

Escin’s Multifaceted Therapeutic Profile in Treatment and Post-Treatment of Various Cancers: A Comprehensive Review
 ,
,  and
and
Abstract
:1. Introduction
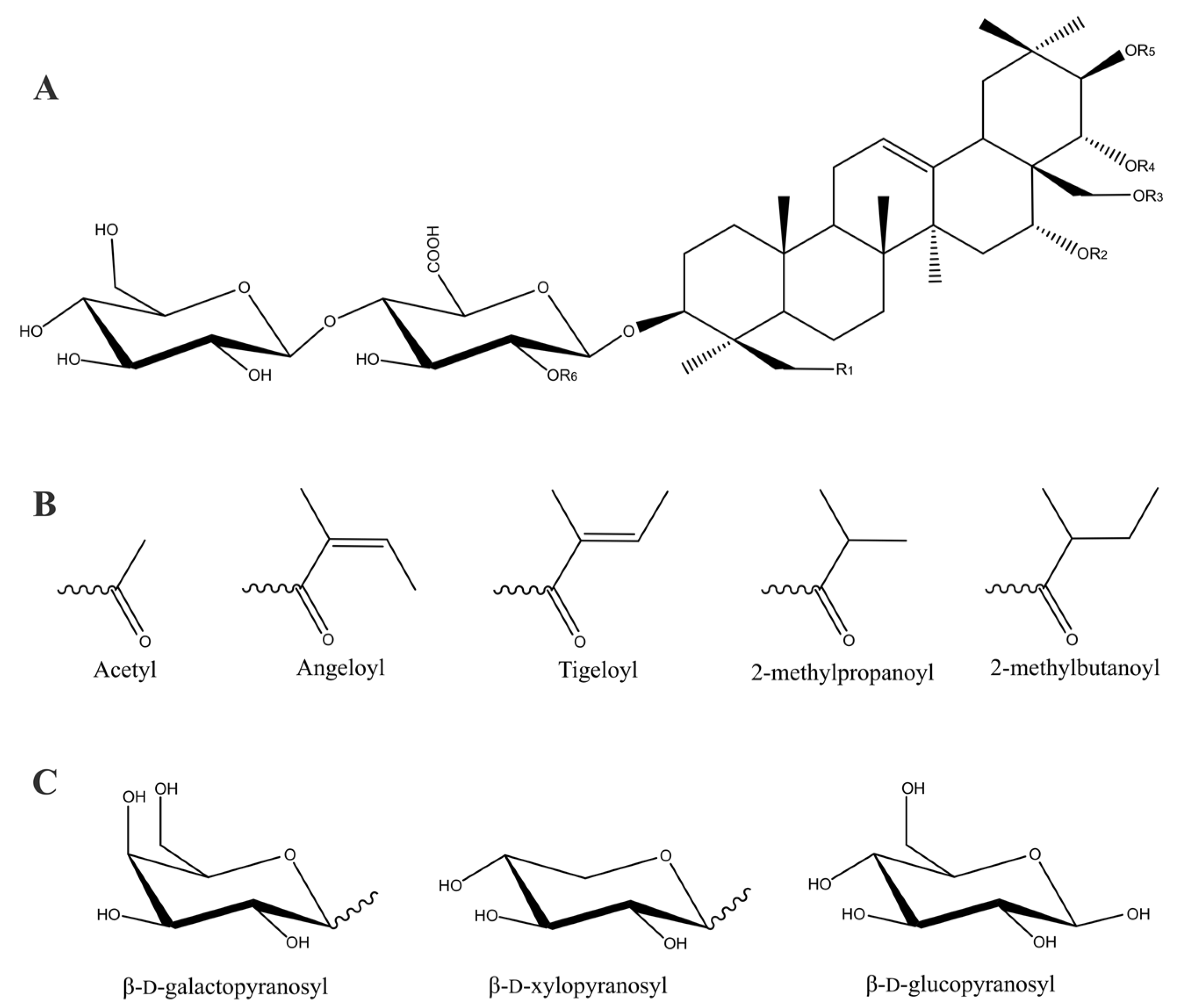
2. Chemistry of Escin
3. Pharmacokinetics and Bioavailability of Escin
4. Escin’s Relevant Biological Activities in Cancer Therapy
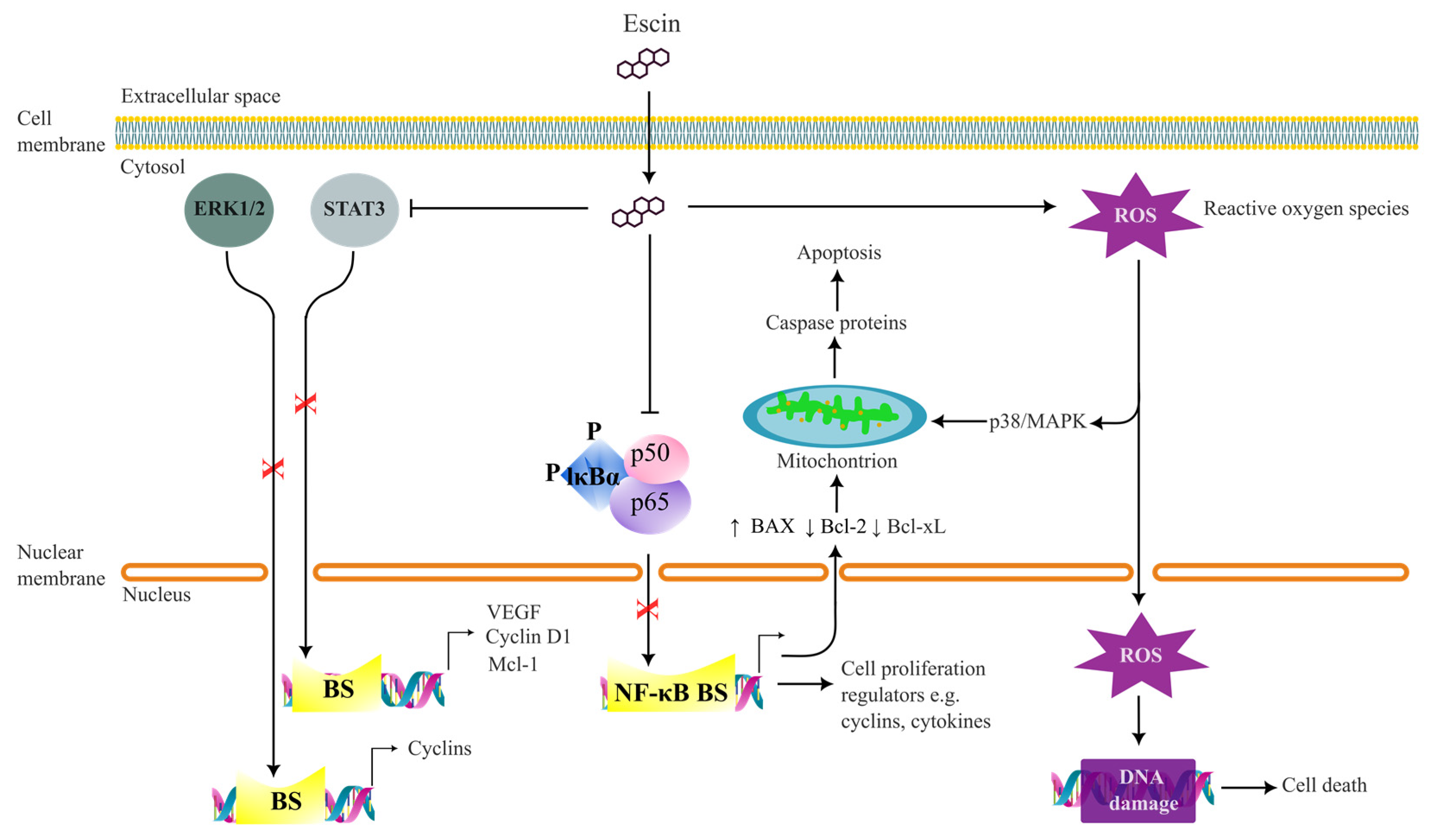
4.1. Anti-Cancer Effects
4.2. Anti-Angiogenetic Effects
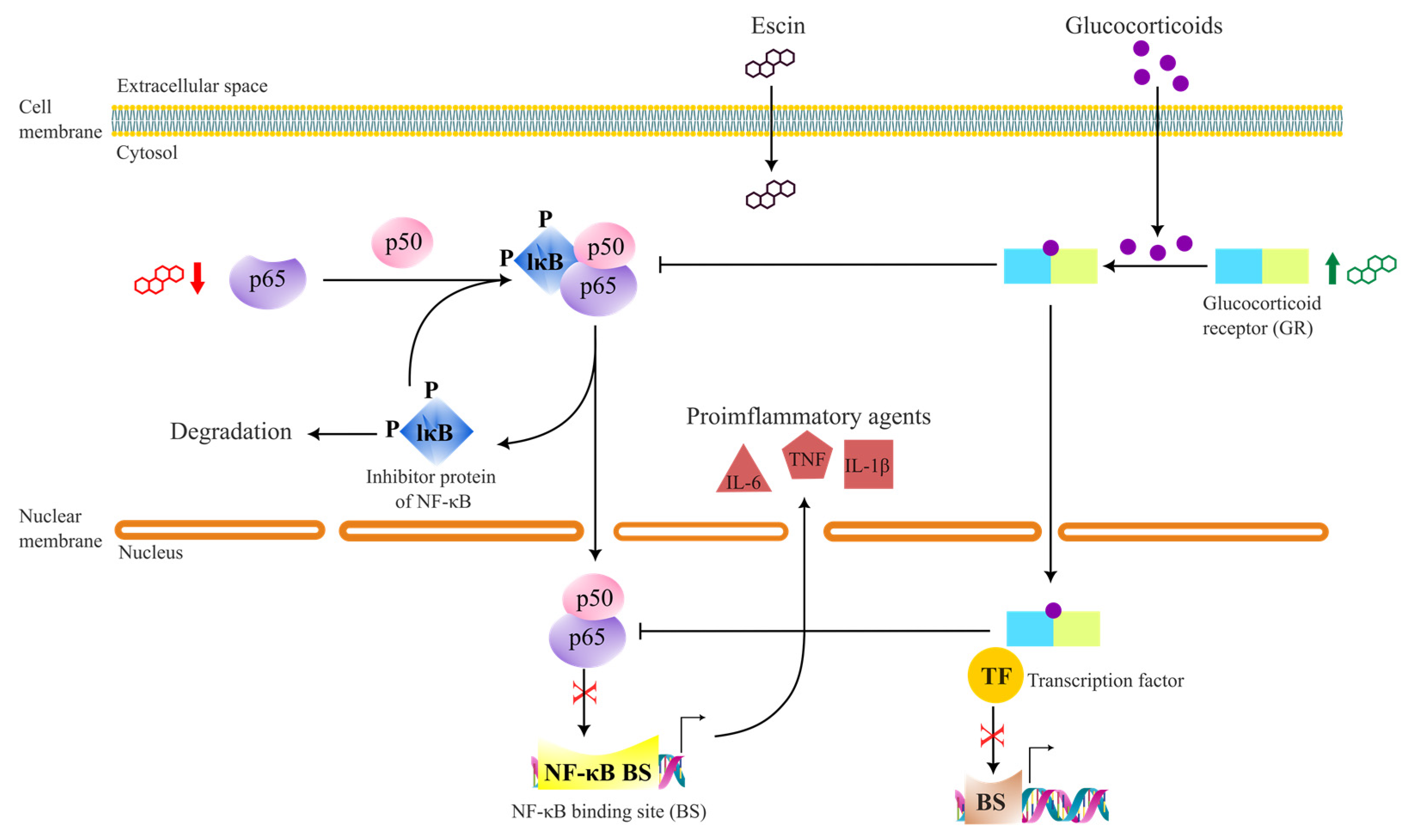
4.3. Anti-Inflammatory Effects
4.4. Antioxidant, Protective and Ameliorative Effects
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Augostine, C.R.; Avery, S.V. Discovery of Natural Products with Antifungal Potential Through Combinatorial Synergy. Front. Microbiol. 2022, 13, 866840. [Google Scholar] [CrossRef] [PubMed]
- Urbina, F.; Lentzos, F.; Invernizzi, C.; Ekins, S. Dual use of artificial-intelligence-powered drug discovery. Nat. Mach. Intell. 2022, 4, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- World Heath Organisation, Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 2 February 2023).
- Sharipov, A.; Tursunov, K.; Fazliev, S.; Azimova, B.; Razzokov, J. Hypoglycemic and Anti-Inflammatory Effects of Triterpene Glycoside Fractions from Aeculus hippocastanum Seeds. Molecules 2021, 26, 3784. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, S.; Yang, Y.; Wang, D.; Gao, H. Natural barrigenol-like triterpenoids: A comprehensive review of their contributions to medicinal chemistry. Phytochemistry 2019, 161, 41–74. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, L.; Yuan, M.; Yu, Q.; Fu, F. Anti-Inflammatory and Gastroprotective Effects of Escin. Nat. Prod. Commun. 2020, 15, 1–10. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Harada, E.; Murakami, T.; Matsuda, H.; Wariishi, N.; Yamahara, J.; Murakami, N.; Kitagawa, I. Escins-Ia, Ib, IIa, IIb, and IIIa, bioactive triterpene oligoglycosides from the seeds of Aesculus hippocastanum L.: Their inhibitory effects on ethanol absorption and hypoglycemic activity on glucose tolerance test. Chem. Pharm. Bull. 1994, 42, 1357–1359. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Murakami, T.; Matsuda, H.; Yamahara, J.; Murakami, N.; Kitagawa, I. Bioactive saponins and glycosides. III. Horse chestnut. (1): The structures, inhibitory effects on ethanol absorption, and hypoglycemic activity of escins Ia, Ib, IIa, IIb, and IIIa from the seeds of Aesculus hippocastanum L. Chem. Pharm. Bull. 1996, 44, 1454–1464. [Google Scholar] [CrossRef]
- Matsuda, H.; Li, Y.; Murakami, T.; Ninomiya, K.; Yamahara, J.; Yoshikawa, M. Effects of escins Ia, Ib, IIa, and IIb from horse chestnut, the seeds of Aesculus hippocastanum L., on acute inflammation in animals. Biol. Pharm. Bull. 1997, 20, 1092–1095. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Murakami, T.; Yamahara, J.; Matsuda, H. Bioactive saponins and glycosides. XII. Horse chestnut. (2): Structures of escins IIIb, IV, V, and VI and isoescins Ia, Ib, and V, acylated polyhydroxyoleanene triterpene oligoglycosides, from the seeds of horse chestnut tree (Aesculus hippocastanum L., Hippocastanaceae). Chem. Pharm. Bull. 1998, 46, 1764–1769. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, M.; Murakami, T.; Otuki, K.; Yamahara, J.; Matsuda, H. Bioactive saponins and glycosides. XIII. Horse chestnut. (3): Quantitative analysis of escins Ia, Ib, IIa, and IIb by means of high performance liquid chromatography. Yakugaku Zasshi 1999, 119, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Assessment Report on Aesculus hippocastanum L., semen; European Medicines Agency: Amsterdam, The Netherlands, 2020.
- Yang, X.W.; Zhao, J.; Cui, Y.X.; Liu, X.H.; Ma, C.M.; Hattori, M.; Zhang, L.H.; Zhang, L.H. Anti-HIV-1 protease triterpenoid saponins from the seeds of Aesculus chinensis. J. Nat. Prod. 1999, 11, 1510–1513. [Google Scholar] [CrossRef] [PubMed]
- Jing Zhao, X.-W.Y.M.H. Three New Triterpene Saponins from the Seeds of Aesculus chinensis. Chem. Pharm. Bull. 2001, 49, 626–628. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yang, X.-W. Four new triterpene saponins from the seeds of Aesculus chinensis. J. Asian Nat. Prod. Res. 2003, 5, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-W.; Zhao, J.; Hattori, M. Three new triterpenoid saponins from the seeds of Aesculus turbinata. J. Asian Nat. Prod. Res. 2008, 10, 243–247. [Google Scholar] [CrossRef]
- Kimura, H.; Jisaka, M.; Kimura, Y.; Katsube, T.; Yokota, K. Attenuating Effect of Saponins Isolated from Edible Horse Chestnuts (Aesculus turbinata BL.) on Increasing Blood Glucose Levels and Their Reducing Bitter Taste. J. Jpn. Soc. Food Sci. Technol.-Nippon Shokuhin Kagaku Kogaku Kaishi-J. Jpn. Soc. Food Sci. Technol. 2006, 53, 31–38. [Google Scholar] [CrossRef]
- Kimura, H.; Watanabe, A.; Jisaka, M.; Yamamoto, T.; Kimura, Y.; Katsube, T.; Yokota, K. Chemical Structures of Saponins from Seeds of Edible Horse Chestnuts (Aesculus turbinata) after Treatment with Wooden Ashes and Their Hypoglycemic Activity. Nippon Shokuhin Kagaku Kogaku Kaishi 2004, 51, 672–679. [Google Scholar] [CrossRef]
- Wu, X.J.; Zhang, M.L.; Cui, X.Y.; Gao, F.; He, Q.; Li, X.J.; Zhang, J.W.; Fawcett, J.P.; Gu, J.K. Comparative pharmacokinetics and bioavailability of escin Ia and isoescin Ia after administration of escin and of pure escin Ia and isoescin Ia in rat. J. Ethnopharmacol. 2012, 139, 201–206. [Google Scholar] [CrossRef]
- Wu, X.J.; Zhang, M.L.; Cui, X.Y.; Paul Fawcett, J.; Gu, J.K. Comparative pharmacokinetics and the bioavailability of escin Ib and isoescin Ib following the administration of escin, pure escin Ib and isoescin Ib in rats. J. Ethnopharmacol. 2014, 151, 839–845. [Google Scholar] [CrossRef]
- Oschmann, R.; Biber, A.; Lang, F.; Stumpf, H.; Kunz, K. Pharmaokinetics of beta-escin after administration of various Aesculus extract containing formulations. Pharmazie 1996, 51, 577–581. [Google Scholar]
- Kunz, K.; Lorkowski, G.; Petersen, G.; Samcova, E.; Schaffler, K.; Wauschkuhn, C.H. Bioavailability of escin after administration of two oral formulations containing Aesculus extract. Arzneimittel-Forschung-Drug Res. 1998, 48, 822–825. [Google Scholar]
- Bässler, D.; Okpanyi, S.; Schrödter, A.; Loew, D.; Schürer, M.; Schulz, H.-U. Bioavailability of β-aescin from horse chestnut seed extract: Comparative clinical studies of two galenic formulations. Adv. Ther. 2003, 20, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Sirtori, C.R. Aescin: Pharmacology, pharmacokinetics and therapeutic profile. Pharm. Res 2001, 44, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Kobryn, J.; Zieba, T.; Sowa, S.K.; Musial, W. Influence of Acetylated Annealed Starch on the Release of beta-Escin from the Anionic and Non-Ionic Hydrophilic Gels. Pharmaceutics 2020, 12, 84. [Google Scholar] [CrossRef] [PubMed]
- Kobryń, J.; Sowa, S.; Gasztych, M.; Dryś, A.; Musiał, W. Influence of Hydrophilic Polymers on theβFactor in Weibull Equation Applied to the Release Kinetics of a Biologically Active Complex ofAesculus hippocastanum. Int. J. Polym. Sci. 2017, 2017, 3486384. [Google Scholar] [CrossRef]
- Huang, S.; Wang, X.; Liu, M.; Lin, Z.; Gu, W.; Zhao, H.; Zhang, Y.; Ding, B.; Liu, J.; Wu, X.; et al. Modification of sodium aescinate into a safer, more stable and effective water-soluble drug by liposome-encapsulation: An in vitro and in vivo study. Drug Deliv. 2022, 29, 1132–1141. [Google Scholar] [CrossRef]
- Cheng, C.L.; Chao, W.T.; Li, Y.H.; Ou, Y.C.; Wang, S.S.; Chiu, K.Y.; Yuan, S.Y. Escin induces apoptosis in human bladder cancer cells: An in vitro and in vivo study. Eur. J Pharm. 2018, 840, 79–88. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, X.; Zhao, P.; Tong, B.; Wei, Z.; Dai, Y. Escin Ia suppresses the metastasis of triple-negative breast cancer by inhibiting epithelial-mesenchymal transition via down-regulating LOXL2 expression. Oncotarget 2016, 7, 23684–23699. [Google Scholar] [CrossRef]
- Akar, S.; Donmez-Altuntas, H.; Hamurcu, Z. β-Escin reduces cancer progression in aggressive MDA-MB-231 cells by inhibiting glutamine metabolism through downregulation of c-myc oncogene. Mol. Biol. Rep. 2022, 49, 7409–7415. [Google Scholar] [CrossRef]
- Mojzisova, G.; Mojzis, J.; Pilatova, M.; Varinska, L.; Ivanova, L.; Strojny, L.; Richnavsky, J. Antiproliferative and antiangiogenic properties of horse chestnut extract. Phytother. Res. 2013, 27, 159–165. [Google Scholar] [CrossRef]
- Mojzisova, G.; Kello, M.; Pilatova, M.; Tomeckova, V.; Vaskova, J.; Vasko, L.; Bernatova, S.; Mirossay, L.; Mojzis, J. Antiproliferative effect of beta-escin—An in vitro study. Acta Biochim. Pol. 2016, 63, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Mazrouei, R.; Raeisi, E.; Lemoigne, Y.; Heidarian, E. Activation of p53 Gene Expression and Synergistic Antiproliferative Effects of 5-Fluorouracil and beta-escin on MCF7 Cells. J. Med. Signals Sens. 2019, 9, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Long, L.; Zhang, X.; Song, K.; Wang, D.; Xiong, X.; Gao, H.; Sha, L. 16-Tigloyl linked barrigenol-like triterpenoid from Semen Aesculi and its anti-tumor activity in vivo and in vitro. RSC Adv. 2019, 9, 31758–31772. [Google Scholar] [CrossRef] [PubMed]
- Patlolla, J.M.; Raju, J.; Swamy, M.V.; Rao, C.V. Beta-escin inhibits colonic aberrant crypt foci formation in rats and regulates the cell cycle growth by inducing p21(waf1/cip1) in colon cancer cells. Mol. Cancer 2006, 5, 1459–1466. [Google Scholar] [CrossRef]
- Seweryn, E.; Glehsk, M.; Środa-Pomianek, K.; Ceremuga, I.; Włodarczyk, M.; Gamian, A. Cytotoxic Effects of Four Aescin Types on Human Colon Adenocarcinoma Cell Lines. Nat. Prod. Commun. 2014, 9, 387–390. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Q.; Li, B.; Xie, J.-M.; Yang, X.-D.; Zhao, K.; Wu, Y.; Ye, Z.-Y.; Chen, Z.-R.; Qin, Z.-H.; et al. Escin-induced DNA damage promotes escin-induced apoptosis in human colorectal cancer cells via p62 regulation of the ATM/γH2AX pathway. Acta Pharmacol. Sin. 2018, 39, 1645–1660. [Google Scholar] [CrossRef]
- Li, B.; Wang, Z.; Xie, J.M.; Wang, G.; Qian, L.Q.; Guan, X.M.; Shen, X.P.; Qin, Z.H.; Shen, G.H.; Li, X.Q.; et al. TIGAR knockdown enhanced the anticancer effect of aescin via regulating autophagy and apoptosis in colorectal cancer cells. Acta Pharm. Sin. 2019, 40, 111–121. [Google Scholar] [CrossRef]
- Shen, D.Y.; Kang, J.H.; Song, W.; Zhang, W.Q.; Li, W.G.; Zhao, Y.; Chen, Q.X. Apoptosis of human cholangiocarcinoma cell lines induced by beta-escin through mitochondrial caspase-dependent pathway. Phytother. Res. 2011, 25, 1519–1526. [Google Scholar] [CrossRef]
- Huang, G.L.; Shen, D.Y.; Cai, C.F.; Zhang, Q.Y.; Ren, H.Y.; Chen, Q.X. beta-escin reverses multidrug resistance through inhibition of the GSK3beta/beta-catenin pathway in cholangiocarcinoma. World J. Gastroenterol. 2015, 21, 1148–1157. [Google Scholar] [CrossRef]
- Lee, H.S.; Hong, J.E.; Kim, E.J.; Kim, S.H. Escin suppresses migration and invasion involving the alteration of CXCL16/CXCR6 axis in human gastric adenocarcinoma AGS cells. Nutr. Cancer 2014, 66, 938–945. [Google Scholar] [CrossRef]
- Harford-Wright, E.; Bidère, N.; Gavard, J. β-escin selectively targets the glioblastoma-initiating cell population and reduces cell viability. Oncotarget 2016, 7, 66865–66879. [Google Scholar] [CrossRef] [PubMed]
- Ming, Z.J.; Hu, Y.; Qiu, Y.H.; Cao, L.; Zhang, X.G. Synergistic effects of β-aescin and 5-fluorouracil in human hepatocellular carcinoma SMMC-7721 cells. Phytomedicine 2010, 17, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.Y.; Fu, F.H.; Li, Z.; Dong, Q.J.; He, J.; Wang, C.H. Escin, a natural mixture of triterpene saponins, exhibits antitumor activity against hepatocellular carcinoma. Planta Med. 2009, 75, 1580–1585. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.M.-L.; Li, F.; Rajendran, P.; Kumar, A.P.; Hui, K.M.; Sethi, G. Identification of β-Escin as a Novel Inhibitor of Signal Transducer and Activator of Transcription 3/Janus-Activated Kinase 2 Signaling Pathway that Suppresses Proliferation and Induces Apoptosis in Human Hepatocellular Carcinoma Cells. J. Pharmacol. Exp. Ther. 2010, 334, 285. [Google Scholar] [CrossRef]
- Zhang, Z.; Gao, J.; Cai, X.; Zhao, Y.; Wang, Y.; Lu, W.; Gu, Z.; Zhang, S.; Cao, P. Escin sodium induces apoptosis of human acute leukemia Jurkat T cells. Phytother. Res. 2011, 25, 1747–1755. [Google Scholar] [CrossRef] [PubMed]
- Harikumar, K.B.; Sung, B.; Pandey, M.K.; Guha, S.; Krishnan, S.; Aggarwal, B.B. Escin, a Pentacyclic Triterpene, Chemosensitizes Human Tumor Cells through Inhibition of Nuclear Factor-κB Signaling Pathway. Mol. Pharmacol. 2010, 77, 818. [Google Scholar] [CrossRef]
- Ji, D.B.; Xu, B.; Liu, J.T.; Ran, F.X.; Cui, J.R. beta-Escin sodium inhibits inducible nitric oxide synthase expression via downregulation of the JAK/STAT pathway in A549 cells. Mol. Carcinog. 2011, 50, 945–960. [Google Scholar] [CrossRef]
- Patlolla, J.M.; Qian, L.; Biddick, L.; Zhang, Y.; Desai, D.; Amin, S.; Lightfoot, S.; Rao, C.V. beta-Escin inhibits NNK-induced lung adenocarcinoma and ALDH1A1 and RhoA/Rock expression in A/J mice and growth of H460 human lung cancer cells. Cancer Prev. Res. 2013, 6, 1140–1149. [Google Scholar] [CrossRef]
- Ciftci, G.A.; Iscan, A.; Kutlu, M. Escin reduces cell proliferation and induces apoptosis on glioma and lung adenocarcinoma cell lines. Cytotechnology 2015, 67, 893–904. [Google Scholar] [CrossRef]
- Kwak, H.; An, H.; Alam, M.B.; Choi, W.-S.; Lee, S.Y.; Lee, S.-H. Inhibition of Migration and Invasion in Melanoma Cells by β-Escin via the ERK/NF-κB Signaling Pathway. Biol. Pharm. Bull. 2018, 41, 1606–1610. [Google Scholar] [CrossRef]
- Zhu, J.; Yu, W.; Liu, B.; Wang, Y.; Shao, J.; Wang, J.; Xia, K.; Liang, C.; Fang, W.; Zhou, C.; et al. Escin induces caspase-dependent apoptosis and autophagy through the ROS/p38 MAPK signalling pathway in human osteosarcoma cells in vitro and in vivo. Cell Death Dis. 2017, 8, e3113. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Ying, J.; Lin, C.; Wang, Y.; Huang, K.; Zhou, Y.; Teng, H. beta-Escin inhibits the proliferation of osteosarcoma cells via blocking the PI3K/Akt pathway. RSC Adv. 2018, 8, 29637–29644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenny, H.A.; Hart, P.C.; Kordylewicz, K.; Lal, M.; Shen, M.; Kara, B.; Chen, Y.J.; Grassl, N.; Alharbi, Y.; Pattnaik, B.R.; et al. The Natural Product beta-Escin Targets Cancer and Stromal Cells of the Tumor Microenvironment to Inhibit Ovarian Cancer Metastasis. Cancers 2021, 13, 3931. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.W.; Wang, S.J.; Zhou, Y.N.; Pan, S.H.; Sun, B. Escin augments the efficacy of gemcitabine through down-regulation of nuclear factor-kappaB and nuclear factor-kappaB-regulated gene products in pancreatic cancer both in vitro and in vivo. J Cancer Res. Clin. Oncol. 2012, 138, 785–797. [Google Scholar] [CrossRef]
- Rimmon, A.; Vexler, A.; Berkovich, L.; Earon, G.; Ron, I.; Lev-Ari, S. Escin Chemosensitizes Human Pancreatic Cancer Cells and Inhibits the Nuclear Factor-kappaB Signaling Pathway. Biochem. Res. Int. 2013, 2013, 251752. [Google Scholar] [CrossRef]
- Omi, K.; Matsuo, Y.; Ueda, G.; Aoyama, Y.; Kato, T.; Hayashi, Y.; Imafuji, H.; Saito, K.; Tsuboi, K.; Morimoto, M.; et al. Escin inhibits angiogenesis by suppressing interleukin8 and vascular endothelial growth factor production by blocking nuclear factorkappaB activation in pancreatic cancer cell lines. Oncol. Rep. 2021, 45, 55. [Google Scholar] [CrossRef]
- Piao, S.; Kang, M.; Lee, Y.J.; Choi, W.S.; Chun, Y.S.; Kwak, C.; Kim, H.H. Cytotoxic effects of escin on human castration-resistant prostate cancer cells through the induction of apoptosis and G2/M cell cycle arrest. Urology 2014, 84, 982.e1–982.e7. [Google Scholar] [CrossRef]
- Yuan, S.Y.; Cheng, C.L.; Wang, S.S.; Ho, H.C.; Chiu, K.Y.; Chen, C.S.; Chen, C.C.; Shiau, M.Y.; Ou, Y.C. Escin induces apoptosis in human renal cancer cells through G2/M arrest and reactive oxygen species-modulated mitochondrial pathways. Oncol. Rep. 2017, 37, 1002–1010. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef]
- Luo, J.-L.; Kamata, H.; Karin, M. IKK/NF-κB signaling: Balancing life and death—A new approach to cancer therapy. J. Clin. Investig. 2005, 115, 2625–2632. [Google Scholar] [CrossRef]
- Taniguchi, K.; Karin, M. NF-kappaB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Verma, I.M. NF-kappaB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Paneerselvam, C.; Ganapasam, S. beta-Escin alleviates cobalt chloride-induced hypoxia-mediated apoptotic resistance and invasion via ROS-dependent HIF-1alpha/TGF-beta/MMPs in A549 cells. Toxicol. Res. 2020, 9, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Varinska, L.; Faber, L.; Kello, M.; Petrovova, E.; Balazova, L.; Solar, P.; Coma, M.; Urdzik, P.; Mojzis, J.; Svajdlenka, E.; et al. beta-Escin Effectively Modulates HUVECS Proliferation and Tube Formation. Molecules 2018, 23, 197. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Xu, B.; Liu, J.T.; Cui, J.R. Effect of beta-escin sodium on endothelial cells proliferation, migration and apoptosis. Vasc. Pharm. 2008, 49, 158–165. [Google Scholar] [CrossRef]
- Cheng, P.; Kuang, F.; Ju, G. Aescin reduces oxidative stress and provides neuroprotection in experimental traumatic spinal cord injury. Free Radic. Biol. Med. 2016, 99, 405–417. [Google Scholar] [CrossRef]
- Diehm, C.; Trampisch, H.J.; Lange, S.; Schmidt, C. Comparison of leg compression stocking and oral horse-chestnut seed extract therapy in patients with chronic venous insufficiency. Lancet 1996, 347, 292–294. [Google Scholar] [CrossRef]
- Dudek-Makuch, M.; Studzińska-Sroka, E. Horse chestnut—Efficacy and safety in chronic venous insufficiency: An overview. Rev. Bras. Farmacogn. 2015, 25, 533–541. [Google Scholar] [CrossRef]
- Vogel, G.; Marek, M.L.; Stoeckert, I. Weitere Untersuchengen zun Wirkungsmechanismus des Rosskastanien-Saponins Aescin. Arzneim. Forsch. 1963, 13, 59–64. [Google Scholar]
- Xin, W.; Zhang, L.; Fan, H.; Jiang, N.; Wang, T.; Fu, F. Escin attenuates acute lung injury induced by endotoxin in mice. Eur. J. Pharm. Sci. 2011, 42, 73–80. [Google Scholar] [CrossRef]
- Xin, W.; Zhang, L.; Sun, F.; Jiang, N.; Fan, H.; Wang, T.; Li, Z.; He, J.; Fu, F. Escin exerts synergistic anti-inflammatory effects with low doses of glucocorticoids in vivo and in vitro. Phytomedicine 2011, 18, 272–277. [Google Scholar] [CrossRef]
- Liu, S.; Wang, H.; Qiu, C.; Zhang, J.; Zhang, T.; Zhou, W.; Lu, Z.; Rausch-Fan, X.; Liu, Z. Escin inhibits lipopolysaccharide-induced inflammation in human periodontal ligament cells. Mol. Med. Rep. 2012, 6, 1150–1154. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, L.; Jiang, N.; Wang, Z.; Chong, Y.; Fu, F. Anti-inflammatory effects of escin are correlated with the glucocorticoid receptor/NF-kappaB signaling pathway, but not the COX/PGF2alpha signaling pathway. Exp. Med. 2013, 6, 419–422. [Google Scholar] [CrossRef]
- Gupta, S.C.; Tyagi, A.K.; Deshmukh-Taskar, P.; Hinojosa, M.; Prasad, S.; Aggarwal, B.B. Downregulation of tumor necrosis factor and other proinflammatory biomarkers by polyphenols. Arch Biochem Biophys 2014, 559, 91–99. [Google Scholar] [CrossRef]
- Zhao, S.-Q.; Xu, S.-Q.; Cheng, J.; Cao, X.-L.; Zhang, Y.; Zhou, W.-P.; Huang, Y.-J.; Wang, J.; Hu, X.-M. Anti-inflammatory effect of external use of escin on cutaneous inflammation: Possible involvement of glucocorticoids receptor. Chin. J. Nat. Med. 2018, 16, 105–112. [Google Scholar] [CrossRef]
- Hiai, S.; Yokoyama, H.; Oura, H. Effect of escin on adrenocorticotropin and corticosterone levels in rat plasma. Chem. Pharm. Bull. 1981, 29, 490–494. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, H.; Fan, H.; Wang, T.; Jiang, N.; Yu, P.; Fu, F. The potent anti-inflammatory agent escin does not increase corticosterone secretion and immune cell apoptosis in mice. Fitoterapia 2011, 82, 861–867. [Google Scholar] [CrossRef]
- Zhang, F.; Li, Y.; Zhang, L.; Mu, G. Synergistic protective effects of escin and lowdose glucocorticoids on bloodretinal barrier breakdown in a rat model of retinal ischemia. Mol. Med. Rep. 2013, 7, 1511–1515. [Google Scholar] [CrossRef]
- Timmermans, S.; Souffriau, J.; Libert, C. A General Introduction to Glucocorticoid Biology. Front. Immunol. 2019, 10, 1545. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, H.; Mao, M.; Liang, C.; Zhang, Y.; Yang, D.; Wei, Z.; Gao, S.; Hu, B.; Wang, L.; et al. Escin Increases the Survival Rate of LPS-Induced Septic Mice Through Inhibition of HMGB1 Release from Macrophages. Cell Physiol. Biochem. 2015, 36, 1577–1586. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.X.; Eerduna, G.W.; Duan, S.J.; Li, T.; Liu, R.X.; Zhang, L.M.; Wang, T.; Fu, F.H. Escin ameliorates the impairments of neurological function and blood brain barrier by inhibiting systemic inflammation in intracerebral hemorrhagic mice. Exp. Neurol. 2021, 337, 113554. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.E.M.; Ahmed, S.F.; Eltrawy, A.H.; Yousef, R.S.; Ali, H.S.; Mahmoud, A.R.; Abd-Elhamid, T.H. Pretreatment with Coenzyme Q10 Combined with Aescin Protects against Sepsis-Induced Acute Lung Injury. Cells Tissues Organs 2021, 210, 195–217. [Google Scholar] [CrossRef]
- Domanski, D.; Zegrocka-Stendel, O.; Perzanowska, A.; Dutkiewicz, M.; Kowalewska, M.; Grabowska, I.; Maciejko, D.; Fogtman, A.; Dadlez, M.; Koziak, K. Molecular Mechanism for Cellular Response to beta-Escin and Its Therapeutic Implications. PLoS ONE 2016, 11, e0164365. [Google Scholar] [CrossRef] [PubMed]
- Selvakumar, G.P.; Janakiraman, U.; Essa, M.M.; Justin Thenmozhi, A.; Manivasagam, T. Escin attenuates behavioral impairments, oxidative stress and inflammation in a chronic MPTP/probenecid mouse model of Parkinson’s disease. Brain Res. 2014, 1585, 23–36. [Google Scholar] [CrossRef]
- Suryavanshi, S.V.; Kulkarni, Y.A. Attenuation of Cardiac Autonomic Neuropathy by Escin in Diabetic Rats. Pharmacology 2021, 106, 211–217. [Google Scholar] [CrossRef]
- Gur, F.; Cengiz, M.; Kutlu, H.M.; Cengiz, B.P.; Ayhanci, A. Molecular docking analyses of Escin as regards cyclophosphamide-induced cardiotoxicity: In vivo and in Silico studies. Toxicol. Appl. Pharm. 2021, 411, 115386. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Jiang, Y.; Wang, W.; Ma, J.; Chen, M. Escin activates AKT-Nrf2 signaling to protect retinal pigment epithelium cells from oxidative stress. Biochem. Biophys. Res. Commun. 2015, 468, 541–547. [Google Scholar] [CrossRef]
- Hu, X.M.; Zeng, F.D. Protective effects of sodium beta-aescin on ischemia-reperfusion injury in rat brain. Yao Xue Xue Bao 2004, 39, 419–423. [Google Scholar]
- Zhang, L.; Fu, F.; Zhang, X.; Zhu, M.; Wang, T.; Fan, H. Escin attenuates cognitive deficits and hippocampal injury after transient global cerebral ischemia in mice via regulating certain inflammatory genes. Neurochem. Int. 2010, 57, 119–127. [Google Scholar] [CrossRef]
- Selvakumar, G.P.; Manivasagam, T.; Rekha, K.R.; Jayaraj, R.L.; Elangovan, N. Escin, a novel triterpene, mitigates chronic MPTP/p-induced dopaminergic toxicity by attenuating mitochondrial dysfunction, oxidative stress, and apoptosis. J. Mol. Neurosci. 2015, 55, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Jiang, X.; Pan, R.; Zhou, X.; Qin, D.; Xiong, R.; Wang, Y.; Qiu, W.; Wu, A.; Wu, J. Escins Isolated from Aesculus chinensis Bge. Promote the Autophagic Degradation of Mutant Huntingtin and Inhibit its Induced Apoptosis in HT22 cells. Front. Pharm. 2020, 11, 116. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, X.; Wu, L.; Li, Y.; Wang, L.; Zhao, X.; Zhao, T.; Zhang, L.; Yan, Z.; Wei, G. Ameliorative effects of escin on neuropathic pain induced by chronic constriction injury of sciatic nerve. J. Ethnopharmacol. 2021, 267, 113503. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Li, J.; Xu, Q.; Xie, H.; Wang, L.; Zhang, M. Sodium aescinate alleviates bone cancer pain in rats by suppressing microglial activation via p38 MAPK/c-Fos signaling. Mol. Cell. Toxicol. 2022, 18, 605–614. [Google Scholar] [CrossRef]
- Sikorska, M.; Dutkiewicz, M.; Zegrocka-Stendel, O.; Kowalewska, M.; Grabowska, I.; Koziak, K. Beneficial effects of beta-escin on muscle regeneration in rat model of skeletal muscle injury. Phytomedicine 2021, 93, 153791. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Compound | R1 | R2 | R3 | R4 | R5 | R6 | Ref. |
|---|---|---|---|---|---|---|---|
| Escin Ia | OH | H | H | Acetyl | Tigeloyl | β-d-glucopyranosyl | [8,12] |
| Escin Ib | OH | H | H | Acetyl | Angeloyl | β-d-glucopyranosyl | [8,12] |
| Escin IIa | OH | H | H | Acetyl | Tigeloyl | β-d-xylopyranosyl | [8] |
| Escin IIb | OH | H | H | Acetyl | Angeloyl | β-d-xylopyranosyl | [8] |
| Escin IIIa | H | H | H | Acetyl | Tigeloyl | β-d-galactopyranosyl | [8,9] |
| Escin IIIb | H | H | H | Acetyl | Angeloyl | β-d-galactopyranosyl | [11] |
| Escin IV | OH | H | H | Acetyl | Acetyl | β-d-glucopyranosyl | [11] |
| Escin IVc | OH | H | Acetyl | Tigeloyl | H | β-d-glucopyranosyl | [14] |
| Escin IVd | OH | H | Acetyl | Angeloyl | H | β-d-glucopyranosyl | [14] |
| Escin IVe | OH | H | Tigeloyl | H | H | β-d-glucopyranosyl | [14] |
| Escin IVf | OH | H | Angeloyl | H | H | β-d-glucopyranosyl | [14] |
| Escin IVg | OH | H | H | Tigeloyl | H | β-d-glucopyranosyl | [15] |
| Escin IVh | OH | H | H | Angeloyl | H | β-d-glucopyranosyl | [15] |
| Escin V | OH | H | H | Acetyl | 2-methylpropanoyl | β-d-glucopyranosyl | [11] |
| Escin VI | OH | H | H | Acetyl | 2-methylbutanoyl | β-d-glucopyranosyl | [11] |
| Escin VIb | OH | Angeloyl | H | H | Acetyl | β-d-glucopyranosyl | [15] |
| Isoescin Ia | OH | H | Acetyl | H | Tigeloyl | β-d-glucopyranosyl | [11] |
| Isoescin Ib | OH | H | Acetyl | H | Angeloyl | β-d-glucopyranosyl | [11] |
| Isoescin IIa | OH | H | Acetyl | H | Tigeloyl | β-d-xylopyranosyl | [16] |
| Isoescin IIb | OH | H | Acetyl | H | Angeloyl | β-d-xylopyranosyl | [16] |
| Isoescin IIIa | H | H | Acetyl | H | Tigeloyl | β-d-galactopyranosyl | [16] |
| Isoescin IIIb | H | H | Acetyl | H | Angeloyl | β-d-galactopyranosyl | [16] |
| Isoescin V | OH | H | Acetyl | H | 2-methylpropanoyl | β-d-glucopyranosyl | [11] |
| Isoescin VIa | OH | H | Acetyl | H | 2-methylbutanoyl | β-d-glucopyranosyl | [17] |
| Isoescin VIIa | OH | H | Acetyl | H | Tigeloyl | β-d-galactopyranosyl | [17] |
| Isoescin VIIIa | H | H | Acetyl | H | Angeloyl | β-d-glucopyranosyl | [17] |
| Deacylescin I | OH | H | H | H | H | β-d-glucopyranosyl | [8,18] |
| Deacylescin Ia | OH | H | H | H | Tigeloyl | β-d-glucopyranosyl | [19] |
| Deacylescin Ib | OH | H | H | H | Angeloyl | β-d-glucopyranosyl | [19] |
| Deacylescin II | OH | H | H | H | H | β-d-xylopyranosyl | [10,18] |
| Deacylescin IIa | OH | H | H | H | Tigeloyl | β-d-xylopyranosyl | [19] |
| Deacylescin IIb | OH | H | H | H | Angeloyl | β-d-xylopyranosyl | [19] |
| Deacylescin III | H | H | H | H | H | β-d-galactopyranosyl | [8] |
| Cancer Type | Model System | Anti-Cancer Effects and Mechanisms of Escin by | Ref. | ||
|---|---|---|---|---|---|
| Inducing Cell Apoptosis | Decreasing Cell Proliferation | Inhibiting Metastasis and Invasion | |||
| Bladder cancer | T24, J82, TCCSUP, and RT-4 cell lines | Through death receptor and mitochondria mediated pathways | ↓ NF–κB p65, partially affects STAT3 expression | [28] | |
| Breast cancer | MDA-MB-231 cell line | ↓ LOXL2, ↓ c-Myc, ↓ GSL1, ↑ASCT2 | [29,30] | ||
| Mice and MCF-7 cell line | Yes | Yes | ↑ p53, ↓ Bcl-2 | [31,32,33,34] | |
| Colon cancer | HT-29 cell line | Cell cycle arrest at G1/S phase mediated by induction of p21WAF1/CIP1 | [35] | ||
| LoVo cell line | Yes | [36] | |||
| Colorectal cancer | Mice and HCT116, HCT8 cell lines | Through DNA damage | Yes | [37] | |
| Mice and HCT116, HCT8 cell lines | Through DNA damage ↑ TIGAR ↑ ROS | Yes | [38] | ||
| Cholangiocarcinoma | QBC939, MZ-ChA-1 Sk-ChA-1 cell lines | ↑ caspase-3, ↓ Bcl-2 | Cell cycle arrest at G1 and G2/M phases | [39] | |
| QBC939 cell line | Sensitised to 5-FU and VCR | ↓ P-glycoprotein, ↓ GSK3b/ catenin | [40] | ||
| Gastric cancer | AGS cell lines | Through Akt signalling pathway | [41] | ||
| Glioblastoma multiforme | Classical and mesenchymal glioblastoma-initiating cells | Through mitochondria-mediated pathway | Yes | [42] | |
| Hepatocellular carcinoma | SMMC-7721 cell line | ↑ caspases 3, 8, 9; ↓ Bcl-2 with 5-FU | Cell cycle arrest at G0/G1 with 5-FU | [43] | |
| HepG2 cell line | ↑ PARP, AIF, BAX, and Bcl-2 | Cell cycle arrest at G1/S phase | [44] | ||
| HepG2 cell line | ↓ Akt/JAK/STAT, ↓ cyclin D1, ↓ Bcl-2, ↓ Bcl-xL, ↓ survivin, ↓ Mcl-1, ↓ VEGF; sensitised to Dx, PTX | [45] | |||
| Leukaemia | Jurkat T-cell line | ↑ Caspases-3, 8, 9, ↓ PARP, ↓ Bcl-2, ↑ ROS | Yes | [32,46] | |
| CEM cell line | Yes | [32] | |||
| KBM-5 cell line | TNF-induced apoptosis | ↓ TNF ↓ NF–κB | ↓ TNF ↓ NF–κB | [47] | |
| Lung cancer | A549 cell line | Through JAK/STAT signalling pathway | ↓ iNOS | [48] | |
| Mice and H460 cell line | ↓ ALDH1A1 ↓ p-Akt, ↑ p21 | ↓ RhoA and Rock | [49] | ||
| A549 cell line | ↑ BAX, ↑ caspase-3 | Cell cycle arrest at G0/G1phase | [50] | ||
| Melanoma | SK-MEL5 and B16F10 cell lines | ↓ NF–κB, ↓ IκB | Through ERK1/2 signalling | [51] | |
| Osteosarcoma | Mice and MNNG/HOS, Saos-2, MG-63, U-2OS, HUVEC cell lines | Through ROS/p38 MAPK signalling pathway | [52] | ||
| Mice and MG-63, OS732 cell lines | ↑ Caspases-3, 8, 9 | ↓ PI3K/Akt pathway | [53] | ||
| Ovarian cancer | HeyA8, SNU-119, Kuramochi, Ovcar4, and Ovcar5 cell lines | ↓ Autophagy-dependent CSC differentiation, ↓ Stromal ECM production driven by HIF1α | [54] | ||
| Pancreatic cancer | Mice and BxPC-3, PANC-1 cell lines | ↓ NF–κB, ↓ c-Myc, ↓ COX-2, ↓ cyclin D1, ↓ survivin, ↓ Bcl-xL, ↓ Bcl-2, ↑ caspase-3 | [55] | ||
| COLO357, MIA-Paca, Panc-1, cell lines | Yes | ↓ NF–κB, ↓ cyclin D, sensitised cells to cisplatin | [56] | ||
| BxPC-3, AsPC-1, SW1990 cell lines | ↓ NF-κB | ↓ IL-8, ↓ VEGF | [57] | ||
| Prostate cancer | Mice and CRPC, PC-3, DU-145 cell lines | ↑ c-caspase-3, ↑ BAX, ↓ Bcl-2, ↓ cIAP-1, ↓ cIAP-2 ↓ xIAP, ↑ PARP | Cell cycle arrest at G2/M-phase | [58] | |
| Renal cancer | 786-O and Caki-1 cell lines | ↓ Bcl-2, ↑ ROS | Cell cycle arrest at G2/M arrest | [59] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fazliev, S.; Tursunov, K.; Razzokov, J.; Sharipov, A. Escin’s Multifaceted Therapeutic Profile in Treatment and Post-Treatment of Various Cancers: A Comprehensive Review. Biomolecules 2023, 13, 315. https://doi.org/10.3390/biom13020315
Fazliev S, Tursunov K, Razzokov J, Sharipov A. Escin’s Multifaceted Therapeutic Profile in Treatment and Post-Treatment of Various Cancers: A Comprehensive Review. Biomolecules. 2023; 13(2):315. https://doi.org/10.3390/biom13020315
Chicago/Turabian StyleFazliev, Sunnatullo, Khurshid Tursunov, Jamoliddin Razzokov, and Avez Sharipov. 2023. "Escin’s Multifaceted Therapeutic Profile in Treatment and Post-Treatment of Various Cancers: A Comprehensive Review" Biomolecules 13, no. 2: 315. https://doi.org/10.3390/biom13020315