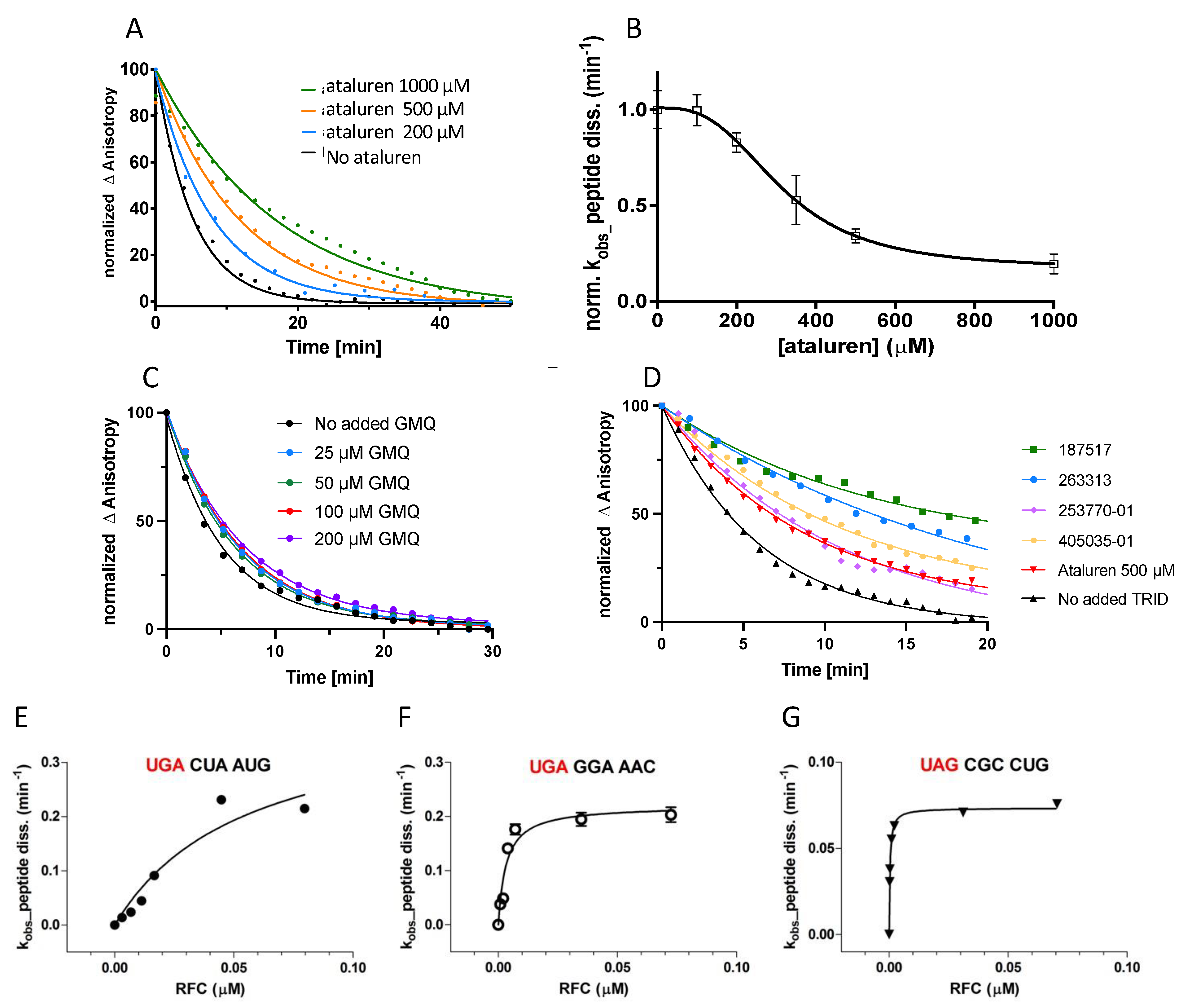
3.4.1. Screening New TRID Candidates
3.4.2. Determining the Effect of Stop Codon Identity and Downstream Codon Sequence on Peptide Release Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brenner, S.; Stretton, A.O.; Kaplan, S. Genetic code: The ‘nonsense’triplets for chain termination and their suppression. Nature 1965, 206, 994–998. [Google Scholar] [CrossRef] [PubMed]
- Shalev, M.; Baasov, T. When proteins start to make sense: Fine-tuning of aminoglycosides for PTC suppression therapy. MedChemComm 2014, 5, 1092–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keeling, K.M.; Xue, X.; Gunn, G.; Bedwell, D.M. Therapeutics based on stop codon readthrough. Annu. Rev. Genom. Hum. Genet. 2014, 15, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardi, S.; Testa, M.F.; Pinotti, M.; Branchini, A. Translation termination codons in protein synthesis and disease. Adv. Protein Chem. Struct. Biol. 2022, 132, 1–48. [Google Scholar]
- Salvatori, F.; Breveglieri, G.; Zuccato, C.; Finotti, A.; Bianchi, N.; Borgatti, M.; Feriotto, G.; Destro, F.; Canella, A.; Brognara, E.; et al. Production of β-globin and adult hemoglobin following G418 treatment of erythroid precursor cells from homozygous β039 thalassemia patients. Am. J. Hematol. 2009, 84, 720–728. [Google Scholar] [CrossRef] [Green Version]
- Goldmann, T.; Overlack, N.; Möller, F.; Belakhov, V.; van Wyk, M.; Baasov, T.; Wolfrum, U.; Nagel-Wolfrum, K. A comparative evaluation of NB30, NB54 and PTC124 in translational read-through efficacy for treatment of an USH1C nonsense mutation. EMBO Mol. Med. 2012, 4, 1186–1199. [Google Scholar] [CrossRef]
- Stenson, P.D.; Mort, M.; Ball, E.V.; Evans, K.; Hayden, M.; Heywood, S.; Hussain, M.; Phillips, A.D.; Cooper, D.N. The Human Gene Mutation Database: Towards a comprehensive repository of inherited mutation data for medical research, genetic diagnosis and next-generation sequencing studies. Hum. Genet. 2017, 136, 665–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mort, M.; Ivanov, D.; Cooper, D.N.; Chuzhanova, N.A. A meta-analysis of nonsense mutations causing human genetic disease. Hum. Mutat. 2008, 29, 1037–1047. [Google Scholar] [CrossRef]
- Martins-Dias, P.; Romão, L. Nonsense suppression therapies in human genetic diseases. Cell. Mol. Life Sci. 2021, 78, 4677–4701. [Google Scholar] [CrossRef]
- Zhang, H.; Ng, M.Y.; Chen, Y.; Cooperman, B.S. Kinetics of initiating polypeptide elongation in an IRES-dependent system. Elife 2016, 5, e13429. [Google Scholar] [CrossRef]
- Ng, M.Y.; Zhang, H.; Weil, A.; Singh, V.; Jamiolkowski, R.; Baradaran-Heravi, A.; Roberge, M.; Jacobson, A.; Friesen, W.; Welch, E.; et al. New in vitro assay measuring direct interaction of nonsense suppressors with the eukaryotic protein synthesis machinery. ACS Med. Chem. Lett. 2018, 9, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.Y.; Li, H.; Ghelfi, M.D.; Goldman, Y.E.; Cooperman, B.S. Ataluren and aminoglycosides stimulate read-through of nonsense codons by orthogonal mechanisms. Proc. Natl. Acad. Sci. USA 2021, 118, e2020599118. [Google Scholar] [CrossRef]
- Huang, S.; Bhattacharya, A.; Ghelfi, M.D.; Li, H.; Fritsch, C.; Chenoweth, D.M.; Goldman, Y.E.; Cooperman, B.S. Ataluren binds to multiple protein synthesis apparatus sites and competitively inhibits release factor-dependent termination. Nat. Commun. 2022, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Welch, E.M.; Barton, E.R.; Zhuo, J.; Tomizawa, Y.; Friesen, W.J.; Trifillis, P.; Paushkin, S.; Patel, M.; Trotta, C.R.; Hwang, S.; et al. PTC124 targets genetic disorders caused by nonsense mutations. Nature 2007, 447, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Peltz, S.W.; Morsy, M.; Welch, E.M.; Jacobson, A. Ataluren as an agent for therapeutic nonsense suppression. Annu. Rev. Med. 2013, 64, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leier, A.; Bedwell, D.M.; Chen, A.T.; Dickson, G.; Keeling, K.M.; Kesterson, R.A.; Korf, B.R.; Lago, T.T.; Müller, U.F.; Popplewell, L.; et al. Mutation-directed therapeutics for neurofibromatosis type I. Mol. Ther. Nucleic Acids 2020, 5, 739–753. [Google Scholar] [CrossRef]
- Oishi, N.; Duscha, S.; Boukari, H.; Meyer, M.; Xie, J.; Wei, G.; Schrepfer, T.; Roschitzki, B.; Böttger, E.C.; Schacht, J. XBP1 mitigates aminoglycoside-induced endoplasmic reticulum stress and neuronal cell death. Cell Death Dis. 2015, 6, e1763. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, N.; Sonenberg, N. Proposing a mechanism of action for ataluren. Proc. Natl. Acad. Sci. USA 2016, 113, 12353–12355. [Google Scholar] [CrossRef] [Green Version]
- Bidou, L.; Bugaud, O.; Belakhov, V.; Baasov, T.; Namy, O. Characterization of new-generation aminoglycoside promoting premature termination codon readthrough in cancer cells. RNA Biol. 2017, 14, 378–388. [Google Scholar] [CrossRef] [Green Version]
- Jospe-Kaufman, M.; Siomin, L.; Fridman, M. The relationship between the structure and toxicity of aminoglycoside antibiotics. Bioorg. Med. Chem. Lett. 2020, 30, 127218. [Google Scholar] [CrossRef]
- Wangen, J.R.; Green, R. Stop codon context influences genome-wide stimulation of termination codon readthrough by aminoglycosides. Elife 2020, 9, e52611. [Google Scholar] [CrossRef] [PubMed]
- Crawford, D.K.; Alroy, I.; Sharpe, N.; Goddeeris, M.M.; Williams, G. ELX-02 generates protein via premature stop codon read-through without inducing native stop codon read-through proteins. J. Pharmacol. Exp. Ther. 2020, 374, 264–272. [Google Scholar] [CrossRef]
- Michorowska, S. Ataluren—Promising Therapeutic Premature Termination Codon Readthrough Frontrunner. Pharmaceuticals 2021, 14, 785. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.; Du, M.; Wong, E.; Mutyam, V.; Li, Y.; Chen, J.; Wangen, J.; Thrasher, K.; Fu, L.; Peng, N.; et al. A small molecule that induces translational readthrough of CFTR nonsense mutations by eRF1 depletion. Nat. Commun. 2021, 12, 4358. [Google Scholar] [CrossRef] [PubMed]
- Baradaran-Heravi, A.; Balgi, A.D.; Hosseini-Farahabadi, S.; Choi, K.; Has, C.; Roberge, M. Effect of small molecule eRF3 degraders on premature termination codon readthrough. Nucleic Acids Res. 2021, 49, 3692–3708. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.E.; Lewis, C.A.; He, L.; Bulik-Sullivan, E.C.; Gallant, S.C.; Mascenik, T.M.; Dang, H.; Cholon, D.M.; Gentzsch, M.; Morton, L.C.; et al. Small-molecule eRF3a degraders rescue CFTR nonsense mutations by promoting premature termination codon readthrough. J. Clin. Investig. 2022, 132, e154571. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Roy, B.; Friesen, W.J.; Tomizawa, Y.; Leszyk, J.D.; Zhuo, J.; Johnson, B.; Dakka, J.; Trotta, C.R.; Xue, X.; Mutyam, V.; et al. Ataluren stimulates ribosomal selection of near-cognate tRNAs to promote nonsense suppression. Proc. Natl. Acad. Sci. USA 2016, 113, 12508–12513. [Google Scholar] [CrossRef] [Green Version]
- Kessel, D.; Morgan, A.; Garbo, G.M. Sites and efficacy of photodamage by tin etiopurpurin in vitro using different delivery systems. Photochem. Photobiol. 1991, 54, 193–196. [Google Scholar] [CrossRef]
- Bidou, L.; Bugaud, O.; Merer, G.; Coupet, M.; Hatin, I.; Chirkin, E.; Karri, S.; Demais, S.; François, P.; Cintrat, J.C.; et al. 2-Guanidino-quinazoline promotes the readthrough of nonsense mutations underlying human genetic diseases. Proc. Natl. Acad. Sci. USA 2022, 119, e2122004119. [Google Scholar] [CrossRef]
- Morrill, C.; Friesen, W.J.; Babu, S.; Baiazitov, R.Y.; Du, W.; Karloff, D.B.; Lee, C.S.; Moon, Y.C.; Ren, H.; Sierra, J.; et al. Guanidino quinazolines and pyrimidines promote readthrough of premature termination codons in cells with native nonsense mutations. Bioorg. Med. Chem. Lett. 2022, 76, 128989. [Google Scholar] [CrossRef]
- Dabrowski, M.; Bukowy-Bieryllo, Z.; Zietkiewicz, E. Translational readthrough potential of natural termination codons in eucaryotes--The impact of RNA sequence. RNA Biol. 2015, 12, 950–958. [Google Scholar] [CrossRef] [PubMed]
- Cridge, A.G.; Crowe-McAuliffe, C.; Mathew, S.F.; Tate, W.P. Eukaryotic translational termination efficiency is influenced by the 3′ nucleotides within the ribosomal mRNA channel. Nucleic Acids Res. 2018, 46, 1927–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tate, W.P.; Cridge, A.G.; Brown, C.M. ‘Stop’in protein synthesis is modulated with exquisite subtlety by an extended RNA translation signal. Biochem. Soc. Trans. 2018, 46, 1615–1625. [Google Scholar] [CrossRef]
- Palma, M.; Lejeune, F. Deciphering the molecular mechanism of stop codon readthrough. Biol. Rev. 2021, 96, 310–329. [Google Scholar] [CrossRef] [PubMed]
- Biziaev, N.; Sokolova, E.; Yanvarev, D.V.; Toropygin, I.Y.; Shuvalov, A.; Egorova, T.; Alkalaeva, E. Recognition of 3′ nucleotide context and stop codon readthrough are determined during mRNA translation elongation. J. Biol. Chem. 2022, 298, 102133. [Google Scholar] [CrossRef]
- Balic, Z.; Hubmacher, D. Promoting translational readthrough to augment fibrillin-1 (FBN1) deposition in Marfan syndrome fibroblasts: A proof-of-concept study. bioRxiv 2022. [Google Scholar] [CrossRef]
- Welcome to the CFTR2. Available online: https://cftr2.org/mutations_history (accessed on 15 May 2022).
- Lashkevich, K.A.; Shlyk, V.I.; Kushchenko, A.S.; Gladyshev, V.N.; Alkalaeva, E.Z.; Dmitriev, S.E. CTELS: A cell-free system for the analysis of translation termination rate. Biomolecules 2020, 10, 911. [Google Scholar] [CrossRef]
- Susorov, D.; Egri, S.; Korostelev, A.A. Termi-Luc: A versatile assay to monitor full-protein release from ribosomes. RNA 2020, 26, 2044–2050. [Google Scholar] [CrossRef]
- Loughran, G.; Howard, M.T.; Firth, A.E.; Atkins, J.F. Avoidance of reporter assay distortions from fused dual reporters. RNA 2017, 23, 1285–1289. [Google Scholar] [CrossRef] [Green Version]
- Ko, W.; Porter, J.J.; Sipple, M.T.; Edwards, K.M.; Lueck, J.D. Efficient suppression of endogenous CFTR nonsense mutations using anticodon-engineered transfer RNAs. Mol. Ther. Nucleic Acids 2022, 28, 685–701. [Google Scholar] [CrossRef]
- Lueck, J.D.; Yoon, J.S.; Perales-Puchalt, A.; Mackey, A.L.; Infield, D.T.; Behlke, M.A.; Pope, M.R.; Weiner, D.B.; Skach, W.R.; McCray, P.B.; et al. Engineered transfer RNAs for suppression of premature termination codons. Nat. Commun. 2019, 10, 1. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 80S Ribosome Preparation a | Fluorescent Label | Relative Peptide/80S b |
|---|---|---|
| 40S + 60S | None | 1.00 c |
| Atto647 | 0.90 ± 0.10 | |
| KCl-treated 80S | None | 0.56 ± 0.07 |
| Atto647 | 0.56 ± 0.12 | |
| Alexa 488 | 0.53 ± 0.03 | |
| Cy3 | 0.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghelfi, M.D.; Bhat, S.Y.; Li, H.; Cooperman, B.S. A High-Throughput Assay for In Vitro Determination of Release Factor-Dependent Peptide Release from a Pretermination Complex by Fluorescence Anisotropy—Application to Nonsense Suppressor Screening and Mechanistic Studies. Biomolecules 2023, 13, 242. https://doi.org/10.3390/biom13020242
Ghelfi MD, Bhat SY, Li H, Cooperman BS. A High-Throughput Assay for In Vitro Determination of Release Factor-Dependent Peptide Release from a Pretermination Complex by Fluorescence Anisotropy—Application to Nonsense Suppressor Screening and Mechanistic Studies. Biomolecules. 2023; 13(2):242. https://doi.org/10.3390/biom13020242
Chicago/Turabian StyleGhelfi, Mikel D., Saleem Y. Bhat, Hong Li, and Barry S. Cooperman. 2023. "A High-Throughput Assay for In Vitro Determination of Release Factor-Dependent Peptide Release from a Pretermination Complex by Fluorescence Anisotropy—Application to Nonsense Suppressor Screening and Mechanistic Studies" Biomolecules 13, no. 2: 242. https://doi.org/10.3390/biom13020242
APA StyleGhelfi, M. D., Bhat, S. Y., Li, H., & Cooperman, B. S. (2023). A High-Throughput Assay for In Vitro Determination of Release Factor-Dependent Peptide Release from a Pretermination Complex by Fluorescence Anisotropy—Application to Nonsense Suppressor Screening and Mechanistic Studies. Biomolecules, 13(2), 242. https://doi.org/10.3390/biom13020242