
Intestinal Barrier in Post-Campylobacter jejuni Irritable Bowel Syndrome

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Ethics
2.3. Campylobacter-Specific PCR from Stool and Mucosal Samples
2.4. Electrophysiological Measurements on Colon Biopsies
2.5. Immunohistochemistry of Colonic Epithelium
2.6. Statistics
3. Results
3.1. Patients’ Disease Characteristics
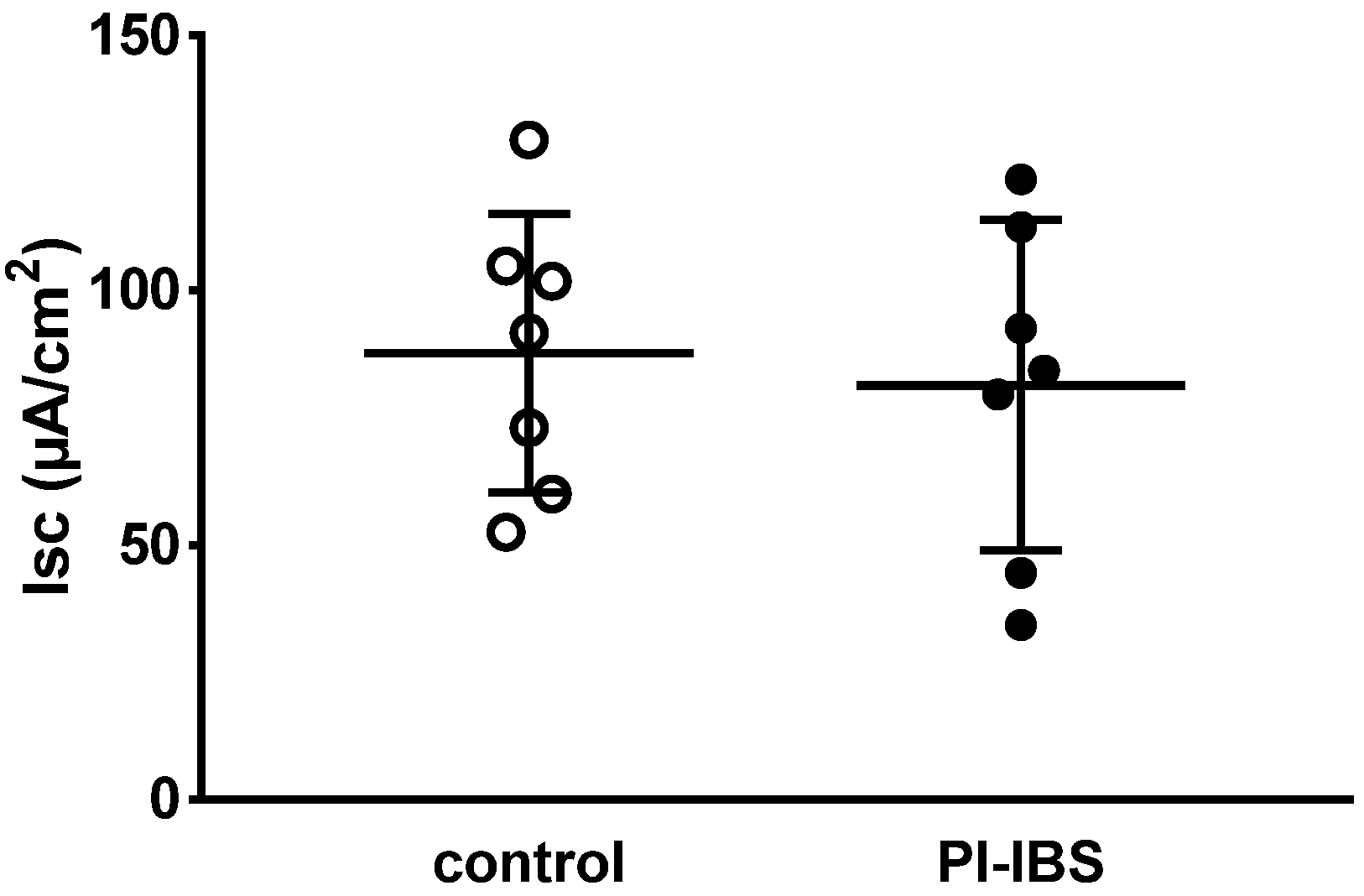
3.2. Active Sodium Absorption through the Epithelial Sodium Channel (ENaC) Was Not Affected
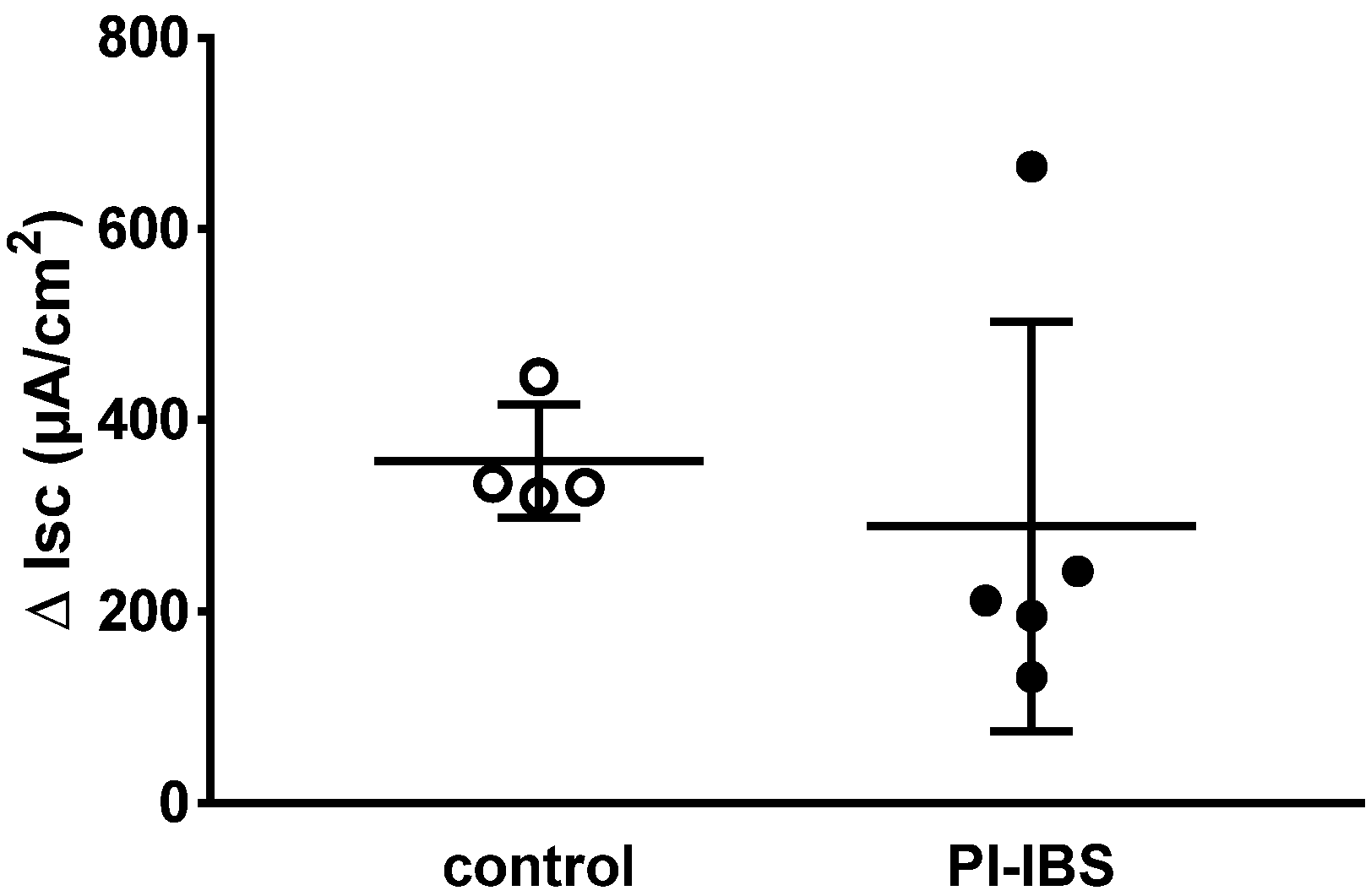
3.3. Electrogenic Anion Secretion
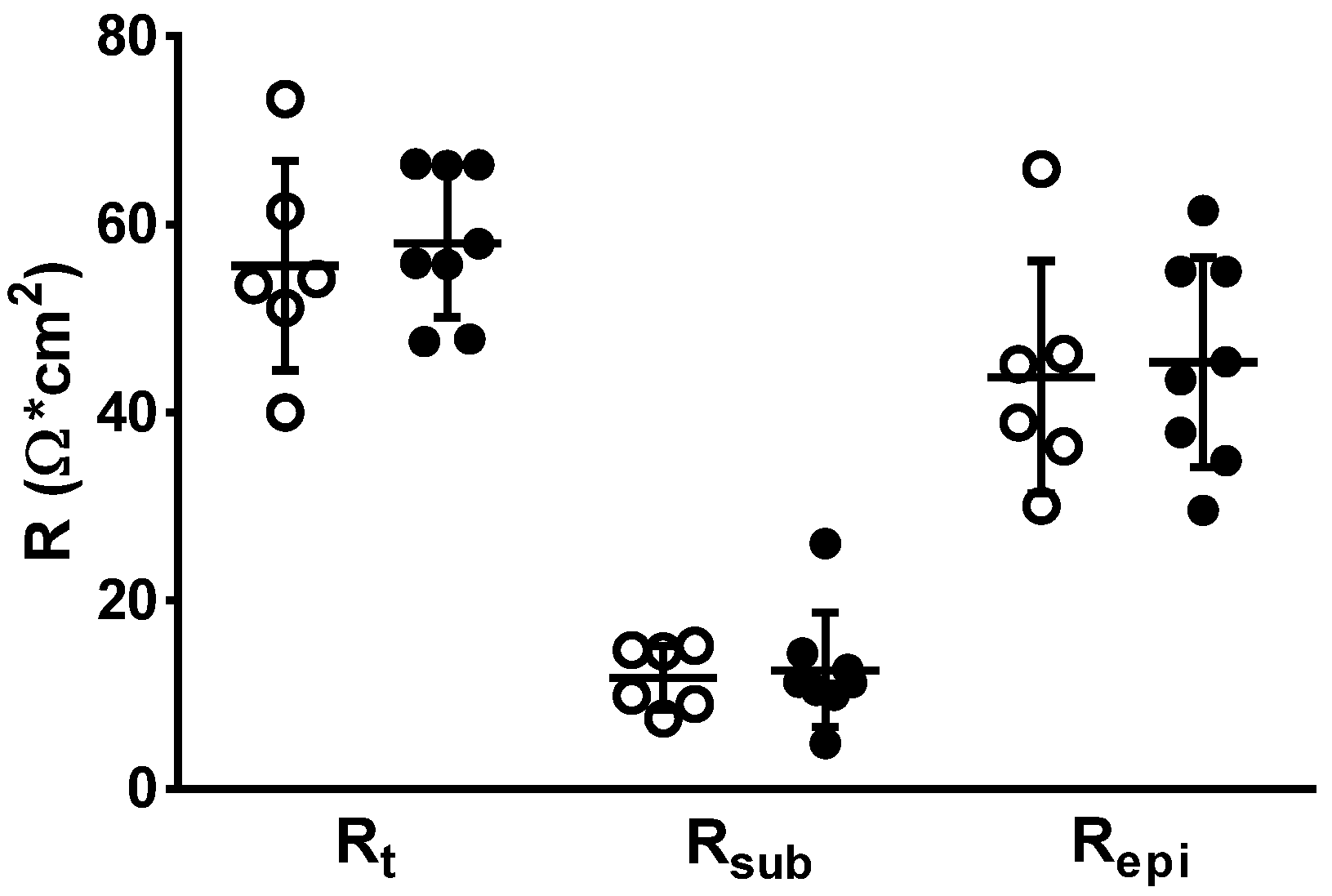
3.4. Impedance Spectroscopy for Determining Epithelial Resistance (Repi)
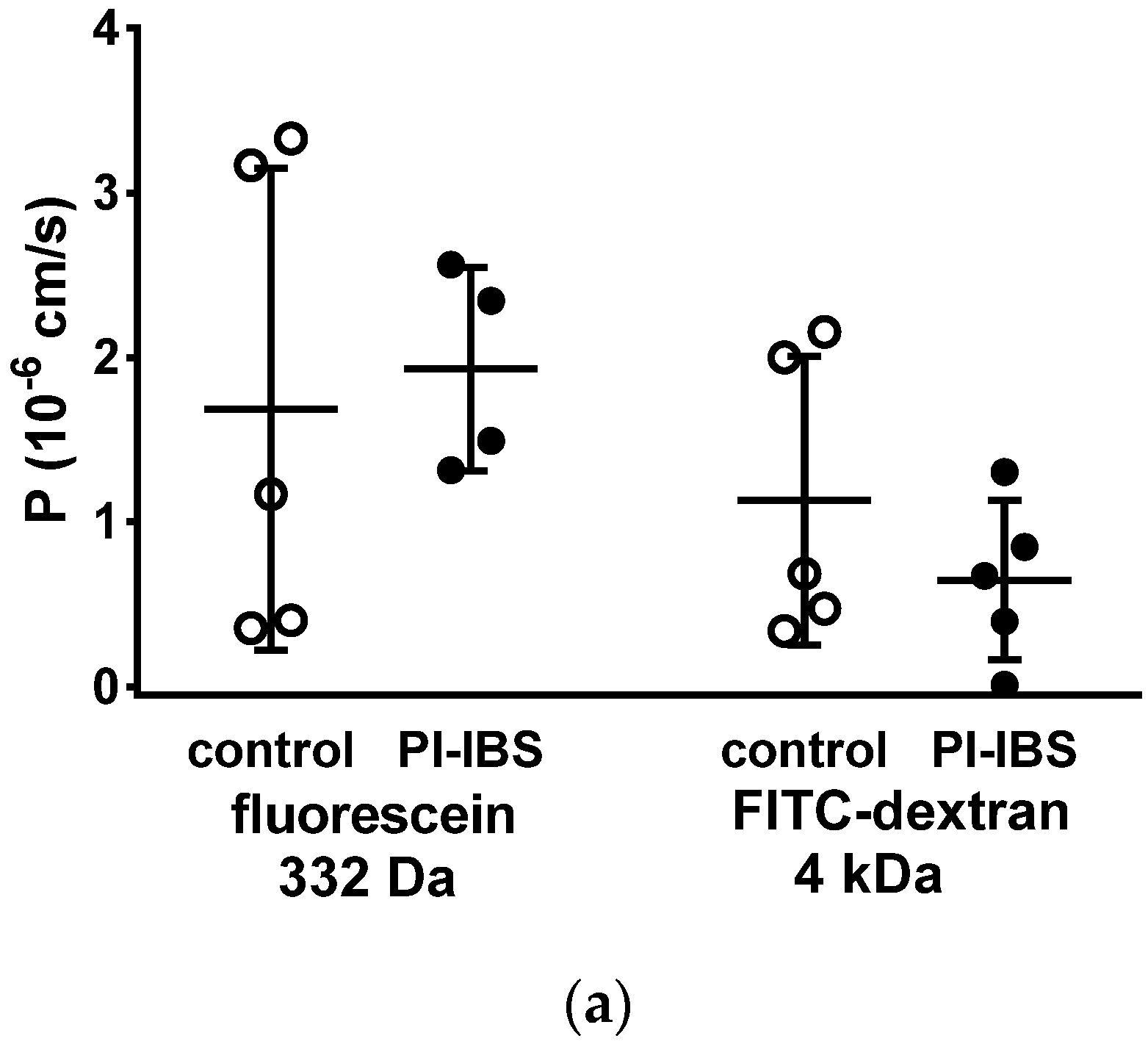
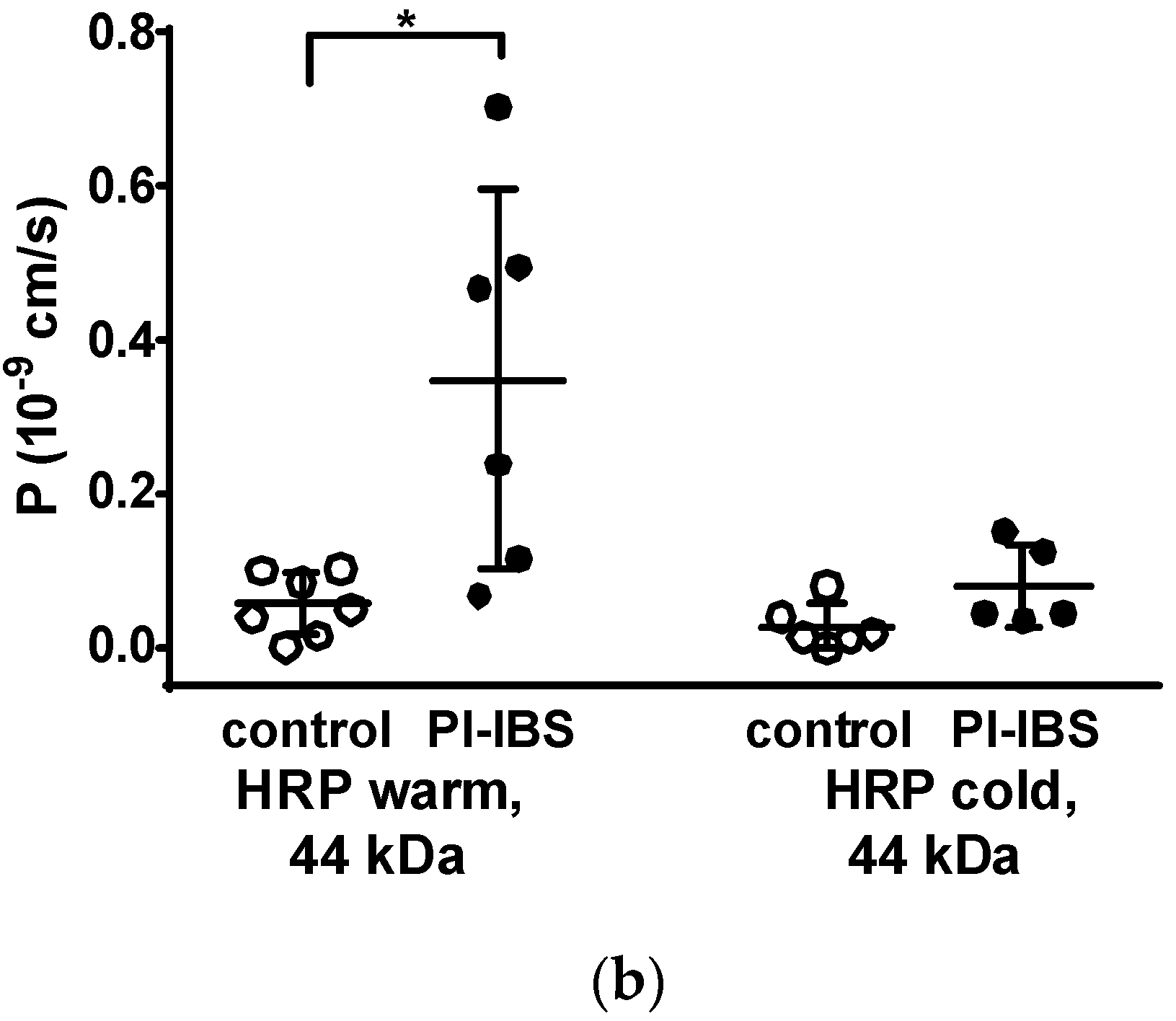
3.5. HRP Fluxes Indicate Macromolecule Permeability
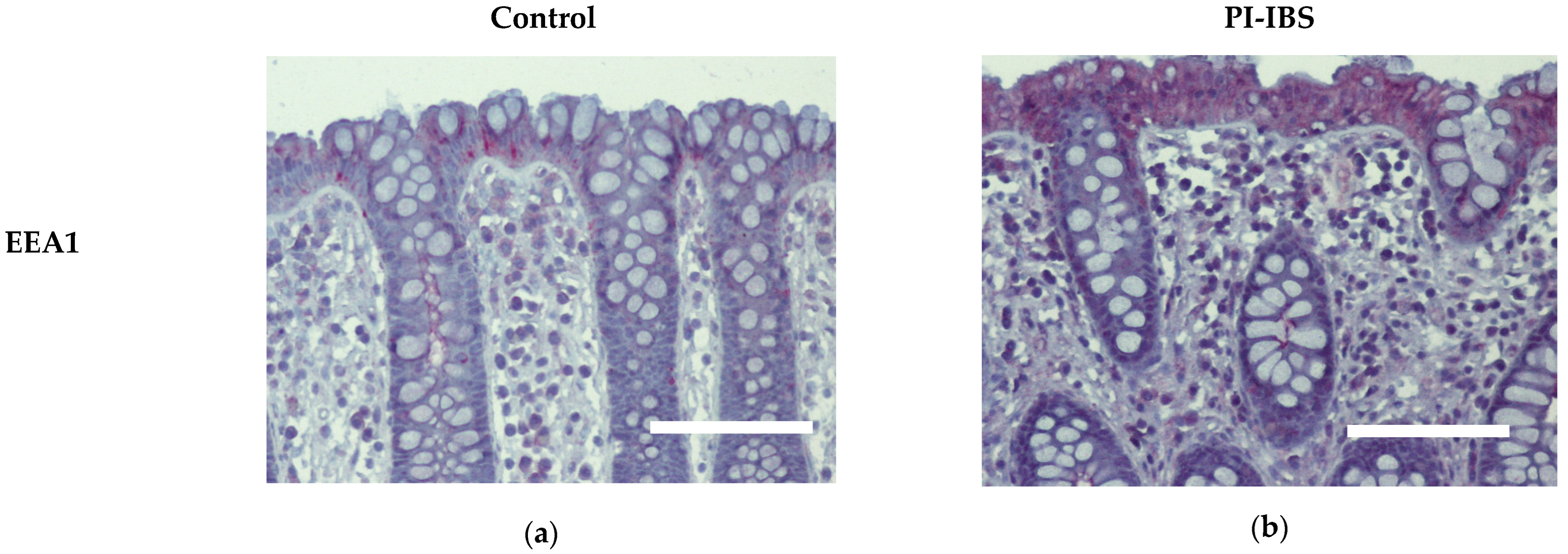
3.6. EEA1 as a Marker for Endocytosis
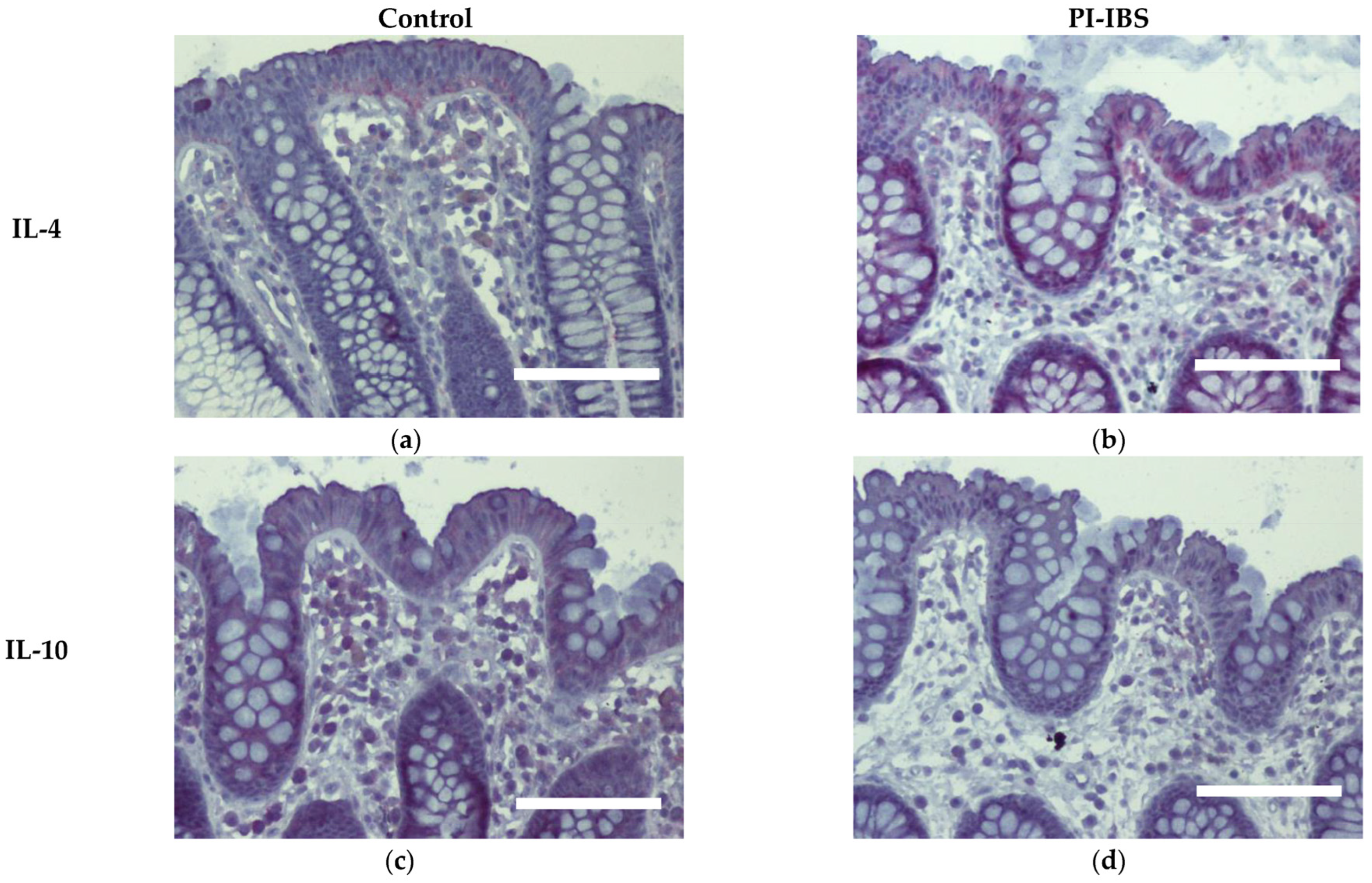
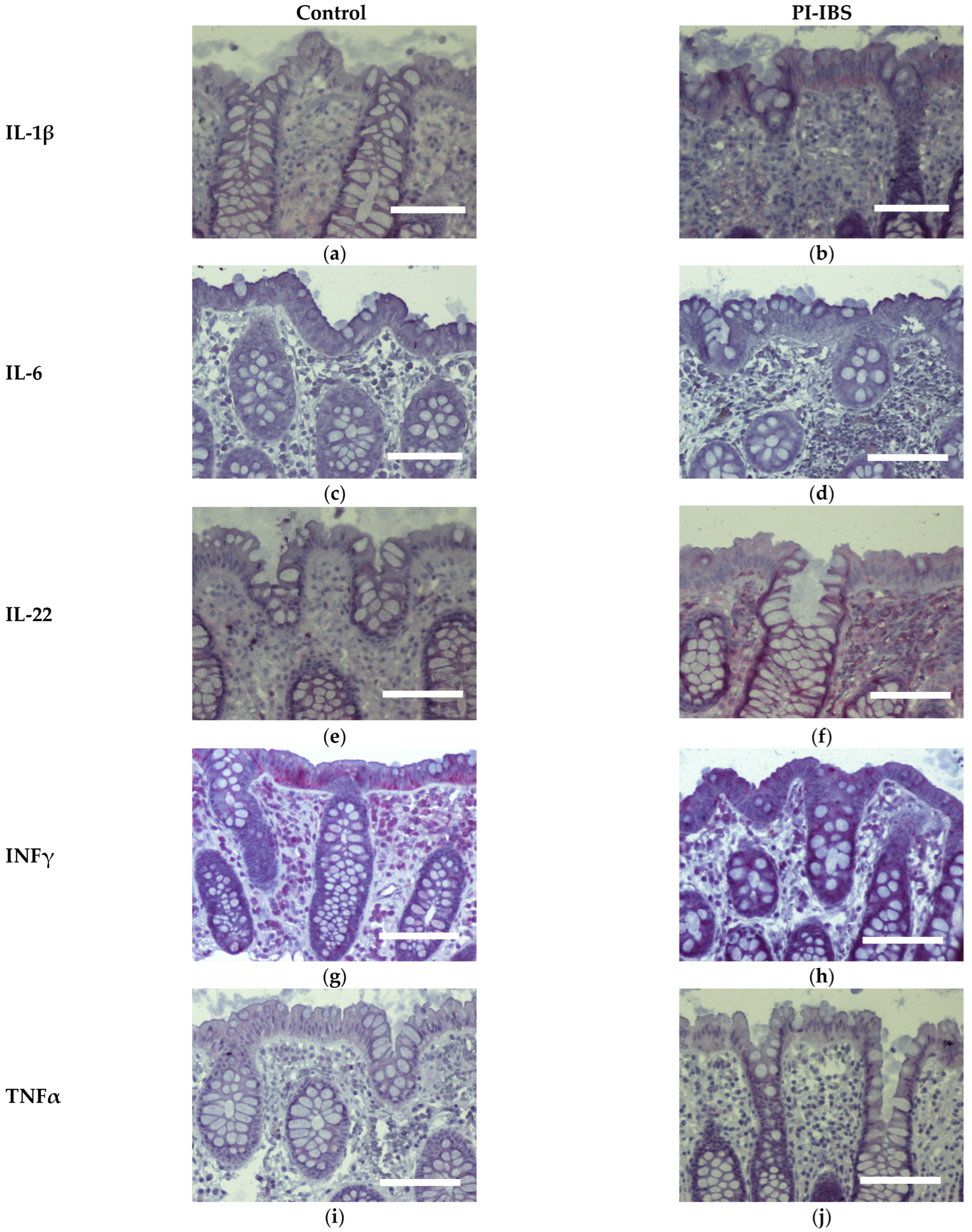
3.7. Cytokine Signals of IL-1β, IL-4 and IL-22 Are Intensified on Colon Epithelial Cells
4. Discussion
4.1. Characterization of Epithelial Barrier Function in the Colon
4.2. Diarrhea and/or Constipation in IBS
4.3. Leaky Gut Concept and Cytokine Induction of Barrier Disturbance
4.4. IBS, a Common Consequence from Campylobacteriosis with No Persistence of Campylobacter jejuni in the Microbiota of PI-IBS Patients
4.5. Therapeutic Approaches in PI-IBS
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spiller, R.C.; Jenkins, D.; Thornley, J.P.; Hebden, J.M.; Wright, T.; Skinner, M.; Neal, K.R. Increased rectal mucosal enteroendocrine cells, T lymphocytes, and increased gut permeability following acute Campylobacter enteritis and in post-dysenteric irritable bowel syndrome. Gut 2000, 47, 804–811. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Castaño-Rodríguez, N. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef]
- Lovell, R.M.; Ford, A.C. Global prevalence of and risk factors for irritable bowel syndrome: A meta-analysis. Clin. Gastroenterol. Hepatol. 2012, 10, 712–721. [Google Scholar]
- Scallan Walter, E.J.; Crim, S.M.; Bruce, B.B.; Griffin, P.M. Postinfectious Irritable Bowel Syndrome After Campylobacter Infection. Am. J. Gastroenterol. 2019, 114, 1649–1656. [Google Scholar] [CrossRef]
- Bücker, R.; Krug, S.M.; Moos, V.; Bojarski, C.; Schweiger, M.R.; Kerick, M.; Fromm, A.; Janßen, S.; Fromm, M.; Hering, N.A.; et al. Campylobacter jejuni impairs sodium transport and epithelial barrier function via cytokine release in human colon. Mucosal. Immunol. 2018, 11, 474–485. [Google Scholar] [CrossRef]
- de Sá, F.D.L.; Heimesaat, M.M.; Bereswill, S.; Nattramilarasu, P.K.; Schulzke, J.D.; Bücker, R. Resveratrol prevents Campylobacter jejuni-induced leaky gut by restoring occludin and claudin-5 in the paracellular leak pathway. Front. Pharmacol. 2021, 12, 640572. [Google Scholar] [CrossRef]
- Lobo de Sá, F.D.; Butkevych, E.; Nattramilarasu, P.K.; Fromm, A.; Mousavi, S.; Moos, V.; Golz, J.C.; Stingl, K.; Kittler, S.; Seinige, D.; et al. Curcumin Mitigates Immune-Induced Epithelial Barrier Dysfunction by Campylobacter jejuni. Int. J. Mol. Sci. 2019, 20, 4830. [Google Scholar] [CrossRef]
- Lobo de Sá, F.D.; Backert, S.; Nattramilarasu, P.K.; Mousavi, S.; Sandle, G.I.; Bereswill, S.; Heimesaat, M.M.; Schulzke, J.D.; Bücker, R. Vitamin D Reverses Disruption of Gut Epithelial Barrier Function Caused by Campylobacter jejuni. Int. J. Mol. Sci. 2021, 22, 8872. [Google Scholar] [CrossRef]
- Mayr, A.M.; Lick, S.; Bauer, J.; Thärigen, D.; Busch, U.; Huber, I. Rapid detection and differentiation of Campylobacter jejuni, Campylobacter coli and Campylobacter lari in food, using multiplex real-time PCR. J. Food Prot. 2010, 73, 241–250. [Google Scholar] [CrossRef]
- Bürgel, N.; Bojarski, C.; Mankertz, J.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Mechanisms of diarrhea in collagenous colitis. Gastroenterology 2002, 123, 433–443. [Google Scholar] [CrossRef]
- Barmeyer, C.; Erko, I.; Fromm, A.; Bojarski, C.; Loddenkemper, C.; Dames, P.; Kerick, M.; Siegmund, B.; Fromm, M.; Schweiger, M.R.; et al. ENaC dysregulation through activation of MEK1/2 contributes to impaired sodium absorption in lymphocytic colitis. Inflamm. Bowel Dis. 2016, 22, 539–547. [Google Scholar] [CrossRef]
- Bücker, R.; Schulz, E.; Günzel, D.; Bojarski, C.; Lee, I.F.; John, L.J.; Wiegand, S.; Janßen, T.; Wieler, L.H.; Dobrindt, U.; et al. α-Haemolysin of Escherichia coli in IBD: A potentiator of inflammatory activity in the colon. Gut 2014, 63, 1893–1901. [Google Scholar] [CrossRef]
- John, L. Mechanisms of Macromolecular Transcytosis during Crohn’s Disease: Investigations on the Cell Model T84 and Human Colon Biopsies. Ph.D. Dissertation, Freie Universität Berlin, Berlin, Germany, 2014. [Google Scholar]
- Berin, M.C.; Yang, P.C.; Ciok, L.; Waserman, S.; Perdue, M.H. Role for IL-4 in macromolecular transport across human intestinal epithelium. Am. J. Physiol. 1999, 276, C1046–C1052. [Google Scholar] [CrossRef]
- Cohen, E.; Ophir, I.; Henis, Y.I.; Bacher, A.; Ben Shaul, Y. Effect of temperature on the assembly of tight junctions and on the mobility of lipids in membranes of HT29 cells. J. Cell Sci. 1990, 97, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Punnonen, E.L.; Ryhänen, K.; Marjomäki, V.S. At reduced temperature, endocytic membrane traffic is blocked in multivesicular carrier endosomes in rat cardiac myocytes. Eur. J. Cell Biol. 1998, 75, 344–352. [Google Scholar] [CrossRef]
- Schinnerling, K.; Geelhaar-Karsch, A.; Allers, K.; Friebel, J.; Conrad, K.; Loddenkemper, C.; Kühl, A.; Erben, U.; Ignatius, R.; Moos, V.; et al. Role of dendritic cells in the pathogenesis of Whipple’s disease. Infect. Immun. 2015, 83, 482–491. [Google Scholar] [CrossRef]
- Buschmann, M.M.; Shen, L.; Rajapakse, H.; Raleigh, D.R.; Wang, Y.; Wang, Y.; Lingaraju, A.; Zha, J.; Abbott, E.; McAuley, E.M.; et al. Occludin OCEL-domain interactions are required for maintenance and regulation of the tight junction barrier to macromolecular flux. Mol. Biol. Cell. 2013, 24, 3056–3068. [Google Scholar] [CrossRef]
- Krug, S.M.; Amasheh, S.; Richter, J.F.; Milatz, S.; Günzel, D.; Westphal, J.K.; Huber, O.; Schulzke, J.D.; Fromm, M. Tricellulin forms a barrier to macromolecules in tricellular tight junctions without affecting ion permeability. Mol. Biol. Cell. 2009, 20, 3713–3724. [Google Scholar] [CrossRef]
- Bellono, N.W.; Bayrer, J.R.; Leitch, D.B.; Castro, J.; Zhang, C.; O’Donnell, T.A.; Brierley, S.M.; Ingraham, H.A.; Julius, D. Enterochromaffin Cells Are Gut Chemosensors that Couple to Sensory Neural Pathways. Cell 2017, 170, 185–198. [Google Scholar] [CrossRef]
- Ng, Q.X.; Soh, A.Y.S.; Loke, W.; Lim, D.Y.; Yeo, W.S. The role of inflammation in irritable bowel syndrome (IBS). J. Inflamm. Res. 2018, 11, 345–349. [Google Scholar] [CrossRef]
- Lobo de Sá, F.D.; Schulzke, J.D.; Bücker, R. Diarrheal Mechanisms and the Role of Intestinal Barrier Dysfunction in Campylobacter Infections. Curr. Top. Microbiol. Immunol. 2021, 431, 203–231. [Google Scholar] [PubMed]
- Miner-Williams, W.M.; Stevens, B.R.; Moughan, P.J. Are intact peptides absorbed from the healthy gut in the adult human? Nutr. Res. Rev. 2014, 27, 308–329. [Google Scholar] [CrossRef] [PubMed]
- Matysiak-Budnik, T.; Candalh, C.; Dugave, C.; Namane, A.; Cellier, C.; Cerf-Bensussan, N.; Heyman, M. Alterations of the intestinal transport and processing of gliadin peptides in celiac disease. Gastroenterology 2003, 125, 696–707. [Google Scholar] [CrossRef] [PubMed]
- Krug, S.M.; Bojarski, C.; Fromm, A.; Lee, I.M.; Dames, P.; Richter, J.F.; Turner, J.R.; Fromm, M.; Schulzke, J.D. Tricellulin is regulated via interleukin-13-receptor α2, affects macromolecule uptake, and is decreased in ulcerative colitis. Mucosal Immunol. 2018, 11, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Gwee, K.A.; Collins, S.M.; Read, N.W.; Rajnakova, A.; Deng, Y.; Graham, J.C.; McKendrick, M.W.; Moochhala, S.M. Increased rectal mucosal expression of interleukin 1beta in recently acquired post-infectious irritable bowel syndrome. Gut 2003, 52, 523–526. [Google Scholar] [CrossRef]
- Macsharry, J.; O’Mahony, L.; Fanning, A.; Bairead, E.; Sherlock, G.; Tiesman, J.; Fulmer, A.; Kiely, B.; Dinan, T.G.; Shanahan, F.; et al. Mucosal cytokine imbalance in irritable bowel syndrome. Scand. J. Gastroenterol. 2008, 43, 1467–1476. [Google Scholar] [CrossRef]
- Awad, K. Epitheliale Barrierefunktion des Kolons Beim DIARRHOE-Dominanten und Beim Mischtyp-Reizdarmsyndrom. Ph.D. Dissertation, Charité—Universitätsmedizin Berlin, Berlin, Germany, 2022. [Google Scholar]
- Brooks, P.T.; Brakel, K.A.; Bell, J.A.; Bejcek, C.E.; Gilpin, T.; Brudvig, J.M.; Mansfield, L.S. Transplanted human fecal microbiota enhanced Guillain Barré syndrome autoantibody responses after Campylobacter jejuni infection in C57BL/6 mice. Microbiome 2017, 5, 92. [Google Scholar] [CrossRef]
- Parthasarathy, G.; Mansfield, L.S. Recombinant interleukin-4 enhances Campylobacter jejuni invasion of intestinal pig epithelial cells (IPEC-1). Microb. Pathog. 2009, 47, 38–46. [Google Scholar] [CrossRef]
- Edwards, L.A.; Nistala, K.; Mills, D.C.; Stephenson, H.N.; Zilbauer, M.; Wren, B.W.; Dorrell, N.; Lindley, K.J.; Wedderburn, L.R.; Bajaj-Elliott, M. Delineation of the innate and adaptive T-cell immune outcome in the human host in response to Campylobacter jejuni infection. PLoS ONE 2010, 5, e15398. [Google Scholar] [CrossRef]
- Brand, S.; Beigel, F.; Olszak, T.; Zitzmann, K.; Eichhorst, S.T.; Otte, J.M.; Diepolder, H.; Marquardt, A.; Jagla, W.; Popp, A.; et al. IL-22 is increased in active Crohn’s disease and promotes proinflammatory gene expression and intestinal epithelial cell migration. Am. J. Physiol. Gastrointest Liver Physiol. 2006, 290, G827–G838. [Google Scholar] [CrossRef]
- Yuki, N.; Kuwabara, S. Axonal Guillain-Barré syndrome: Carbohydrate mimicry and pathophysiology. J. Peripher. Nerv. Syst. 2007, 12, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Morales, W.; Triantafyllou, K.; Parodi, G.; Weitsman, S.; Park, S.C.; Rezaie, A.; Pichetshote, N.; Lin, E.; Pimentel, M. Immunization with cytolethal distending toxin B produces autoantibodies to vinculin and small bowel bacterial changes in a rat model of postinfectious irritable bowel syndrome. Neurogastroenterol. Motil. 2020, 32, e13875. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, M.; Morales, W.; Pokkunuri, V.; Brikos, C.; Kim, S.M.; Kim, S.E.; Triantafyllou, K.; Weitsman, S.; Marsh, Z.; Marsh, E.; et al. Autoimmunity Links Vinculin to the Pathophysiology of Chronic Functional Bowel Changes Following Campylobacter jejuni Infection in a Rat Model. Dig Dis. Sci. 2015, 60, 1195–1205. [Google Scholar] [CrossRef] [PubMed]
- Barbara, G.; Wang, B.; Stanghellini, V.; de Giorgio, R.; Cremon, C.; Di Nardo, G.; Trevisani, M.; Campi, B.; Geppetti, P.; Tonini, M.; et al. Mast cell-dependent excitation of visceral-nociceptive sensory neurons in irritable bowel syndrome. Gastroenterology 2007, 132, 26–37. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Control (Mean ± SD) | PI-IBS (Mean ± SD) | p Value |
|---|---|---|---|
| EEA1 | 1.87 ± 0.20 | 3.57 ± 0.27 | 0.0005 *** |
| IL-1β | 1.57 ± 0.24 | 2.90 ± 0.40 | 0.0168 * |
| IL-4 | 2.90 ± 0.29 | 3.83 ± 0.13 | 0.0152 * |
| IL-6 | 1.10 ± 0.07 | 1.10 ± 0.04 | 0.9999 ns |
| IL-10 | 2.43 ± 0.36 | 1.97 ± 0.47 | 0.4444 ns |
| IL-22 | 1.63 ± 0.26 | 2.80 ± 0.33 | 0.0190 * |
| INFγ | 3.03 ± 0.32 | 2.93 ± 0.31 | 0.8265 ns |
| TNFα | 1.43 ± 0.16 | 1.33 ± 0.11 | 0.6171 ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omarova, S.; Awad, K.; Moos, V.; Püning, C.; Gölz, G.; Schulzke, J.-D.; Bücker, R. Intestinal Barrier in Post-Campylobacter jejuni Irritable Bowel Syndrome. Biomolecules 2023, 13, 449. https://doi.org/10.3390/biom13030449
Omarova S, Awad K, Moos V, Püning C, Gölz G, Schulzke J-D, Bücker R. Intestinal Barrier in Post-Campylobacter jejuni Irritable Bowel Syndrome. Biomolecules. 2023; 13(3):449. https://doi.org/10.3390/biom13030449
Chicago/Turabian StyleOmarova, Sholpan, Karem Awad, Verena Moos, Christoph Püning, Greta Gölz, Jörg-Dieter Schulzke, and Roland Bücker. 2023. "Intestinal Barrier in Post-Campylobacter jejuni Irritable Bowel Syndrome" Biomolecules 13, no. 3: 449. https://doi.org/10.3390/biom13030449
APA StyleOmarova, S., Awad, K., Moos, V., Püning, C., Gölz, G., Schulzke, J.-D., & Bücker, R. (2023). Intestinal Barrier in Post-Campylobacter jejuni Irritable Bowel Syndrome. Biomolecules, 13(3), 449. https://doi.org/10.3390/biom13030449