
Research Progress in Calcitonin Gene-Related Peptide and Bone Repair
Abstract
:1. Overview of CGRP
1.1. Structure of CGRP
1.2. Receptors of CGRP
1.3. CGRP Synthesis, Storage, and Release
1.4. Physiology and Pathophysiology of CGRP
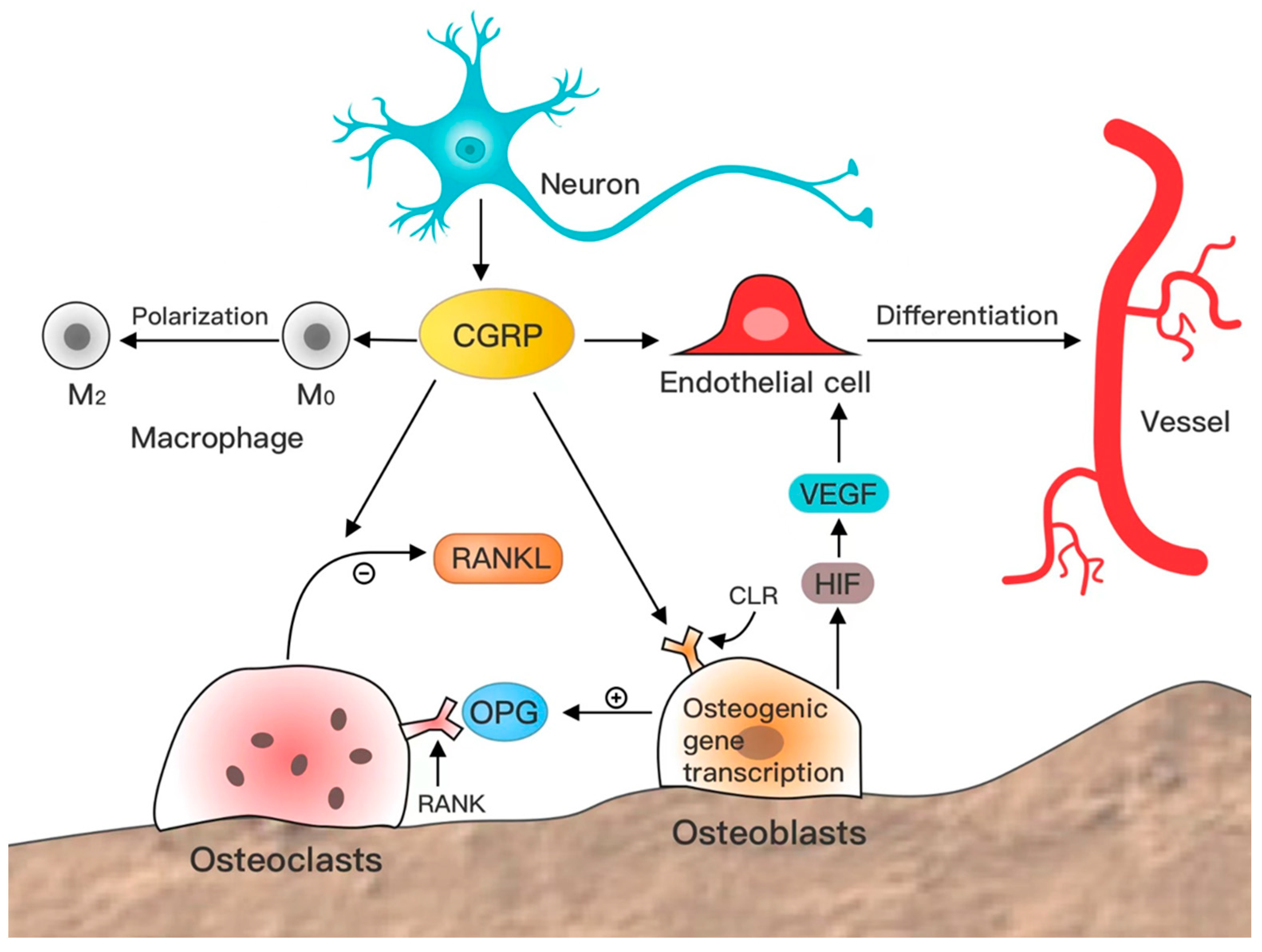
2. Effects of CGRP on Bone Repair
2.1. Distribution of CGRP and Receptors in Bone Tissue
2.2. CGRP Promotes Osteogenesis
2.3. CGRP Inhibits Bone Resorption
2.4. CGRP-Induced Angiogenesis
2.5. CGRP Regulates the Immune Microenvironment
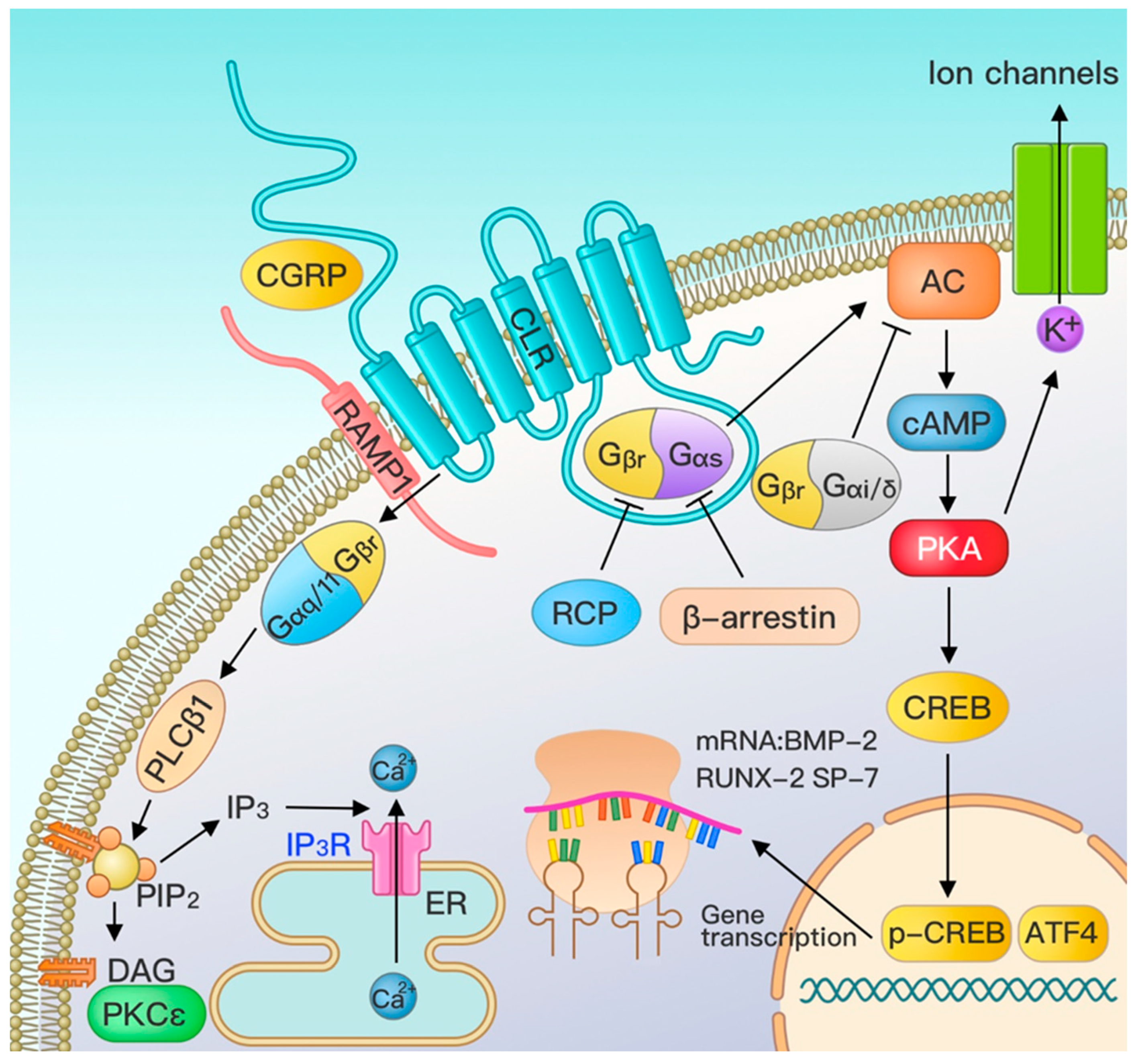
3. Effect of CGRP on Osteogenic Effect Pathways
3.1. Major Pathways of CGRP—G Protein-Coupled Receptor Pathway
3.2. Other Signal Pathway Crosstalk
4. CGRP in Orthopedic Treatment
4.1. Prospects for the Application of CGRP as a Drug
4.2. Biomaterials Can Promote Bone Repair by Modulating CGRP
4.3. Modulation of CGRP for Bone Repair by Electrical Stimulation
5. Other Insights on CGRP and Bone Repair Studies
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Russell, F.A.; King, R.; Smillie, S.J.; Kodji, X.; Brain, S.D. Calcitonin gene-related peptide: Physiology and pathophysiology. Physiol. Rev. 2014, 94, 1099–1142. [Google Scholar] [CrossRef]
- Edvinsson, L. The Trigeminovascular Pathway: Role of CGRP and CGRP Receptors in Migraine. Headache 2017, 57, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Mulderry, P.K.; Ghatei, M.A.; Spokes, R.A.; Jones, P.M.; Pierson, A.M.; Hamid, Q.A.; Kanse, S.; Amara, S.G.; Burrin, J.M.; Legon, S.; et al. Differential expression of alpha-CGRP and beta-CGRP by primary sensory neurons and enteric autonomic neurons of the rat. Neuroscience 1988, 25, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Conner, A.C.; Hay, D.L.; Howitt, S.G.; Kilk, K.; Langel, U.; Wheatley, M.; Smith, D.M.; Poyner, D.R. Interaction of calcitonin-gene-related peptide with its receptors. Biochem. Soc. Trans. 2002, 30, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Watkins, H.A.; Rathbone, D.L.; Barwell, J.; Hay, D.L.; Poyner, D.R. Structure-activity relationships for α-calcitonin gene-related peptide. Br. J. Pharmacol. 2013, 170, 1308–1322. [Google Scholar] [CrossRef]
- Chang, C.P.; Pearse, R.V., 2nd; O’Connell, S.; Rosenfeld, M.G. Identification of a seven transmembrane helix receptor for corticotropin-releasing factor and sauvagine in mammalian brain. Neuron 1993, 11, 1187–1195. [Google Scholar] [CrossRef]
- Flühmann, B.; Muff, R.; Hunziker, W.; Fischer, J.A.; Born, W. A human orphan calcitonin receptor-like structure. Biochem. Biophys. Res. Commun. 1995, 206, 341–347. [Google Scholar] [CrossRef]
- Aiyar, N.; Rand, K.; Elshourbagy, N.A.; Zeng, Z.; Adamou, J.E.; Bergsma, D.J.; Li, Y. A cDNA encoding the calcitonin gene-related peptide type 1 receptor. J. Biol. Chem. 1996, 271, 11325–11329. [Google Scholar] [CrossRef] [PubMed]
- McLatchie, L.M.; Fraser, N.J.; Main, M.J.; Wise, A.; Brown, J.; Thompson, N.; Solari, R.; Lee, M.G.; Foord, S.M. RAMPs regulate the transport and ligand specificity of the calcitonin-receptor-like receptor. Nature 1998, 393, 333–339. [Google Scholar] [CrossRef]
- Luebke, A.E.; Dahl, G.P.; Roos, B.A.; Dickerson, I.M. Identification of a protein that confers calcitonin gene-related peptide responsiveness to oocytes by using a cystic fibrosis transmembrane conductance regulator assay. Proc. Natl. Acad. Sci. USA 1996, 93, 3455–3460. [Google Scholar] [CrossRef]
- Egea, S.C.; Dickerson, I.M. Direct interactions between calcitonin-like receptor (CLR) and CGRP-receptor component protein (RCP) regulate CGRP receptor signaling. Endocrinology 2012, 153, 1850–1860. [Google Scholar] [CrossRef] [PubMed]
- Evans, B.N.; Rosenblatt, M.I.; Mnayer, L.O.; Oliver, K.R.; Dickerson, I.M. CGRP-RCP, a novel protein required for signal transduction at calcitonin gene-related peptide and adrenomedullin receptors. J. Biol. Chem. 2000, 275, 31438–31443. [Google Scholar] [CrossRef]
- Choi, J.E.; Di Nardo, A. Skin neurogenic inflammation. Semin. Immunopathol. 2018, 40, 249–259. [Google Scholar] [CrossRef]
- Arkless, K.; Argunhan, F.; Brain, S.D. CGRP Discovery and Timeline. Handb. Exp. Pharmacol. 2019, 255, 1–12. [Google Scholar] [PubMed]
- Mi, J.; Xu, J.K.; Yao, Z.; Yao, H.; Li, Y.; He, X.; Dai, B.Y.; Zou, L.; Tong, W.X.; Zhang, X.T.; et al. Implantable Electrical Stimulation at Dorsal Root Ganglions Accelerates Osteoporotic Fracture Healing via Calcitonin Gene-Related Peptide. Adv. Sci. (Weinh) 2022, 9, e2103005. [Google Scholar] [CrossRef]
- Messlinger, K. The big CGRP flood-sources, sinks and signalling sites in the trigeminovascular system. J. Headache Pain 2018, 19, 22. [Google Scholar] [CrossRef]
- McCulloch, J.; Uddman, R.; Kingman, T.A.; Edvinsson, L. Calcitonin gene-related peptide: Functional role in cerebrovascular regulation. Proc. Natl. Acad. Sci. USA 1986, 83, 5731–5735. [Google Scholar] [CrossRef]
- Ursell, P.C.; Ren, C.L.; Albala, A.; Danilo, P., Jr. Nonadrenergic noncholinergic innervation. Anatomic distribution of calcitonin gene-related peptide-immunoreactive tissue in the dog heart. Circ. Res. 1991, 68, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Amrutkar, D.V.; Mataji, A.; Salmasi, H.; Hay-Schmidt, A.; Sheykhzade, M.; Messlinger, K.; Olesen, J.; Jansen-Olesen, I. Evidence for CGRP re-uptake in rat dura mater encephali. Br. J. Pharmacol. 2010, 161, 1885–1898. [Google Scholar] [CrossRef]
- Portaluppi, F.; Trasforini, G.; Margutti, A.; Vergnani, L.; Ambrosio, M.R.; Rossi, R.; Bagni, B.; Pansini, R.; degli Uberti, E.C. Circadian rhythm of calcitonin gene-related peptide in uncomplicated essential hypertension. J. Hypertens. 1992, 10, 1227–1234. [Google Scholar] [CrossRef]
- Wang, F.; Deng, Y.; Wang, J.; Yu, L.; Ding, F.; Lian, W.; Liu, Q.; Lin, X. The PLGA nanoparticles for sustainable release of CGRP to ameliorate the inflammatory and vascular disorders in the lung of CGRP-deficient rats. Drug Deliv. 2021, 28, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Sohn, I.; Sheykhzade, M.; Edvinsson, L.; Sams, A. The effects of CGRP in vascular tissue—Classical vasodilation, shadowed effects and systemic dilemmas. Eur. J. Pharmacol. 2020, 881, 173205. [Google Scholar] [CrossRef] [PubMed]
- Smillie, S.J.; King, R.; Kodji, X.; Outzen, E.; Pozsgai, G.; Fernandes, E.; Marshall, N.; de Winter, P.; Heads, R.J.; Dessapt-Baradez, C.; et al. An ongoing role of α-calcitonin gene-related peptide as part of a protective network against hypertension, vascular hypertrophy, and oxidative stress. Hypertension 2014, 63, 1056–1062. [Google Scholar] [CrossRef]
- Petersen, K.A.; Birk, S.; Lassen, L.H.; Kruuse, C.; Jonassen, O.; Lesko, L.; Olesen, J. The CGRP-antagonist, BIBN4096BS does not affect cerebral or systemic haemodynamics in healthy volunteers. Cephalalgia 2005, 25, 139–147. [Google Scholar] [CrossRef]
- Ichikawa, H.; Matsuo, S.; Wakisaka, S.; Akai, M. Fine structure of calcitonin gene-related peptide-immunoreactive nerve fibres in the rat temporomandibular joint. Arch. Oral Biol. 1990, 35, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Neychev, D.; Sbirkova, T.; Ivanovska, M.; Raycheva, R.; Murdjeva, M.; Atanasov, D. Correlation between CGRP Levels and the Neuropathic and Inflammatory Component of Postoperative Pain. Folia Med. 2020, 62, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Schou, W.S.; Ashina, S.; Amin, F.M.; Goadsby, P.J.; Ashina, M. Calcitonin gene-related peptide and pain: A systematic review. J. Headache Pain 2017, 18, 34. [Google Scholar] [CrossRef] [PubMed]
- Nakamura-Craig, M.; Gill, B.K. Effect of neurokinin A, substance P and calcitonin gene related peptide in peripheral hyperalgesia in the rat paw. Neurosci. Lett. 1991, 124, 49–51. [Google Scholar] [CrossRef]
- Mogil, J.S.; Miermeister, F.; Seifert, F.; Strasburg, K.; Zimmermann, K.; Reinold, H.; Austin, J.S.; Bernardini, N.; Chesler, E.J.; Hofmann, H.A.; et al. Variable sensitivity to noxious heat is mediated by differential expression of the CGRP gene. Proc. Natl. Acad. Sci. USA 2005, 102, 12938–12943. [Google Scholar] [CrossRef]
- Iyengar, S.; Johnson, K.W.; Ossipov, M.H.; Aurora, S.K. CGRP and the Trigeminal System in Migraine. Headache 2019, 59, 659–681. [Google Scholar] [CrossRef]
- Russo, A.F. Calcitonin gene-related peptide (CGRP): A new target for migraine. Annu. Rev. Pharmacol. Toxicol. 2015, 55, 533–552. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.F.; Hay, D.L. CGRP physiology, pharmacology, and therapeutic targets: Migraine and beyond. Physiol. Rev. 2023, 103, 1565–1644. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, L.; Haanes, K.A.; Warfvinge, K.; Krause, D.N. CGRP as the target of new migraine therapies—successful translation from bench to clinic. Nat Rev Neurol 2018, 14, 338–350. [Google Scholar] [CrossRef]
- Sánchez-Robles, E.M.; Girón, R.; Paniagua, N.; Rodríguez-Rivera, C.; Pascual, D.; Goicoechea, C. Monoclonal Antibodies for Chronic Pain Treatment: Present and Future. Int. J. Mol. Sci. 2021, 22, 10325. [Google Scholar] [CrossRef]
- Doods, H.; Hallermayer, G.; Wu, D.; Entzeroth, M.; Rudolf, K.; Engel, W.; Eberlein, W. Pharmacological profile of BIBN4096BS, the first selective small molecule CGRP antagonist. Br. J. Pharmacol. 2000, 129, 420–423. [Google Scholar] [CrossRef]
- Paone, D.V.; Shaw, A.W.; Nguyen, D.N.; Burgey, C.S.; Deng, J.Z.; Kane, S.A.; Koblan, K.S.; Salvatore, C.A.; Mosser, S.D.; Johnston, V.K.; et al. Potent, orally bioavailable calcitonin gene-related peptide receptor antagonists for the treatment of migraine: Discovery of N-[(3R,6S)-6-(2,3-difluorophenyl)-2-oxo-1- (2,2,2-trifluoroethyl)azepan-3-yl]-4- (2-oxo-2,3-dihydro-1H-imidazo[4,5-b]pyridin- 1-yl)piperidine-1-carboxamide (MK-0974). J. Med. Chem. 2007, 50, 5564–5567. [Google Scholar]
- Hewitt, D.J.; Aurora, S.K.; Dodick, D.W.; Goadsby, P.J.; Ge, Y.J.; Bachman, R.; Taraborelli, D.; Fan, X.; Assaid, C.; Lines, C.; et al. Randomized controlled trial of the CGRP receptor antagonist MK-3207 in the acute treatment of migraine. Cephalalgia 2011, 31, 712–722. [Google Scholar] [CrossRef]
- Croop, R.; Madonia, J.; Stock, D.A.; Thiry, A.; Forshaw, M.; Murphy, A.; Coric, V.; Lipton, R.B. Zavegepant nasal spray for the acute treatment of migraine: A Phase 2/3 double-blind, randomized, placebo-controlled, dose-ranging trial. Headache 2022, 62, 1153–1163. [Google Scholar] [CrossRef]
- Diener, H.C.; Barbanti, P.; Dahlöf, C.; Reuter, U.; Habeck, J.; Podhorna, J. BI 44370 TA, an oral CGRP antagonist for the treatment of acute migraine attacks: Results from a phase II study. Cephalalgia 2011, 31, 573–584. [Google Scholar] [CrossRef]
- Markham, A. Erenumab: First Global Approval. Drugs 2018, 78, 1157–1161. [Google Scholar] [CrossRef]
- Scott, L.J. Rimegepant: First Approval. Drugs 2020, 80, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.J. Ubrogepant: First Approval. Drugs 2020, 80, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Deeks, E.D. Atogepant: First Approval. Drugs 2022, 82, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Lamb, Y.N. Galcanezumab: First Global Approval. Drugs 2018, 78, 1769–1775. [Google Scholar] [CrossRef] [PubMed]
- Hoy, S.M. Fremanezumab: First Global Approval. Drugs 2018, 78, 1829–1834. [Google Scholar] [CrossRef]
- Dhillon, S. Eptinezumab: First Approval. Drugs 2020, 80, 733–739. [Google Scholar] [CrossRef]
- Smillie, S.J.; Brain, S.D. Calcitonin gene-related peptide (CGRP) and its role in hypertension. Neuropeptides 2011, 45, 93–104. [Google Scholar] [CrossRef]
- Sun, L.; Wang, G.; He, M.; Mei, Z.; Zhang, F.; Liu, P. Effect and mechanism of the CACNA2D1-CGRP pathway in osteoarthritis-induced ongoing pain. Biomed. Pharmacother. 2020, 129, 110374. [Google Scholar] [CrossRef]
- Bjurholm, A.; Kreicbergs, A.; Brodin, E.; Schultzberg, M. Substance P- and CGRP-immunoreactive nerves in bone. Peptides 1988, 9, 165–171. [Google Scholar] [CrossRef]
- Haegerstrand, A.; Dalsgaard, C.J.; Jonzon, B.; Larsson, O.; Nilsson, J. Calcitonin gene-related peptide stimulates proliferation of human endothelial cells. Proc. Natl. Acad. Sci. USA 1990, 87, 3299–3303. [Google Scholar] [CrossRef]
- Hosoi, J.; Murphy, G.F.; Egan, C.L.; Lerner, E.A.; Grabbe, S.; Asahina, A.; Granstein, R.D. Regulation of Langerhans cell function by nerves containing calcitonin gene-related peptide. Nature 1993, 363, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Naot, D.; Cornish, J. The role of peptides and receptors of the calcitonin family in the regulation of bone metabolism. Bone 2008, 43, 813–818. [Google Scholar] [CrossRef] [PubMed]
- Chrissobolis, S.; Zhang, Z.; Kinzenbaw, D.A.; Lynch, C.M.; Russo, A.F.; Faraci, F.M. Receptor activity-modifying protein-1 augments cerebrovascular responses to calcitonin gene-related peptide and inhibits angiotensin II-induced vascular dysfunction. Stroke 2010, 41, 2329–2334. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Qu, J.; Zhu, H.; Wang, J.; He, H.; Xie, X.; Wu, R.; Lu, Q. Corrigendum: CGRP Regulates the Age-Related Switch Between Osteoblast and Adipocyte Differentiation. Front Cell Dev. Biol. 2021, 9, 715740. [Google Scholar] [CrossRef]
- Pajarinen, J.; Lin, T.; Gibon, E.; Kohno, Y.; Maruyama, M.; Nathan, K.; Lu, L.; Yao, Z.; Goodman, S.B. Mesenchymal stem cell-macrophage crosstalk and bone healing. Biomaterials 2019, 196, 80–89. [Google Scholar] [CrossRef]
- Marrella, A.; Lee, T.Y.; Lee, D.H.; Karuthedom, S.; Syla, D.; Chawla, A.; Khademhosseini, A.; Jang, H.L. Engineering vascularized and innervated bone biomaterials for improved skeletal tissue regeneration. Mater. Today 2018, 21, 362–376. [Google Scholar] [CrossRef]
- Brazill, J.M.; Beeve, A.T.; Craft, C.S.; Ivanusic, J.J.; Scheller, E.L. Nerves in Bone: Evolving Concepts in Pain and Anabolism. J. Bone Miner. Res. 2019, 34, 1393–1406. [Google Scholar] [CrossRef]
- Ballica, R.; Valentijn, K.; Khachatryan, A.; Guerder, S.; Kapadia, S.; Gundberg, C.; Gilligan, J.; Flavell, R.A.; Vignery, A. Targeted expression of calcitonin gene-related peptide to osteoblasts increases bone density in mice. J. Bone Miner. Res. 1999, 14, 1067–1074. [Google Scholar] [CrossRef]
- Schinke, T.; Liese, S.; Priemel, M.; Haberland, M.; Schilling, A.F.; Catala-Lehnen, P.; Blicharski, D.; Rueger, J.M.; Gagel, R.F.; Emeson, R.B.; et al. Decreased bone formation and osteopenia in mice lacking alpha-calcitonin gene-related peptide. J. Bone Miner. Res. 2004, 19, 2049–2056. [Google Scholar] [CrossRef]
- Hukkanen, M.; Konttinen, Y.T.; Rees, R.G.; Gibson, S.J.; Santavirta, S.; Polak, J.M. Innervation of bone from healthy and arthritic rats by substance P and calcitonin gene related peptide containing sensory fibers. J. Rheumatol. 1992, 19, 1252–1259. [Google Scholar]
- Hu, B.; Lv, X.; Wei, L.; Wang, Y.; Zheng, G.; Yang, C.; Zang, F.; Wang, J.; Li, J.; Wu, X.; et al. Sensory Nerve Maintains Intervertebral Disc Extracellular Matrix Homeostasis Via CGRP/CHSY1 Axis. Adv. Sci. (Weinh) 2022, 9, e2202620. [Google Scholar] [CrossRef]
- Vignery, A.; McCarthy, T.L. The neuropeptide calcitonin gene-related peptide stimulates insulin-like growth factor I production by primary fetal rat osteoblasts. Bone 1996, 18, 331–335. [Google Scholar] [CrossRef]
- Liang, W.; Zhuo, X.; Tang, Z.; Wei, X.; Li, B. Calcitonin gene-related peptide stimulates proliferation and osteogenic differentiation of osteoporotic rat-derived bone mesenchymal stem cells. Mol. Cell. Biochem. 2015, 402, 101–110. [Google Scholar] [CrossRef]
- He, H.; Chai, J.; Zhang, S.; Ding, L.; Yan, P.; Du, W.; Yang, Z. CGRP may regulate bone metabolism through stimulating osteoblast differentiation and inhibiting osteoclast formation. Mol. Med. Rep. 2016, 13, 3977–3984. [Google Scholar] [CrossRef]
- Elefteriou, F.; Ahn, J.D.; Takeda, S.; Starbuck, M.; Yang, X.; Liu, X.; Kondo, H.; Richards, W.G.; Bannon, T.W.; Noda, M.; et al. Leptin regulation of bone resorption by the sympathetic nervous system and CART. Nature 2005, 434, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Functions of Osteocalcin in Bone, Pancreas, Testis, and Muscle. Int. J. Mol. Sci. 2020, 21, 7513. [Google Scholar] [CrossRef]
- Wang, Y.; Xia, C.; Chen, Y.; Jiang, T.; Hu, Y.; Gao, Y. Resveratrol Synergistically Promotes BMP9-Induced Osteogenic Differentiation of Mesenchymal Stem Cells. Stem Cells Int. 2022, 2022, 8124085. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, K.; Bandyopadhyay, A.; Harfe, B.D.; Cox, K.; Kakar, S.; Gerstenfeld, L.; Einhorn, T.; Tabin, C.J.; Rosen, V. BMP2 activity, although dispensable for bone formation, is required for the initiation of fracture healing. Nat. Genet. 2006, 38, 1424–1429. [Google Scholar] [CrossRef]
- Rosen, V. BMP2 signaling in bone development and repair. Cytokine Growth Factor Rev. 2009, 20, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Qi, X.; Wu, X.; Lin, S.; Shi, J.; Zhang, W.; Jiang, X. Regulating macrophage-MSC interaction to optimize BMP-2-induced osteogenesis in the local microenvironment. Bioact. Mater 2023, 25, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.; Zhang, G.; Tan, Y.H. Calcitonin gene-related peptide stimulates BMP-2 expression and the differentiation of human osteoblast-like cells in vitro. Acta Pharmacol. Sin. 2013, 34, 1467–1474. [Google Scholar] [CrossRef]
- Wang, L.; Shi, X.; Zhao, R.; Halloran, B.P.; Clark, D.J.; Jacobs, C.R.; Kingery, W.S. Calcitonin-gene-related peptide stimulates stromal cell osteogenic differentiation and inhibits RANKL induced NF-kappaB activation, osteoclastogenesis and bone resorption. Bone 2010, 46, 1369–1379. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Zhang, S.J.; Wang, X.D.; Yang, Z.H.; Sun, Y.N.; Gupta, A.; Hou, R.; Lei, D.L.; Hu, K.J.; Ye, W.M.; et al. Calcitonin gene-related peptide enhances osteogenic differentiation and recruitment of bone marrow mesenchymal stem cells in rats. Exp. Ther. Med. 2019, 18, 1039–1046. [Google Scholar] [CrossRef]
- Zhou, R.; Yuan, Z.; Liu, J.; Liu, J. Calcitonin gene-related peptide promotes the expression of osteoblastic genes and activates the WNT signal transduction pathway in bone marrow stromal stem cells. Mol. Med. Rep. 2016, 13, 4689–4696. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, J.; Ruan, Y.C.; Yu, M.K.; O’Laughlin, M.; Wise, H.; Chen, D.; Tian, L.; Shi, D.; Wang, J.; et al. Implant-derived magnesium induces local neuronal production of CGRP to improve bone-fracture healing in rats. Nat. Med. 2016, 22, 1160–1169. [Google Scholar] [CrossRef]
- Appelt, J.; Baranowsky, A.; Jahn, D.; Yorgan, T.; Köhli, P.; Otto, E.; Farahani, S.K.; Graef, F.; Fuchs, M.; Herrera, A.; et al. The neuropeptide calcitonin gene-related peptide alpha is essential for bone healing. EBioMedicine 2020, 59, 102970. [Google Scholar] [CrossRef] [PubMed]
- Onuoha, G.N.; Alpar, E.K. Elevation of plasma CGRP and SP levels in orthopedic patients with fracture neck of femur. Neuropeptides 2000, 34, 116–120. [Google Scholar] [CrossRef]
- Takahashi, N.; Matsuda, Y.; Sato, K.; de Jong, P.R.; Bertin, S.; Tabeta, K.; Yamazaki, K. Neuronal TRPV1 activation regulates alveolar bone resorption by suppressing osteoclastogenesis via CGRP. Sci. Rep. 2016, 6, 29294. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, K.; Hirukawa, K.; Nakamura, H.; Togari, A. Inhibitory effect of CGRP on osteoclast formation by mouse bone marrow cells treated with isoproterenol. Neurosci. Lett. 2005, 379, 47–51. [Google Scholar] [CrossRef]
- Akopian, A.; Demulder, A.; Ouriaghli, F.; Corazza, F.; Fondu, P.; Bergmann, P. Effects of CGRP on human osteoclast-like cell formation: A possible connection with the bone loss in neurological disorders. Peptides 2000, 21, 559–564. [Google Scholar] [CrossRef]
- Toda, M.; Suzuki, T.; Hosono, K.; Kurihara, Y.; Kurihara, H.; Hayashi, I.; Kitasato, H.; Hoka, S.; Majima, M. Roles of calcitonin gene-related peptide in facilitation of wound healing and angiogenesis. Biomed. Pharmacother. 2008, 62, 352–359. [Google Scholar] [CrossRef]
- Aoki, M.; Tamai, K.; Saotome, K. Substance P- and calcitonin gene-related peptide-immunofluorescent nerves in the repair of experimental bone defects. Int. Orthop. 1994, 18, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.C.; Chen, S.; Zheng, L.; Qin, L. Angiogenesis Assays for the Evaluation of Angiogenic Properties of Orthopaedic Biomaterials - A General Review. Adv. Healthc. Mater. 2017, 6, 1600434. [Google Scholar] [CrossRef]
- Maeda, Y.; Miwa, Y.; Sato, I. Expression of CGRP, vasculogenesis and osteogenesis associated mRNAs in the developing mouse mandible and tibia. Eur. J. Histochem. 2017, 61, 2750. [Google Scholar] [CrossRef] [PubMed]
- Bidegain, M.; Roos, B.A.; Hill, E.L.; Howard, G.A.; Balkan, W. Calcitonin gene-related peptide (CGRP) in the developing mouse limb. Endocr. Res. 1995, 21, 743–755. [Google Scholar] [CrossRef] [PubMed]
- Mi, J.; Xu, J.; Yao, H.; Li, X.; Tong, W.; Li, Y.; Dai, B.; He, X.; Chow, D.; Li, G.; et al. Calcitonin Gene-Related Peptide Enhances Distraction Osteogenesis by Increasing Angiogenesis. Tissue Eng. Part A 2021, 27, 87–102. [Google Scholar] [CrossRef]
- Zheng, S.; Li, W.; Xu, M.; Bai, X.; Zhou, Z.; Han, J.; Shyy, J.Y.; Wang, X. Calcitonin gene-related peptide promotes angiogenesis via AMP-activated protein kinase. Am. J. Physiol. Cell Physiol. 2010, 299, C1485–C1492. [Google Scholar] [CrossRef] [PubMed]
- Mapp, P.I.; McWilliams, D.F.; Turley, M.J.; Hargin, E.; Walsh, D.A. A role for the sensory neuropeptide calcitonin gene-related peptide in endothelial cell proliferation in vivo. Br. J. Pharmacol. 2012, 166, 1261–1271. [Google Scholar] [CrossRef]
- Toda, M.; Suzuki, T.; Hosono, K.; Hayashi, I.; Hashiba, S.; Onuma, Y.; Amano, H.; Kurihara, Y.; Kurihara, H.; Okamoto, H.; et al. Neuronal system-dependent facilitation of tumor angiogenesis and tumor growth by calcitonin gene-related peptide. Proc. Natl. Acad. Sci. USA 2008, 105, 13550–13555. [Google Scholar] [CrossRef]
- Wu, J.; Liu, S.; Wang, Z.; Ma, S.; Meng, H.; Hu, J. Calcitonin gene-related peptide promotes proliferation and inhibits apoptosis in endothelial progenitor cells via inhibiting MAPK signaling. Proteome Sci. 2018, 16, 18. [Google Scholar] [CrossRef]
- Bo, Y.; Yan, L.; Gang, Z.; Tao, L.; Yinghui, T. Effect of calcitonin gene-related peptide on osteoblast differentiation in an osteoblast and endothelial cell co-culture system. Cell Biol. Int. 2012, 36, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.Y.; Guo, J.; Yang, W.F.; Tao, Z.Y.; Lan, X.; Wang, L.; Xu, J.; Qin, L.; Su, Y.X. Biodegradable magnesium implant enhances angiogenesis and alleviates medication-related osteonecrosis of the jaw in rats. J. Orthop. Translat. 2022, 33, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Chen, H.; Jiang, Y.; Yuan, Y.; Zhang, Q.; Guo, Q.; Gong, P. CGRP regulates the dysfunction of peri-implant angiogenesis and osseointegration in streptozotocin-induced diabetic rats. Bone 2020, 139, 115464. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, J.; Mi, J.; He, X.; Pan, Q.; Zheng, L.; Zu, H.; Chen, Z.; Dai, B.; Li, X.; et al. Biodegradable magnesium combined with distraction osteogenesis synergistically stimulates bone tissue regeneration via CGRP-FAK-VEGF signaling axis. Biomaterials 2021, 275, 120984. [Google Scholar] [CrossRef]
- Brown, B.N.; Badylak, S.F. Expanded applications, shifting paradigms and an improved understanding of host-biomaterial interactions. Acta Biomater. 2013, 9, 4948–4955. [Google Scholar] [CrossRef]
- Gibon, E.; Lu, L.; Goodman, S.B. Aging, inflammation, stem cells, and bone healing. Stem Cell Res. Ther. 2016, 7, 44. [Google Scholar] [CrossRef]
- Yang, L.; Sakurai, T.; Kamiyoshi, A.; Ichikawa-Shindo, Y.; Kawate, H.; Yoshizawa, T.; Koyama, T.; Iesato, Y.; Uetake, R.; Yamauchi, A.; et al. Endogenous CGRP protects against neointimal hyperplasia following wire-induced vascular injury. J. Mol. Cell. Cardiol. 2013, 59, 55–66. [Google Scholar] [CrossRef]
- Feng, Y.; Tang, Y.; Guo, J.; Wang, X. Inhibition of LPS-induced TNF-alpha production by calcitonin gene-related peptide (CGRP) in cultured mouse peritoneal macrophages. Life Sci. 1997, 61, PL 281–PL 287. [Google Scholar] [CrossRef]
- Millet, I.; Vignery, A. The neuropeptide calcitonin gene-related peptide inhibits TNF-alpha but poorly induces IL-6 production by fetal rat osteoblasts. Cytokine 1997, 9, 999–1007. [Google Scholar] [CrossRef]
- Niedermair, T.; Straub, R.H.; Brochhausen, C.; Grässel, S. Impact of the Sensory and Sympathetic Nervous System on Fracture Healing in Ovariectomized Mice. Int. J. Mol. Sci. 2020, 21, 405. [Google Scholar] [CrossRef]
- Loi, F.; Córdova, L.A.; Zhang, R.; Pajarinen, J.; Lin, T.H.; Goodman, S.B.; Yao, Z. The effects of immunomodulation by macrophage subsets on osteogenesis in vitro. Stem. Cell Res. Ther. 2016, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Jiang, Y.; Wang, B.; Guo, Y.; Gong, P.; Xiang, L. Deficiency of Calcitonin Gene-Related Peptide Affects Macrophage Polarization in Osseointegration. Front Physiol. 2020, 11, 733. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wu, B.; Yuan, Y.; Zhang, X.; Guo, Y.; Gong, P.; Xiang, L. CGRP-modulated M2 macrophages regulate osteogenesis of MC3T3-E1 via Yap1. Arch. Biochem. Biophys. 2021, 697, 108697. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, M.; Rhee, C.; Utsunomiya, T.; Zhang, N.; Ueno, M.; Yao, Z.; Goodman, S.B. Modulation of the Inflammatory Response and Bone Healing. Front. Endocrinol. (Lausanne) 2020, 11, 386. [Google Scholar] [CrossRef]
- Chen, J.; Ma, G.; Liu, W.; Liu, Y.; Ding, Y. The influence of the sensory neurotransmitter calcitonin gene-related peptide on bone marrow mesenchymal stem cells from ovariectomized rats. J. Bone Miner. Metab. 2017, 35, 473–484. [Google Scholar] [CrossRef]
- Zhang, Q.; Guo, Y.; Yu, H.; Tang, Y.; Yuan, Y.; Jiang, Y.; Chen, H.; Gong, P.; Xiang, L. Receptor activity-modifying protein 1 regulates the phenotypic expression of BMSCs via the Hippo/Yap pathway. J. Cell. Physiol. 2019, 234, 13969–13976. [Google Scholar] [CrossRef]
- Michelangeli, V.P.; Fletcher, A.E.; Allan, E.H.; Nicholson, G.C.; Martin, T.J. Effects of calcitonin gene-related peptide on cyclic AMP formation in chicken, rat, and mouse bone cells. J. Bone Miner. Res. 1989, 4, 269–272. [Google Scholar] [CrossRef]
- Villa, I.; Melzi, R.; Pagani, F.; Ravasi, F.; Rubinacci, A.; Guidobono, F. Effects of calcitonin gene-related peptide and amylin on human osteoblast-like cells proliferation. Eur. J. Pharmacol. 2000, 409, 273–278. [Google Scholar] [CrossRef]
- Mrak, E.; Guidobono, F.; Moro, G.; Fraschini, G.; Rubinacci, A.; Villa, I. Calcitonin gene-related peptide (CGRP) inhibits apoptosis in human osteoblasts by β-catenin stabilization. J. Cell. Physiol. 2010, 225, 701–708. [Google Scholar] [CrossRef]
- Owan, I.; Ibaraki, K. The role of calcitonin gene-related peptide (CGRP) in macrophages: The presence of functional receptors and effects on proliferation and differentiation into osteoclast-like cells. Bone Miner. 1994, 24, 151–164. [Google Scholar] [CrossRef]
- Yoo, Y.M.; Kwag, J.H.; Kim, K.H.; Kim, C.H. Effects of neuropeptides and mechanical loading on bone cell resorption in vitro. Int. J. Mol. Sci. 2014, 15, 5874–5883. [Google Scholar] [CrossRef] [PubMed]
- Calland, J.W.; Harris, S.E.; Carnes, D.L., Jr. Human pulp cells respond to calcitonin gene-related peptide in vitro. J. Endod. 1997, 23, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Yang, Q.; Xiong, W.; Li, G.H.; Liao, H.; Xiao, J.; Li, F. Effect of CGRP-adenoviral vector transduction on the osteoblastic differentiation of rat adipose-derived stem cells. PLoS ONE 2013, 8, e72738. [Google Scholar] [CrossRef] [PubMed]
- Seifert, H.; Sawchenko, P.; Chesnut, J.; Rivier, J.; Vale, W.; Pandol, S.J. Receptor for calcitonin gene-related peptide: Binding to exocrine pancreas mediates biological actions. Am. J. Physiol. 1985, 249, G147–G151. [Google Scholar] [CrossRef]
- Edvinsson, L.; Fredholm, B.B.; Hamel, E.; Jansen, I.; Verrecchia, C. Perivascular peptides relax cerebral arteries concomitant with stimulation of cyclic adenosine monophosphate accumulation or release of an endothelium-derived relaxing factor in the cat. Neurosci. Lett. 1985, 58, 213–217. [Google Scholar] [CrossRef]
- Takamori, M.; Yoshikawa, H. Effect of calcitonin gene-related peptide on skeletal muscle via specific binding site and G protein. J. Neurol. Sci. 1989, 90, 99–109. [Google Scholar] [CrossRef]
- Main, M.J.; Brown, J.; Brown, S.; Fraser, N.J.; Foord, S.M. The CGRP receptor can couple via pertussis toxin sensitive and insensitive G proteins. FEBS Lett. 1998, 441, 6–10. [Google Scholar] [CrossRef]
- Dickerson, I.M. Role of CGRP-receptor component protein (RCP) in CLR/RAMP function. Curr. Protein Pept. Sci. 2013, 14, 407–415. [Google Scholar] [CrossRef]
- Cottrell, G.S. CGRP Receptor Signalling Pathways. Handb. Exp. Pharmacol. 2019, 255, 37–64. [Google Scholar]
- Weston, C.; Winfield, I.; Harris, M.; Hodgson, R.; Shah, A.; Dowell, S.J.; Mobarec, J.C.; Woodlock, D.A.; Reynolds, C.A.; Poyner, D.R.; et al. Receptor Activity-modifying Protein-directed G Protein Signaling Specificity for the Calcitonin Gene-related Peptide Family of Receptors. J. Biol. Chem. 2016, 291, 21925–21944. [Google Scholar] [CrossRef]
- Hankenson, K.D.; Gagne, K.; Shaughnessy, M. Extracellular signaling molecules to promote fracture healing and bone regeneration. Adv. Drug Deliv. Rev. 2015, 94, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Hoff, A.O.; Catala-Lehnen, P.; Thomas, P.M.; Priemel, M.; Rueger, J.M.; Nasonkin, I.; Bradley, A.; Hughes, M.R.; Ordonez, N.; Cote, G.J.; et al. Increased bone mass is an unexpected phenotype associated with deletion of the calcitonin gene. J. Clin. Invest. 2002, 110, 1849–1857. [Google Scholar] [CrossRef] [PubMed]
- Kauther, M.D.; Neuerburg, C.; Wefelnberg, F.; Bachmann, H.S.; Schlepper, R.; Hilken, G.; Broecker-Preuss, M.; Grabellus, F.; Schilling, A.F.; Jäger, M.; et al. RANKL-associated suppression of particle-induced osteolysis in an aged model of Calcitonin and α-CGRP deficiency. Biomaterials 2013, 34, 2911–2919. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Li, L.; Cui, X.; Tang, Z.; Wei, X.; Pan, H.; Li, B. Enhanced proliferation and differentiation effects of a CGRP- and Sr-enriched calcium phosphate cement on bone mesenchymal stem cells. J. Appl. Biomater. Funct. Mater. 2016, 14, e431–e440. [Google Scholar] [CrossRef]
- Yu, X.; Liu, S.; Chen, X.; Du, Y.; Yin, X.; Du, Y.; Li, S. Calcitonin gene related peptide gene-modified rat bone mesenchymal stem cells are effective seed cells in tissue engineering to repair skull defects. Histol. Histopathol. 2019, 34, 1229–1241. [Google Scholar]
- Irie, K.; Hara-Irie, F.; Ozawa, H.; Yajima, T. Calcitonin gene-related peptide (CGRP)-containing nerve fibers in bone tissue and their involvement in bone remodeling. Microsc. Res. Tech. 2002, 58, 85–90. [Google Scholar] [CrossRef]
- Gao, F.; Lv, T.R.; Zhou, J.C.; Qin, X.D. Effects of obesity on the healing of bone fracture in mice. J. Orthop Surg Res. 2018, 13, 145. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, B.; Yang, Y.; Peng, B.; Ye, R. PLGA Containing Human Adipose-Derived Stem Cell-Derived Extracellular Vesicles Accelerates the Repair of Alveolar Bone Defects via Transfer of CGRP. Oxid. Med. Cell Longev. 2022, 2022, 4815284. [Google Scholar] [CrossRef]
- Zhao, D.; Witte, F.; Lu, F.; Wang, J.; Li, J.; Qin, L. Current status on clinical applications of magnesium-based orthopaedic implants: A review from clinical translational perspective. Biomaterials 2017, 112, 287–302. [Google Scholar] [CrossRef]
- Ding, Y.; Yan, Q.; Ruan, J.W.; Zhang, Y.Q.; Li, W.J.; Zhang, Y.J.; Li, Y.; Dong, H.; Zeng, Y.S. Electro-acupuncture promotes survival, differentiation of the bone marrow mesenchymal stem cells as well as functional recovery in the spinal cord-transected rats. BMC Neurosci. 2009, 10, 35. [Google Scholar] [CrossRef]
- Bucelli, R.C.; Gonsiorek, E.A.; Kim, W.Y.; Bruun, D.; Rabin, R.A.; Higgins, D.; Lein, P.J. Statins decrease expression of the proinflammatory neuropeptides calcitonin gene-related peptide and substance P in sensory neurons. J. Pharmacol. Exp. Ther. 2008, 324, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Tuzmen, C.; Campbell, P.G. Crosstalk between neuropeptides SP and CGRP in regulation of BMP2-induced bone differentiation. Connect. Tissue Res. 2018, 59, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, L. Role of CGRP in Migraine. Handb. Exp. Pharmacol. 2019, 255, 121–130. [Google Scholar]
- Sharma, K.; Sharma, K.K.; Sharma, A.; Jain, R. Peptide-based drug discovery: Current status and recent advances. Drug Discov. Today 2023, 28, 103464. [Google Scholar] [CrossRef]
- Fosgerau, K.; Hoffmann, T. Peptide therapeutics: Current status and future directions. Drug Discov. Today 2015, 20, 122–128. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Target Spot | Name | Type | Current Status |
|---|---|---|---|
| CGRPR | Olcegepant [35] | Nonpeptide | Phase II Clinical Trial |
| CGRPR | Telcegepant [36] | Nonpeptide | Phase III Clinical Trial |
| CGRPR | MK-3207 [37] | Nonpeptide | Phase II Clinical Trial |
| CGRPR | Zavegapant [38] | Nonpeptide | Phase II/III Clinical Trial |
| CGRPR | BI 44,370 [39] | Nonpeptide | Phase II Clinical Trial |
| CGRPR | Erenumab [40] | Monoclonal antibody | FDA approved in 2018 |
| CGRPR | Rimegepant [41] | Nonpeptide | FDA approved in 2020 |
| CGRPR | Ubrogepant [42] | Nonpeptide | FDA approved in 2020 |
| CGRPR | Atogepant [43] | Nonpeptide | FDA approved in 2022 |
| CGRP | Galcanezumab [44] | Monoclonal antibody | FDA approved in 2018 |
| CGRP | Fremanezumab [45] | Monoclonal antibody | FDA approved in 2018 |
| CGRP | Eptinezumab [46] | Monoclonal antibody | FDA approved in 2020 |
| Cell Category | Author | Year | Signal Pathway | Experimental Results |
|---|---|---|---|---|
| BMSCs | Liping Wang [72] | 2010 | RANKL/NF-κB | Enhance the osteogenic differentiation ability and inhibit osteoclast formation |
| Wei Liang [63] | 2015 | ALP, collagen I, BMP2, osteonectin, and RUNX2 are upregulated, leading to increased proliferation and osteogenic differentiation | ||
| Ri Zhou [74] | 2016 | Wnt/β-catenin | Enhance the osteogenic differentiation ability | |
| Jie Chen [105] | 2017 | The proliferation and osteogenic differentiation abilities are enhanced, and the adipogenic differentiation is inhibited in a dose-dependent manner | ||
| Sen Jia [73] | 2019 | The expression of ALP and RUNX2 is upregulated, and the proliferation and migration abilities are enhanced | ||
| Qin Zhang [106] | 2019 | Hippo/Yap | Upregulate ALP, RUNX2, and OPN | |
| Yanjun Guo [93] | 2020 | Enhanced expression of VEGF, ALP, and OPN and reduced production of reactive oxygen species (ROS) induced by high glucose | ||
| Hang Li [54] | 2021 | The osteogenic differentiation ability is enhanced, and the adipogenic differentiation ability is weakened | ||
| Osteoblast | Michelangeli [107] | 1989 | cAMP | Upregulate cAMP |
| Vignery [62] | 1996 | cAMP | Insulin-like growth factor expression is upregulated, and the osteogenic differentiation ability is elevated | |
| Millet [99] | 1997 | cAMP | Inhibit the TNF-α production stimulated by lipopolysaccharide and IL-1, but promote IL6 expression | |
| Villa [108] | 2000 | cAMP | Enhance the proliferation ability | |
| Mrak [109] | 2010 | Wnt/β-catenin | Inhibit apoptosis | |
| Yang Bo [91] | 2013 | The expression of OC, ALP, and collagen is upregulated, and the osteogenic ability differentiation is increased | ||
| Haitao He [64] | 2016 | RANKL/NF-κB | ATF4 and OC expression is upregulated, and the osteogenic differentiation ability is enhanced | |
| Macrophage | Owan [110] | 1994 | cAMP | Inhibit osteoclast formation |
| Feng [98] | 1997 | cAMP | Inhibit LPS-induced TNF-α upregulation | |
| Yeong-Min [111] | 2014 | RANKL/NF-κB | Upregulate OPG expression and inhibit bone resorption | |
| Qin Zhang [103] | 2021 | Hippo/Yap | Inhibit and promote the osteogenic factor secretion in M2 macrophages in the early and late stages, respectively | |
| Osteoclast | Akopian [80] | 2000 | cAMP | Inhibit osteoclast formation in a dose-dependent manner |
| Ishizuka [79] | 2005 | RANKL/NF-κB | Inhibit osteoclast formation | |
| EPCs | Haegerstrand [50] | 1990 | cAMP | Enhance the proliferation ability |
| Shuai Zheng [87] | 2010 | cAMP | Activate AMPK-eNOS and enhance angiogenesis | |
| Yang Bo [91] | 2013 | Upregulate OC, ALP, and COL expression in vascular endothelial cells and osteoblast co-culture system | ||
| Jianqun Wu [90] | 2018 | MAPK | Enhance the proliferation ability and inhibit apoptosis | |
| Jie Mi [86] | 2021 | PI3K/AKT | The proliferation ability and angiogenesis are enhanced. The osteogenic differentiation ability of BMSCs is increased | |
| Ye Li [94] | 2021 | FAK/VEGF | Enhanced migration ability, promote FAK phosphorylation of and upregulate VEGF expression | |
| PDSCs | Yifeng Zhang [75] | 2016 | cAMP | SP7 and ALP expression is upregulated, and proliferation and osteogenic differentiation ability are enhanced |
| DPSCs | Calland [112] | 1997 | cAMP | BMP2 expression is upregulated, and the osteogenic differentiation ability is elevated |
| MG63 | Gang Tian [71] | 2013 | cAMP | BMP2 expression is upregulated, and the osteogenic differentiation ability is enhanced |
| ADSCs | Zhong Fang [113] | 2013 | ALP expression in the overexpressed CGRP group is upregulated, and the cell proliferation and osteogenic differentiation ability are enhanced | |
| DRG | Jie Mi [15] | 2021 | cAMP | Electrical stimulation promotes the synthesis and release of CGRP in DRG and enhances the H-type blood vessel formation and osteoporotic fracture healing |
| Model | Author | Year | Modeling Method | Phenotype |
|---|---|---|---|---|
| Gene editing | Ballica [58] | 1999 | Construction of mouse model of CGRP overexpression | Inhibit osteoclasts, stimulate insulin-like growth factor, and inhibit the tumor necrosis factor-α production |
| Hoff [122] | 2002 | Construction of CGRP knockout mouse model | The bone mass of gene-knockout mice was maintained after ovariectomy, and that of gene-knockout wild-type mice decreased within two months. | |
| Schinke [59] | 2004 | Construction of CALCA and CGRP knockout mouse model | CALCA knockout mice revealed a high bone mass, while CGRP knockout mice showed a low bone mass. | |
| Toda [81] | 2008 | Construction of CGRP knockout mouse model | The expression of vascular endothelial growth factor within wound granulation tissue of CGRP knockout mice decreased. Angiogenesis and wound closure was significantly inhibited. | |
| Lei Yang [97] | 2013 | Construction of CGRP knockout mouse model | CGRP can inhibit oxidative stress and the proliferation of vascular smooth muscle cells induced by vascular injury. | |
| Kauther [123] | 2013 | Construction of CGRP knockout mouse model | In mice, OPG and OCN increased significantly, osteoclasts elevated, and RANKL decreased significantly. | |
| Takahashi [78] | 2016 | Construction of TRPV1 knockout mouse model | TRPV1 affects osteoclast formation by CGRP regulation | |
| Niedermair [100] | 2020 | Construction of CGRP knockout mouse model | CGRP relieved pain and promoted the polarization of M2 macrophages but did not affect bone maturation. | |
| Appelt [76] | 2020 | Construction of CGRP knockout mouse model | The number of bone-forming osteoblasts in CGRP-deficient mice decreased significantly, and bone healing was poor. | |
| Bone graft | Zhong Fang [113] | 2013 | Implantation of CGRP overexpressed ADSCs/β-TCP Bracket | ADSCs overexpressing the CGRP/β-TCP stent promotes bone repair |
| Wei Liang [124] | 2016 | Implantation of CPC containing CGRP and Sr | CPC containing CGRP and Sr promotes bone repair among osteoporotic rats | |
| Xijiao Yu [125] | 2019 | Implantation of CGRP overexpressed collagen scaffolds in BMSCs | BMScs collagen scaffold overexpressing CGRP leads to skull repair in rats | |
| Sen Jia [73] | 2019 | Construction of a rat model of distraction osteogenesis | CGRP enhances new bone formation by elevating the migration and differentiation of bone marrow stromal cells. | |
| Ye Li [94] | 2021 | Construction of a rat model of distraction osteogenesis | Magnesium-containing intramedullary nail promotes bone defect repair in rats by the upregulating CGRP/FAK/VEGF pathway. | |
| Wangyong Zhu [92] | 2021 | Construction of a rat model of drug-related osteonecrosis | Magnesium grafts promote angiogenesis and bone repair by regulating VEGF and CGRP, thereby alleviating drug-related osteonecrosis. | |
| Injection of drugs | Mapp [88] | 2012 | Normal knee joint | Promote angiogenesis |
| Yanjun Guo [93] | 2020 | Construction of diabetic rat model | CGRP injection overexpressed BMSCs can elevate ALP activity and promote mRNA and protein expression of VEGF, ALP, and OPN | |
| Hang Li | 2021 | Construction of senile mouse model and osteoporotic mouse model | Promote bone formation in aged mice, decrease fat accumulation, and delay osteoporosis occurrence in mice. | |
| Other | Aoki [82] | 1994 | Tibial bone defect model | Local blood flow and callus formation increased. |
| Irie [126] | 2002 | Normal bone tissue | There are abundant CGRP-positive nerve fibers around the bone tissue. | |
| Maeda [84] | 2017 | Normal fetal rat bone tissue | CGRP is essential in osteogenesis and angiogenesis in bone development. | |
| Feng Gao [127] | 2018 | Construction of obese mouse model | In obese mice, CGRP, FGF, and TGF-β levels decreased, while TNF-α levels increased, and bone repair was delayed. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Qin, H.; Deng, J.; Xu, H.; Liu, S.; Weng, J.; Zeng, H. Research Progress in Calcitonin Gene-Related Peptide and Bone Repair. Biomolecules 2023, 13, 838. https://doi.org/10.3390/biom13050838
Wang Q, Qin H, Deng J, Xu H, Liu S, Weng J, Zeng H. Research Progress in Calcitonin Gene-Related Peptide and Bone Repair. Biomolecules. 2023; 13(5):838. https://doi.org/10.3390/biom13050838
Chicago/Turabian StyleWang, Qichang, Haotian Qin, Jiapeng Deng, Huihui Xu, Su Liu, Jian Weng, and Hui Zeng. 2023. "Research Progress in Calcitonin Gene-Related Peptide and Bone Repair" Biomolecules 13, no. 5: 838. https://doi.org/10.3390/biom13050838