Abstract
Adipose tissue is composed of adipocytes, stromal vascular fraction, nerves, surrounding immune cells, and the extracellular matrix. Under various physiological or pathological conditions, adipose tissue shifts cellular composition, lipid storage, and organelle dynamics to respond to the stress; this remodeling is called “adipose tissue plasticity”. Adipose tissue plasticity includes changes in the size, species, number, lipid storage capacity, and differentiation function of adipocytes, as well as alterations in the distribution and cellular composition of adipose tissue. This plasticity has a major role in growth, obesity, organismal protection, and internal environmental homeostasis. Moreover, certain thresholds exist for this plasticity with significant individualized differences. Here, we comprehensively elaborate on the specific connotation of adipose tissue plasticity and the relationship between this plasticity and the development of many diseases. Meanwhile, we summarize possible strategies for treating obesity in response to adipose tissue plasticity, intending to provide new insights into the dynamic changes in adipose tissue and contribute new ideas to relevant clinical problems.
1. Introduction
Adipose tissue (AT) is a highly active, flexible organ with great remodeling ability and functional pleiotropism [1], and its significance in systemic physiology has recently been highlighted [2]. Because AT includes mature adipocytes, blood vessels, neurons, fibroblasts, and different immune cells such as macrophages, the extracellular matrix (ECM), adipose stem cells (ASCs), and preadipocytes, it is considered a tissue [3]. These many elements work in sync in AT to sustain AT homeostasis. AT is divided into white and brown ATs (WAT and BAT, respectively), and beige AT is the portion of WAT that resembles BAT. WAT and BAT have unique functions and are closely related during the body’s adaptation to changes in energy demand. WAT, including subcutaneous AT (SAT) and visceral AT (VAT), stores energy in the form of lipid droplets. These droplets protect the remaining body’s organs from the toxicity of ectopic lipid deposits [4]. Thermogenic AT (brown and beige) generates heat by oxidizing fatty acids, etc., thereby facilitating energy expenditure and obesity prevention [5]. The majority of thermogenic adipocyte heat production depends on the uncoupling protein 1 (UCP1) pathway in the inner mitochondrial membrane, and free fatty acids (FFAs), glucose, lactate, and ketone bodies act as thermogenic fuels [6,7].
Excessive fat storage leads to individuals being affected by overweight or obesity. This results in a series of health disorders, including metabolic syndrome, stroke, coronary heart disease, and type 2 diabetes mellitus (T2DM) [8]. The global prevalence of obesity is continuously increasing. At present, approximately one-third and one-tenth of the global population are affected by overweight and obesity, respectively [9].
Recent advancements in technology such as single-cell RNA sequencing (scRNA-seq) and single-nucleus sequencing (snRNA-seq) have facilitated our understanding of the heterogeneity and functionality of AT depots [10,11,12]. We herein present a new and complete definition of AT plasticity for the first time. According to us, AT plasticity is defined as the active adaptability of AT to undergo significant changes in its various tissue components under physiological or pathological conditions up to a certain threshold. These changes include those in adipocyte size and number, blood vessels and nerves, infiltrating immune cells, adipokine secretion, differentiation and dedifferentiation capacity, etc. We also highlight the multifaceted interpretation of AT plasticity and the presence of certain thresholds as well as individual variability for this plasticity. We also elaborate on possible strategies related to obesity treatment.
2. Plasticity in Adipocyte Size
Adipocytes can enlarge or contract to adjust to regional and systemic metabolic changes (Figure 1A). This size change is accomplished by adipocyte hypertrophy or atrophy [13]. Adipocytes are highly plastic, and their size can increase from 20 μm to hundreds of micrometers in diameter during hypertrophy, and this mainly depends on the triglyceride content [14]. Adipocyte hypertrophy increases inflammation and cell death [15], and excessive adipocyte hypertrophy may be a stress that causes ongoing inflammation in AT [16]. The hypertrophy of subcutaneous abdominal adipocytes is a hallmark of increased WAT in people with obesity, which is positively associated with glucose tolerance and hyperinsulinemia and may increase T2DM risk [17].

Figure 1.
The plasticity in adipocyte size, number, and species. (A) Plasticity in the adipocyte size. Adipocyte size plasticity is accomplished by hypertrophy or atrophy. (B) Plasticity in the adipocyte number. The increase in the number of adipocytes occurs through cell proliferation, while the decrease in the number of adipocytes is promoted by apoptosis. (C) Plasticity in adipocyte species. The white adipose tissues and brown adipose tissues can be interconverted under certain conditions (such as browning and whitening).
Adipocyte size is regulated by various environmental and physiological factors. Prolonged exposure to mild cold enlarges BAT mass and activity, which is restored when AT is exposed to a warm environment [18]. Research has demonstrated a noticeable increase in visceral fat, particularly in the epididymal WAT, in mice with advancing age. Morphological investigations have corroborated that this augmentation primarily arises from the enlargement of adipocytes. Certain dietary interventions, such as a 72 h fasting regimen, have been observed to induce a significant reduction in adipocyte size among mice, with more pronounced effects observed in younger cohorts. Following a 72 h refeeding period, adipocytes in younger mice reverted to their initial dimensions, whereas those in adult and senescent mice exhibited sustained but marginally reduced sizes compared to pre-fasting levels [19]. An organism’s sex and AT location also seem to regulate adipocyte size plasticity. Females have smaller adipocytes in the omentum than in the SAT, whereas males typically have larger adipocytes and have little difference in adipocyte size in both sites [20]. This might be related to sex hormones, as the aforementioned differences in AT morphology decrease in women after menopause, more closely resembling the expression pattern in men [21].
In vivo, adipocyte hypertrophy and its degree are also regulated by local and systemic factors. As adipocyte size increases, the adipocytes come in contact with adjacent cells and ECM components and are subjected to increased mechanical stress and decreased oxygen diffusion. This leads to hypoxia and AT inflammation at the threshold, thus preventing the increase in adipocyte size [22]. Adipocyte size is suggested to be regulated by volume-sensitive adipocyte membrane proteins, such as SWELL11 [23]. The expression of inflammation-related genes NF-κB and tumor necrosis factor (TNF)-γ is positively correlated with adipocyte size. This reflects that increased adipocyte size is correlated with increased inflammatory responses [24]. The expression of immune-related genes such as serum amyloid A is also significantly higher in large adipocytes [25]. Adipokine secretion also has a role in size regulation, with extra-large adipocytes (>127 μm) exhibiting an increased secretion of pro-inflammatory adipocytokines interleukin-6 (IL-6), monocyte chemotactic protein-1 (MCP-1), and IL-8 compared with small adipocytes (<73 μm) [15], Increased expression of lipid droplet surface proteins Cell death induces DFFA-like effector protein A (CIDEA) and Fat-specific Protein 27 in the SAT may increase adipocyte size by increasing triglyceride storage [26]. The metabolic rhythms of peroxisome proliferator-activated receptor γ (PPARγ) acetylation also influences adipocyte size [27].
3. Plasticity in Adipocyte Number
To increase or decrease adipocyte number, new cells must be produced de novo, and existing cells must be destroyed (Figure 1B) [13]. Although most adipose growth occurs during childhood and adolescence, with fluctuations in energy balance, WAT can continue to undergo expansion during adulthood [28].
The increase in adipocyte number occurs mainly through cell proliferation, often also called adipogenesis, involving the transition from multipotent mesenchymal stem cells (MSCs) to preadipocytes and then to adipocytes [29]. The specific molecular regulatory mechanisms are mentioned subsequently (8.1). As confirmed in many experiments, AT expansion occurs primarily from adipocyte proliferation, thereby preventing metabolic diseases by maintaining the function of adipocytes and adequate lipid storage capacity within them. Lu et al. induced proliferative SAT expansion by injecting an adipose cocktail in mice, thereby demonstrating improved glucose tolerance and insulin sensitivity [30]. PPARγ agonist thiazolidinediones (TZD) improve systemic insulin sensitivity through proliferative SAT expansion despite weight gain [31].
Adipocyte hyperplasia rather than adipocyte hypertrophy manifests increased VAT mass in the population affected by obesity [32]. The proliferative capacity of VAT is greater than that of SAT. Although adipocyte proliferation in SAT increases during diet-induced obesity, adipogenesis may be limited to VAT at obesity onset [33].
Adipocyte numbers may decrease through apoptosis. Adipocyte apoptosis occurs under physiological conditions, with increased apoptotic marker expression in mature adipocytes in a thermoneutral (30 °C) environment compared with controls (22 °C) [34]. Untargeted metabolomics have revealed that apoptotic brown adipocytes release highly enriched purine metabolites, thereby enhancing the expression of thermogenic programs in healthy adipocytes [35]. Adenine nucleotide translocase 2 (ANT2) participates in adipocyte apoptosis, and ANT2 expression is downregulated during cold stimulation with a corresponding decrease in apoptosis ([36], p. 2). In people with morbid obesity, apoptosis increases with the exacerbation of metabolic-associated fatty liver disease (MAFLD) [37].
There is a strong link between changes in adipocyte size and their overall number. In SAT, the total number of adipocytes tends to decrease as their size increases, and fewer new adipocytes are formed in hypertrophic AT [38].
4. Plasticity of Adipocyte Species
4.1. Browning
Browning is defined as the transformation of WAT into tissue with BAT-like function and activity. It involves changes in the lipid droplet structure, robust mitochondrial biogenesis, and the upregulation of transcriptional programs supporting high levels of local fuel oxidation (Figure 1C) [39]. In healthy physiological conditions, BAT plays a crucial role in thermogenesis and energy expenditure, whereas WAT primarily functions as an energy reservoir. However, during obesity, WAT can become dysfunctional due to chronic inflammation, leading to impaired metabolic regulation. This dysfunction increases the risk of cardiovascular disease and T2DM. WAT browning is a potential therapeutic approach, as BAT activation prevents the deleterious effects of dysfunctional white adipocyte-induced lipid overflow [33,40].
4.1.1. Transcriptional and Epigenetic Regulation
Several proteins, including PPARγ coactivator 1α (PGC-1α) [41], PR-domain-containing 16 (PRDM16) [42], and early B-cell factor 2 [43], interact with PPARγ to regulate WAT browning. Many members of the bone morphogenetic protein (BMP) family, a subgroup of the transforming growth factor-β (TGF-β) superfamily, are involved in regulating browning. BMP4 promotes browning and improves insulin sensitivity [44], and BMP7 functions in WAT browning by increasing the expression of the transcriptional regulators PRDM16, PGC-1α, and PPARγ [45,46]. Experiments have demonstrated that the knockdown of SMAD3, which is part of the TGF-β signaling pathway, induces AT browning and prevents obesity and diabetes. The deletion of the transcription factor FoxO1 can lead to the same outcome [47]. This suggests that TGF-β is involved in browning [48]. Genetically inhibited Notch signaling may also promote WAT browning and inhibit obesity [49]. Many epigenetic regulatory molecules such as histone acetyltransferases, histone deacetylases, histone methyltransferases, and histone demethylases are associated with WAT browning [50]. miRNAs play a bidirectional role in the activation of BAT and beige adipose tissue [51]. Regarding positive regulation, under cold conditions, miR-455 is specifically expressed in BAT to activate brown adipocytes and adipose-specific miR-455 transgenic mice exhibited browning in the SAT [52]. miR-199a/214 [53], miR-155 [54], miR-327 [55], miR-19b [56], miR-133 [57], and miR-34a [58] are key negative regulators that inhibit the WAT-to-BAT conversion and beige adipocyte recruitment.
4.1.2. Endocrine Hormone Regulation
Catecholamines are browning-inducing hormones that act through β-adrenergic receptors (βARs) on adipocytes to promote browning and increase thermogenesis [59]. Under cold conditions, the sympathetic nervous system is activated and norepinephrine (NE) release increases, thereby binding and activating the βARs on adipocytes. βARs then interact with the GTP-binding protein GS to activate adenylate cyclase, thereby increasing cellular cAMP levels and activating cAMP-dependent protein kinase A (PKA). PKA phosphorylates and modifies lipid droplet-binding proteins A and B and some lipases, such as adipose triglyceride lipase and hormone-sensitive triglyceride lipase, to promote FFA and glycerol release [60,61]. The released FFA activates the UCP1 protein and acts as a substrate for UCP1-mediated thermogenesis [62]. The p38 pathway also has a regulatory role in the aforementioned thermogenesis [63]. Curcumin enhances the expression of β3-adrenergic receptor genes in SAT in mice, thereby increasing plasma NE levels and enhancing thermogenesis [64]. Of note, even in the absence of βARs, cold stimulation can induce beige adipocyte production to promote thermogenesis. This particular beige adipose tissue is known as glycolytic beige (g-beige) adipose tissue [65]. Thyroid hormones (THs) also induce WAT browning and activate BAT function. Serum T4 levels are positively correlated with UCP1, CIDEA, and PRDM16 in WAT [66]. The thyrotropin receptor (TR) agonist-mediated induction of browning may be a treatment-appropriate strategy for obesity, and treatment with the TR agonist GC-1 promotes WAT browning and reduces obesity and diabetes in mice [67]. Coordinated synergistic effects between glucagon and thyroid hormones may correct glucose tolerance and obesity [68].
4.1.3. Environmental and Lifestyle Effects
During exercise, skeletal muscle contraction releases a fibronectin-like peptide called irisin. This hormone, produced in both mice and humans, upregulates PPARα and UCP1 expression, thereby acquiring the ability to induce WAT browning [69], positively affect energy metabolism [70], and improve high-fat diet (HFD)-induced insulin resistance (IR) [71]. The brain-derived neurotrophic factor (BDNF), also released after exercise, induces WAT browning and increases energy expenditure [72]. Lactate production during anaerobic exercise may also lead to WAT browning [73].
The levels of circulating fibroblast growth factor 21 (FGF21) are elevated under cold conditions. FGF21 may promote WAT browning through FGFR1 and scaffold βKlotho, thereby promoting thermogenesis [74,75]. FGF15/19, FGF family members, are also associated with AT plasticity during thermogenic adaptation. FGF19 levels in humans directly correlate with UCP1 expression in the SAT, whereas increased FGF15 or FGF19 levels in mice induce WAT browning [76]. Cold-activated BAT leads to an enhanced uptake of triglyceride-rich lipoproteins through lipoprotein lipase and the transmembrane receptor CD36, thereby accelerating serum triglyceride removal [77]. Although burns also cause WAT browning, this browning is impaired in old patients and is associated with reduced survival in old burn patients [78].
Intermittent fasting selectively stimulates beige adipocyte development in WAT and reduces obesity by modulating the gut microbial composition in order to increase acetate and lactate biosynthesis [79]; acetate [80] and lactate [81] have been proven to be browning agents.
4.2. Whitening
When various external stimuli are removed, such as reheating adipocytes after cold stimulation, catecholamine stimulation is no longer present, and beige or brown adipocytes tend to lose their characteristics. Moreover, they exhibit an increase in white adipocyte characteristics. This process is known as “whitening.” The modifications include a change in internal multilocular lipid droplets to unilocular forms, reduction in mitochondria, and decreased UCP1 expression (Figure 1C). [82]. Although the identity of beige adipocytes is altered, their epigenomic memory is retained from previous cold exposure, and hence, they can activate thermogenic genes faster when the next cold stimulus is applied, which is associated with the action of poised enhancers. This particular process is not evident in brown adipocytes [83]. The glucocorticoid receptor (GR) and its downstream target Zfp423 have a crucial role in whitening. Inguinal subcutaneous WAT (iWAT) from both adipocyte-specific GR knockout mice and adipocyte-specific Zfp423 knockout mice have exhibited delayed whitening after rewarming [83].
5. Plasticity of the Internal Structure of Adipocytes
Internally, adipocytes include lipid droplets, mitochondria, and the nucleus. Under different physiological or pathological conditions, the size and number of lipid droplets as well as the number and activity of mitochondria in adipocytes change significantly to adapt to the energy demand at different stages. Lipid storage in adipose tissue involves the uptake of circulating fatty acids and glucose, which are converted into triglycerides and stored in lipid droplets within adipocytes. Conversely, lipolysis is the process by which these stored triglycerides are broken down into free fatty acids and glycerol, which are released into the bloodstream for energy use. These processes are tightly regulated by hormonal signals, with insulin promoting lipid storage and catecholamines stimulating lipolysis [84]. The expression level of UCP1 protein, a part of the mitochondrial inner membrane associated with heat production, also fluctuates under the aforementioned conditions. For the WAT-to-BAT or WAT-to-beige AT transition, a reduction in adipocyte size and an increase in mitochondrial content is necessary, and this moderate browning serves to protect WAT [85]. Specifically, WAT shows unilocular large lipid droplets, whereas BAT and beige AT exhibit multilocular small lipid droplets. Mitochondrial content and UCP1 levels gradually increase in the order of white to beige to brown adipocytes [86]. Plasticity has also been observed in mitochondrial activity, with atrial natriuretic peptide (ANP) and brain natriuretic peptide (BNP) released by cardiomyocytes during cold exposure. ANP enhances BAT mitochondrial activity in a p38 mitogen-activated protein kinase (p38 MAPK)-dependent manner [87], while BNP increases mitochondrial biosynthesis in WAT [88]. Multiple factors influence these changes. Rosiglitazone, a PPARγ agonist, enhances the expression of UCP1 and mitochondrial thermogenic genes in 3T3-L1 adipocytes [89]. THs and the sympathetic nervous system interact to increase UCP1 expression in BAT [90]. Autophagy is also involved in intracellular remodeling events occurring during brown/beige adipogenesis, and thermogenic activation and inactivation [91].
6. Plasticity of the External Matrix of Adipocytes
Along with adipocytes, AT contains many other cell types and structures, including adipose endothelial cells (AdECs) associated with tissue vascularization, nerve endings, non-differentiated precursor cells in various stages, and immune cells [16]. These cells and structures also have some degree of plasticity.
6.1. Plasticity of Blood Vessels in Adipose Tissue
Inflammation, fibrosis, and impaired angiogenesis are the triad of adipose tissue dysfunction. The relationship between these three conditions is complex, and the relationship between inflammation and angiogenesis in AT remains unclear [92]. AT function and growth are dependent on angiogenesis within the tissue. Blood flow supplies oxygen to the AT and transports nutrients, waste products, and various hormones and cytokines [93]. Various signaling molecules in the AT microenvironment are involved in the modulation of the adipose vascular system (Figure 2A). In healthy physiological conditions, such as exposure to cold, angiogenesis is positively regulated. This process is driven by the production of angiogenic factors, including vascular endothelial growth factor (VEGF), IL-4, and angiopoietin, which induce new blood vessel formation to meet the increased metabolic demand of the tissue [94]. Such changes are reversible, and recovery is possible after stress removal [39]. BMP4, as an angiogenic molecule, may also act as an essential regulator of this process [95]. On the other hand, in pathological conditions, the regulation of angiogenesis becomes more complex. but when this pathway is impaired—such as through PPARγ silencing—AdEC angiogenic activity decreases, leading to reduced FFA uptake and exacerbating inflammation [96]. In acute inflammation, angiogenesis can be positively regulated by inflammatory cytokines like IL-6, IL-8, and TNF-α, which promote the formation of new blood vessels to support tissue repair [97]. However, during chronic inflammation, often triggered by sustained AT expansion and hypoxia, angiogenesis becomes impaired, contributing to tissue dysfunction [92]. Interestingly, anti-inflammatory M2 macrophages are more pro-angiogenic compared with pro-inflammatory M1 macrophages [98]. ASCs in AT may be involved in angiogenesis by secreting miRNA-containing exosomes, such as miR126, miR31, miR92a, miR221, miR30, miR100, and miR486, which can directly regulate angiogenesis [94]. The latest 3D volume fluorescence-imaging procedure reveals that catecholamine signaling exerts crucial effects on AT vascular remodeling. The genetic deletion of the three βAR types would completely inhibit cold-induced vascular plasticity in WAT [99].

Figure 2.
The plasticity in the external matrix of adipocytes. (A) Plasticity of blood vessels in adipose tissue. Under cold conditions, adipose tissue angiogenesis is induced and regulated by factors such as vascular endothelial growth factor (VEGF), interleukin-4 (IL-4), angiopoietin, and bone morphogenetic protein 4 (BMP4). This process is reversible upon the removal of the cold stress. During acute inflammation, adipose tissue angiogenesis is promoted by factors such as interleukin-6 (IL-6), interleukin-8 (IL-8), and tumor necrosis factor-α (TNF-α). However, persistent chronic inflammation inhibits this process due to prolonged hypoxia. (B) Plasticity of nerves in adipose tissue. The cold exposure, the activation of adrenergic, and leptin signaling lead to an increased density of sympathetic axons in adipose tissue, facilitating enhanced thermogenesis. In contrast, this adaptive process is impeded in pathological conditions such as aging or obesity. (C) Plasticity of immune cells in adipose tissue. In obesity, there is an augmented conversion of adipose tissue macrophages into pro-inflammatory M1 macrophages, fostering the development of insulin resistance and inflammation. Conversely, in lean populations or under conditions such as cold exposure or burns, there is an increased transformation of adipose tissue into anti-inflammatory M2 macrophages, promoting angiogenesis, catecholamine production, and enhanced thermogenesis. The activation of MAPK signaling and NF-κB signaling pathways may be associated with M1 polarization, while BMP4, PPAR γ, PPAR δ, IRF-6, and adiponectin contribute to M2 polarization.
6.2. Plasticity of Nerves in Adipose Tissue
Sympathetic nerves in AT regulate the thermogenic capacity of adipocytes, and sympathetic nerve density varies with energy balance requirements [100]. Sympathetic nerves are the route of communication from the brain to AT that facilitates the regulation of adipocyte number and size as well as various functional changes [101] (Figure 2B). The plasticity of AT nerves depends on nerve growth factor (NGF)- and TrkA receptor-mediated cold-evoked signaling. When NGF or TrkA signaling is blocked, sympathetic nerve plasticity in AT is inhibited, thereby preventing the conversion of WAT to beige adipocytes under cold stimulation [102]. Eosinophils can also modulate sympathetic plasticity. Sympathetic activation leads to IL-33 release from stromal cells through adrenergic signaling, which then acts on innate lymphoid cells2 and induces IL-5 production. IL-5 is essential for adipose eosinophil regulation, and eosinophils subsequently regulate axonal growth within the adipose [100]. For the maintenance of sympathetic structural plasticity, leptin signaling is critical. Mice with leptin gene mutations (ob/ob mice) exhibit a significant reduction in sympathetic innervation in subcutaneous WAT and BAT, which is restored after chronic leptin treatment [103]. Leptin exerts its effects on innervation through the agouti-related peptide and pro-opiomelanocortin neurons in the arcuate nucleus of the hypothalamus, acting through brain-derived neurotropic factor-expressing neurons in the paraventricular nucleus of the hypothalamus (BDNFPVH) [103]. Cold-induced neuroimmune cells (CINCs) of bone marrow lineage origin that also secrete BDNF have a role in promoting neural remodeling in iWAT, where CINCs are increasingly recruited to AT and promote axonal survival during cold stimulation [101]. With advancements in techniques such as adipose tissue innervation imaging, sensory nerves have also been shown to play a role in regulating lipolysis, promoting angiogenesis, and in interacting with sympathetic nerves [104]. Under some pathological conditions, including aging and obesity, WAT fails to maintain proper innervation and undergoes “adipose neuropathy”, exhibiting reduced protrusions, loss of synaptic markers, and reduced local expression of neurotrophic factors [105].
6.3. Plasticity of Immune Cells in Adipose Tissue
The scattered distribution of numerous immune cells in AT is closely related to the development of inflammation and obesity-related metabolic disorders [106]. The plasticity of immune cells infiltrating AT is also significant, mainly in the composition of macrophage subsets. Most AT macrophages (ATMs) infiltrating obese fat are arranged around the remnants of dead adipocytes, forming distinctive “crown-like structures” [85]. In obesity, lipid-associated macrophages (LAMs) and proliferative LAMs are significantly increased and become the “dominant” macrophage subpopulation. The expression of cytokines such as chemokine ligand 8, osteopontin, and progranulin is dramatically upregulated in obese macrophages [107]. Macrophages tend to act as pro-inflammatory M1 macrophages in most populations with obesity, whereas they predominantly act as anti-inflammatory M2 macrophages in lean populations [108] (Figure 2C). The activation of MAPK signaling [109] and NF-κB signaling [110] pathways may be associated with ATM polarization to M1 macrophages. BMP4 activates M2 macrophages and inhibits M1 macrophage polarization, and activated M2 macrophages also promote AT angiogenesis [111]. PPARγ [112], PPARδ [113], interferon-regulatory factor-6 [114], and adiponectin [115] are also believed to contribute to M2 polarization. In the WAT of people affected by obesity, M1 macrophages are increased, resulting in pro-IR and pro-inflammatory conditions. Although the absolute number of M2 macrophages increases with obesity development, they continue to be less numerous than M1 macrophages in relative terms [116]. Interestingly, cold exposure or burns are associated with macrophage infiltration, with an increase in M2 macrophages in adipose tissue in both cases. Catecholamine production was promoted, promoting WAT browning and increasing systemic thermogenesis [117,118]. Exercise and cold exposure-induced circulating factor meteorin-like (Metrnl) also promote alternative macrophage activation in AT, correlating with the expression of anti-inflammatory genes in AT [119]. Unlike macrophages in other tissues, macrophages in AT also regulate lipid metabolism and recycle lipids from adipocytes as well as catecholamines from neurons [120].
In addition to that of macrophages, B-2 and T cell plasticity in AT has been reported. B-2 cells accumulate in the AT of diet-induced obese mice, and increased IgG2c antibodies derived from B-2 cells mediate IR [121]. Cytotoxic T cells (CD8+ T cells) are increased in obesity-induced inflammation at the early stages and are related to macrophage recruitment and activation [122]. CD4+ helper T cell 1 (Th1) promotes inflammation in obesity development and increases in number, while CD4+ helper T cell 2 (Th2) has an anti-inflammatory role and decreases in number [123]. VAT Foxp3 regulatory T cells (Tregs) protect AT from inflammation during obesity, a process in which PPAR-γ expression is crucial [124]. The beneficial effects of VAT Tregs and M2 macrophages can be maintained through the synergistic effect of invariant natural killer T cells, which are associated with IL-2 and IL-10 secretion [125].
7. Plasticity of Adipose Tissue Distribution
AT distribution in humans can be classified into SAT, VAT, and bone marrow fat. SAT primarily serves as an energy reservoir and is associated with lower health risks, while VAT is linked to metabolic disorders, including insulin resistance and cardiovascular disease [126]. AT distribution can also shift under some physiological or pathological conditions or the effect of hormones or drugs (Figure 3). For instance, in patients with Cushing’s syndrome, glucocorticoids cause a greater migration of systemic lipids to visceral fat and increased fat consumption in the extremities [127]. AT redistribution from the periphery of the body to the abdomen under physiological conditions seems to result from aging and the effect of sex hormones, thereby adapting to a lower basal metabolic rate and reduced physical activity [128]. Hormones impact the plasticity of AT distribution; for example, postmenopausal women often experience an increase in VAT due to declining estrogen levels, whereas estrogen supplementation may help reduce this effect [129]. Similarly, hyperandrogenemia in patients with polycystic ovary syndrome (PCOS) also stimulates greater fat accumulation toward visceral adiposity [130].

Figure 3.
Plasticity of adipose tissue distribution. Pathological conditions such as Cushing’s syndrome and polycystic ovary syndrome (PCOS), along with physiological changes like menopause and aging, contribute to the shifting of subcutaneous fat to visceral fat. With aging and obesity, there is also an observed ectopic deposition of fat in the liver and skeletal muscle. Notably, the role of adiponectin is paramount in promoting the redistribution of ectopically deposited fat from tissues, such as the liver, back to the subcutaneous fat depot.
Aging also causes changes in fat distribution, and most of the population aged >65 year tend to accumulate fat as the visceral fat depot, resulting in visceral obesity [131]. Adiponectin releases the “adipocyte starvation” signal to the body, facilitating the transfer of lipids from ectopic sites, such as the liver and muscle, to SAT, which helps lower blood lipid concentrations [132]. Additionally, the ethanolic extract of the Platycodon grandiflorus root also reduces blood lipid levels by increasing FFA uptake in epididymal WAT [133].
In a population affected by obesity, fat deposition also occurs in some non-adipose tissues such as the liver and skeletal muscle. This ectopic fat accumulation is often linked to the age-related dysregulation of lipid metabolism in SAT [134]. Lipotoxicity refers to the harmful effects of excess lipids on non-adipose tissues, which can disrupt cellular function and lead to metabolic disorders. However, the presence of adipokines, such as adiponectin, can mitigate lipotoxicity by facilitating the transfer of excess lipids from these ectopic sites back to adipocytes, thereby reducing the burden on non-adipose tissues and improving overall metabolic health in individuals with obesity [132].
8. Plasticity of Adipocyte Differentiation and Dedifferentiation Capacity
8.1. Plasticity of Adipocyte Differentiation Capacity
BAT originates in the paraxial mesoderm, while WAT originates in the mesoderm and neural crest. The different origins of BAT and WAT may influence their different plasticity. Adipocyte differentiation can be understood as adipocyte generation. The first step in AT generation is the spectrum commitment of MSCs to preadipocytes, followed by the growth arrest and differentiation of preadipocytes into lipid-containing functional mature adipocytes [135]. MSCs are precursors of adipocytes as well as common progenitors of myogenic, osteogenic, and chondrogenic cells [136]. Adipose-derived stem cells (ADSCs) exhibit great plasticity and are a potential driving force for stem cell therapy because of their relative accessibility and extensive self-renewal potential [137]. Increasing ADSC conversion to osteoblasts, transplanting autologous osteogenic pre-differentiated ADSCs into the fracture gap, and forming seed cells for bone tissue engineering may be one of the possible therapeutic avenues for refractory fractures [138]. PPARγ [139] and the coactivator CCAAT/enhancer binding protein α (C/EBPα) are the main regulators of adipogenesis [140]. However, none of these regulators are highly expressed in early adipogenesis. Instead, their transcriptional activator C/EBPβ shows transiently high expression, implying that it may act as a key regulator of early adipogenesis [141]. Interestingly, a study found that PPARγ phosphorylation correlates with sex differences in the differentiation of iWAT adipocyte precursor cells (APCs). Namely, in male HFD-fed mice, iWAT APCs were resistant to adipogenesis, whereas perigonadal WAT (gWAT) APCs were activated and pro-adipogenic. By contrast, in HFD-fed female mice, de novo adipogenesis was observed in both iWAT and gWAT [142]. Krüppel-like factors (Klfs), transcriptional regulators, are also highly expressed during early adipogenesis. Among them, Klf4, Klf5, and Klf15 are positively involved in adipogenic differentiation [143], and Klf4 can bind to the C/EBPβ promoter to promote PPARγ and Klf15 expression ([144], p. 4). By contrast, Klf2, Klf3, Klf7, and Klf9 are negatively associated with adipogenic differentiation [143]. TGF-β is also a member of adipogenesis suppressors. These suppressors can inhibit adipogenesis by activating SMAD proteins to promote the expression of ECM-related genes such as collagen genes [145]. Some BMP members in the TGF-β superfamily perform different functions. BMP2 and BMP4 can effectively promote adipocyte production through the BMP-SMAD-lysyl oxidase pathway [146], while BMP7 can promote brown adipocyte production by increasing PRDM16 and PGC-1α [45]. Matrix metalloproteinase 11 (also known as globular plasmolysin 3) in the AT microenvironment acts as a negative regulator of adipogenesis [147].
8.2. Plasticity of Adipocyte Dedifferentiation Capacity
Dedifferentiation, a mechanism of adipocyte plasticity, is the conversion of mature adipocytes back to an undifferentiated progenitor cell-like state [13]. Mature adipocytes successfully cultured using the “ceiling culture” method can, under certain conditions, lose their lipid droplets and revert to progenitor-like cells, called dedifferentiated adipocytes. Dedifferentiated adipocytes have a multispectral differentiation potential both in vivo and in vitro and expresses various ASC markers, suggesting stem cell properties [148]. Under physiological conditions, such as during lactation, adipocytes may lose lipid droplets and dedifferentiate into fibroblast-like cells [13], whereas, during wound healing, myofibroblasts regenerate and differentiate into adipocytes, implying that dedifferentiation is reversible [149]. The dedifferentiation ability of adipocytes is also reflected in tumor progression. In breast cancer, adipocytes present around cancer cells transform into fibroblast-like cells and express high levels of cancer cell markers, thereby driving breast cancer progression [150]. Aberrant Notch signaling activation drives adipocyte dedifferentiation and the transformation of these adipocytes into liposarcoma cells [151]. When 3T3-L1 adipocytes and the pancreatic cancer cells MiaPaCa2 are co-cultured in vitro, MiaPaCa2 cells release Wnt5a, which target Wnt signaling in 3T3-L1 adipocytes. Treatment with secreted trichome-associated protein 5, a Wnt5a inhibitor, blocks downstream Wnt signaling in 3T3-L1 cells and prevents their dedifferentiation [152]. Bleomycin promotes the transformation of adipocytes to myofibroblasts in the SAT [153] and can inhibit adipogenesis by enhancing TGF-β1 expression ([154], p. 1). Adipocyte dedifferentiation is potentially valuable for applications, as inhibiting peritumor adipocyte dedifferentiation or promoting fibroblast-like cell redifferentiation in breast cancer may inhibit breast cancer progression. The reversibility of adipocyte dedifferentiation also suggests a broad range of applications for DFAT cells, which may be used in wound healing.
9. Plasticity of Adipocytes Secreting Adipokines or Extracellular Vesicles
Adipocytes possess important endocrine functions and can secrete various signaling molecules known as adipokines (Figure 4). Adipokines play crucial roles in regulating metabolism, inflammation, and insulin sensitivity, as well as influencing processes such as appetite control and cardiovascular health. Leptin, the first adipokine to be demonstrated, is a critical feedback signal from the brain regarding AT size and status, and it regulates energy expenditure [155]. A deficiency of leptin and its receptors causes severe obesity in relevant mouse models and humans [156]. Leptin primarily transmits nutritional feeding signals to the hypothalamus, resisting obesity onset by reducing appetite and promoting energy expenditure [157]. Leptin also plays a regulatory role, reducing adipogenesis as well as promoting lipolysis, in relation to sympathetic innervation around AT [158]. The expression of the pro-inflammatory adipokines TNF-α and IL-6 is significantly reduced during cold exposure [159]. In obesity, pro-inflammatory pathways within adipocytes, including JNK and NF-κB pathways, are activated and adipokine secretion is altered accordingly. Levels of pro-inflammatory cytokines such as TNF-α [160] and MCP-1 [16] are elevated, whereas those of anti-inflammatory cytokines such as adiponectin are declined [161]. Resistin [162] and retinol-binding protein 4 [163] are also well-studied adipokines.

Figure 4.
Plasticity of adipocytes secreting adipokines or extracellular vesicles. Adipocytes secrete a variety of adipokines, including adiponectin, leptin, IL-6, and TNF-α, which regulate adipogenesis and lipolysis (lower left panel). The expression of pro-inflammatory cytokines such as IL-6 and TNF-α is significantly reduced under cold conditions. In obesity, the expression of leptin and pro-inflammatory cytokines increases, while the expression of anti-inflammatory cytokines, such as adiponectin, decreases. Leptin inhibits the onset of obesity by establishing a link with the hypothalamus to reduce appetite and promote energy expenditure. Adiponectin releases the adipocyte starvation signal so as to promote the redistribution of fat deposited ectopically in the liver/muscle to the subcutaneous fat depots. In addition, adiponectin regulates the vascular tone and endothelial functions, including coronary arteries, which affect the heart. Adipocytes release extracellular vesicles, transporting proteins, mRNA, and non-coding RNA (e.g., miRNA) to target the organs and cells (upper right panel). In a population affected by obesity, the circulating levels of EVs are significantly elevated, thereby regulating a range of processes, such as inflammation, fibrosis, and tumor progression.
AT can regulate systemic homeostasis by releasing extracellular vesicles (EVs) that act on other tissues to regulate lipid and glucose homeostasis (Figure 4). EVs are crucial mediators of cellular communication between AT and other tissues. They deliver proteins, mRNAs, and non-coding RNAs to recipient cells, thereby altering their phenotype [164]. Leptin and adiponectin are also expressed in EVs secreted by 3T3-L1 adipocytes and the human SAT [165]. Obesity significantly increases the circulating levels of EVs enriched with perilipin A, an adipose-specific protein useful for detecting circulating adipocyte-derived EVs [165]. miRNAs contained within secreted EVs are altered in obese AT compared with lean AT. miR-148b and miR-4269 are downregulated, whereas miR-23b and miR-4429 are upregulated [166]. The mRNAs predicted by differentially expressed miRNAs are also expected to change with obesity development through the TGF-β and Wnt/β-catenin signaling pathways, thereby regulating inflammation and fibrosis in other organs [166]. Some EVs are taken up by immune cells and can activate pro-inflammatory macrophages that drive local and systemic inflammation and IR [167]. siRNA- and miRNA-containing paracrine EVs secreted by ASCs in the adipose tissue microenvironment of patients with ovarian cancer contribute to tumor progression [168]. The presence of various adipokines and EVs highlights that AT is indispensable in the regulation of the systemic endocrine system.
10. Thresholds of Adipose Tissue Plasticity and Their Individualized Differences
AT plasticity in all aspects is not never-ending. Beyond a certain threshold, irreversible damage may occur. In the group with obesity, for example, when AT plasticity is within the threshold, the adipocytes become moderately larger and more numerous, inflammation occurs mildly, angiogenesis increases, and thus hypoxia occurs rarely. This may explain why some infants and adolescents with obesity and people with metabolic health obesity (MHO) do not develop metabolic disorders [169,170,171]. By contrast, when AT plasticity exceeds the threshold, in terms of adipocyte size regulation, FFA leaks into the cytoplasm as lipid droplets reach their maximum storage limit. This increases oxidation and endoplasmic reticulum stress in adipocytes, which can result in cellular dysfunction, inflammation, and ultimately contribute to metabolic disorders [172]. SAT is the largest and best storage site for excess lipids. However, its limited ability to recruit precursor cells reduces the production of new adipocytes, forcing reliance on hypertrophy for lipid storage, which ultimately leads to unhealthy AT expansion accompanied by increased inflammation and IR [95,173]. Regarding adipose ECM remodeling, excessive infiltrating macrophages impair mature adipocyte generation, thereby disrupting ECM homeostasis, resulting in fibrotic deposition, and exacerbating inflammation [174]. Limited angiogenesis and subsequent hypoxia are also characteristics of pathological adipogenesis [175]. When the threshold of AT plasticity is breached, hypoxia, fibrosis, inflammation, and other adverse effects occur back-to-back and interact to disrupt body homeostasis.
AT plasticity thresholds vary for individuals. Significant differences exist between individuals in response to excessive energy intake, which may explain the presence of MHO and metabolically unhealthy obesity (MUO) in clinical practice [176] (Figure 5). The physiological basis for MHO existence may be the high threshold of AT plasticity in a population subset. Despite their potential for metabolic abnormalities, because of high threshold of AT plasticity in MHO, the body has a higher capacity for adaptive changes and thus does not show metabolic abnormalities. Genetic variants are an essential influence. Genome-wide association studies have identified a set of loci-harboring genes that possibly control excess body fat distribution and metabolism, with 14 genetic variants associated with a lower metabolic abnormality risk. A study associated with UK Biobank revealed that 11 of these variants were related to a higher body mass index (BMI) and adiposity. This implies that some individuals with obesity are possibly at a lower risk of metabolic abnormalities, partially explaining the reason for the presence of the MHO phenotype [177]. Age differences also contribute to individual differences in AT plasticity. The BAT thermogenic activity progressively decreases with aging, with reduced metabolic capacity and greater susceptibility to chronic diseases [131]. The ASC differentiation and proliferation potential decrease considerably with age, and so does the ability of preadipocytes to differentiate and store lipids, allowing more lipotoxic FFAs to harm other organs [178].
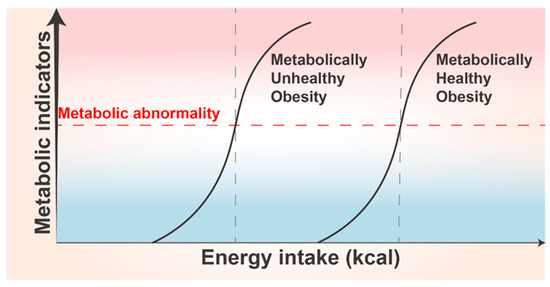
Figure 5.
Individual differences in adipose tissue plasticity. Metabolically unhealthy obese (MUO) populations demonstrate a lower threshold for adipose tissue plasticity compared to metabolically healthy obese (MHO) populations, where the latter exhibit a higher threshold. In cases of equal energy intake, MUO individuals are more susceptible to metabolic disorders, characterized by abnormal increases in various metabolite types and densities. These aberrations subsequently lead to disturbances in a range of metabolic indicators.
11. Relationship between Adipose Tissue Plasticity and Disease
AT plasticity is an active adaptive capacity of AT to self-regulate in response to various bodily changes and to protect the body to a certain degree. As mentioned above, this protective capacity is limited, and beyond the limit, it can contribute to the development of various diseases and turn into an injury.
11.1. Obesity
A hallmark of obesity is unhealthy AT expansion, including excessive hypertrophy and hyperplasia of adipocytes, often beyond the plasticity threshold in adipocyte size and number. Dysfunctional AT also exhibits varying degrees of inflammation, impaired energy metabolism, and cellular senescence, while inducing several systemic complications by promoting ectopic lipid overflow and the release of different adipokines [1]. Persistent inflammation in AT contributes significantly to obesity development. Compared with WAT, BAT from HFD-fed mice exhibits lower immune cell enrichment, inflammatory marker expression, and macrophage infiltration, suggesting that BAT can resist obesity-induced inflammation [179]. Despite some anti-inflammatory mechanisms of BAT, a sustained and intense local inflammatory environment and some pro-inflammatory cytokines directly impair the thermogenic and metabolic activities of BAT and beige AT. This severely affects energy expenditure mechanisms and leads to obesity-related metabolic syndrome and a series of cardiovascular alterations [16,180]. BAT activation and the induction of WAT browning accelerate glucose and lipid uptake and reduce insulin secretion requirements. This may be a novel strategy for improving glucose and lipid metabolism and IR in people with obesity and those with T2DM [86]. The inflammatory environment of AT also disrupts ECM homeostasis, leading to fibrotic deposition. Fibrosis further limits AT healthy expansion in terms of both physical restriction and functional impairment, exacerbating obesity progression [181,182]. The plasticity of adipogenesis is also impaired in obesity development. The preadipocyte proportion in the SAT is reduced in obesity, with a corresponding reduction in the generation of new healthy adipocytes promoting AT dysfunction [183]. Pro-inflammatory macrophages also disrupt de novo adipogenesis in WAT by secreting pro-inflammatory cytokines that inhibit preadipocyte differentiation and impair normal signaling pathways, such as the insulin and PPARγ pathways, leading to increased lipotoxicity and metabolic dysfunction [174].
11.2. Lipodystrophy
Lipodystrophy is a rare disease characterized by selective AT loss. It often manifests as a decrease in SAT of the trunk and extremities and an increase in VAT and fat of the head and neck [184]. Very little amounts of AT can have the same adverse effect on the body’s metabolism as very large amounts of AT. Similar to that in people with obesity, excess lipids are transferred to the liver, bones, and other tissues for ectopic deposition in patients with lipodystrophy and are associated with IR development [185]. Such IR can be improved by the leptin’s action [186]. In a patient with autoimmune lipodystrophy, P. Fischer-Posovszky et al. found that AT loss was associated with cellular inflammation in lymphoid tissue and increased TNF-α concentrations. TNF-α and interferon-γ stimulated CD95 expression and enhanced the formation of CD95 death-inducing signaling complexes in adipocytes, suggesting that CD95-induced apoptosis mediates adipocyte loss [187]. The endoribonuclease Dicer regulates the plasticity of AT species. In HIV-infected patients with lipodystrophy, reduced Dicer levels lead to increased BAT whitening and the downregulation of BAT and beige AT-related gene expression, which triggers metabolic dysregulation [188].
11.3. Insulin Resistance
IR is a pathological state often correlated with obesity and the development of various obesity-related complications. IR development is also inextricably linked to AT plasticity. Hypertrophic adipocytes activate the JNK and NF-κB pathways, initiating the transcription of pro-inflammatory genes and the secretion of adipokines such as TNF-α and IL-6. This process triggers localized and systemic chronic inflammation, which impairs insulin signaling pathways, leading to insulin resistance. Furthermore, macrophage infiltration is promoted, exacerbating the development of T2DM [189]. Lipodystrophy and obesity, as previously described, lead to excessive lipid deposition in other organs such as the liver and muscle, which are hallmarks of and major contributors to IR [190]. Lactate links IR and obesity. Slc16a1, which encodes the major lactate transporter protein, when deleted in adipocytes, can lead to intracellular lactate accumulation. This induces apoptosis and the release of large amounts of pro-inflammatory adipokines, thereby causing persistent inflammation and systemic IR [191]. EVs are critical tools in IR progression, and in addition to the previously described EV functions, EVs may contribute to the exacerbation of cognitive impairment and synaptic damage by IR [192]. Macrophage-produced peptidase D promotes AT fibrosis, while excessive AT fibrosis exacerbates IR progression [193]. New-onset adipogenesis promotes healthy AT expansion and can prevent obesity-associated IR ([194], p. 2). While high lipolysis rates are also associated with IR, serotonin receptor 2B (HTR2B)/5-hydroxytryptamine signaling in AT may be involved in lipolysis regulation. HTR2B expression levels are elevated within VAT in people with obesity, and selective HTR2B antagonists may attenuate the inflammatory response in VAT and obesity-associated IR [195]. L. Wang et al. identified a new cell line, p21Cip1 (Cyclin-dependent kinase Interacting Protein 1)-highly expressing (p21high) cells, through single-cell transcriptomics. These cells in AT cause IR in vivo, which is possibly associated with the NF-κB pathway. Intermittent clearance of p21high cells can alleviate IR [196].
11.4. Polycystic Ovary Syndrome
PCOS is a common endocrine disorder observed in premenopausal women. It is characterized by hyperandrogenism and ovarian dysfunction-related symptoms. PCOS is affected by various epigenetic and environmental factors [197]. High androgen levels can promote the differentiation from preadipocytes to mature adipocytes in the abdomen, and thus aggravate the visceral obesity phenotype [198]. AT regulates PCOS courses mainly through adipokine release. Elevated levels of AT-secreted leptin in women with obesity suppress insulin-induced ovarian steroid production and inhibit the pro-estradiol production of the luteinizing hormone (LH) [199]. Serum adiponectin levels increase with weight loss and have a regulatory influence on the central reproductive endocrine axis, such as inhibiting the release of LH and the gonadotropin-releasing hormone [200]. Decreased levels of endometrial adiponectin receptors AdipoR1 and AdipoR2 are correlated with implantation failure during pregnancy in women with PCOS [201]. In a randomized controlled study, serum resistin levels were significantly lower in people affected by obesity and PCOS who were treated with the insulin sensitizer rosiglitazone, implicating its potential utility in PCOS treatment [202]. The levels of pro-inflammatory adipokines such as MCP-1 were also significantly elevated in patients with PCOS, whereas TNF-α and IL-6 levels were not altered [203]. Ovarian dysfunction and hyperinsulinemia in patients with PCOS are strongly associated with the persistent and chronic inflammation of AT, and both obesity and IR contribute to PCOS progression by exacerbating inflammation [204].
11.5. HAIR-AN Syndrome
HAIR-AN syndrome is characterized by hyperandrogenemia (HA), IR, and acanthosis nigricans (ANs) and is mainly caused by AT dysfunction [205]. A cross-sectional study found that patients with HAIR-AN syndrome had higher leptin levels and lower adiponectin levels than those with PCOS, which implies a more significant role of adipokines in HAIR-AN syndrome [206]. IR and HA have been described previously, where AN was skin pigmentation and hyperkeratosis exhibited in the axillae, posterior neck, etc. [207]. This cutaneous manifestation may predict hyperinsulinemia in people with obesity but without diabetes [208]. Symptoms of AN have also been reported in patients with acquired lipodystrophy and hereditary IR syndrome [209,210].
11.6. Atherosclerosis
A key contributor to atherosclerosis is obesity-induced adipose dysfunction, and ectopic fat deposits, such as in the liver, and epicardial fat deposits also increase the atherosclerosis risk [211]. Adipocyte hypoxia-inducible factor 2α may exhibit anti-atherosclerotic effects by reducing hepatic cholesterol levels [212]. The AT distributed around blood vessels, called perivascular AT (PVAT), is a critical factor affecting cardiovascular system homeostasis, and a bidirectional regulatory pathway exists between the vessel wall and PVAT [213]. PVAT is as malleable as AT in other locations. Physiologically, PVAT provides mechanical support and temperature maintenance for the vasculature. Moreover, it regulates vascular tone and endothelial function by secreting adipokines such as adiponectin [214]. Adiponectin synthesis is regulated by circulating BNPs [214]. However, during vascular injury, PVAT undergoes inflammatory changes and long-term inflammation can lead to pathological vascular remodeling [215]. BMP4 may inhibit atherosclerosis progression by promoting the browning of PVAT, which increases mitochondrial activity and thermogenesis, and by exerting anti-inflammatory effects through the reduction in pro-inflammatory cytokines and macrophage infiltration; browning is considered a protective mechanism of PVAT [216].
11.7. Metabolic-Associated Fatty Liver Disease
MAFLD, also previously known as non-alcoholic fatty liver disease, is closely related to metabolic diseases such as obesity. It increases the danger of liver failure and hepatocellular carcinoma [217]. In fatty liver disease, the lipid content of the liver increases and adipocytes are ectopically deposited in the liver, which is the outcome of a change in the plasticity of AT distribution. Macrophages have a critical role in MAFLD development, and dysregulated AT metabolism can activate hepatic macrophages in MAFLD [218]. Pro-inflammatory macrophages are correlated with MAFLD severity [219]. Adiponectin promotes macrophage polarization to an anti-inflammatory phenotype. Experiments have demonstrated that adiponectin KO mice are more susceptible to steatohepatitis, whereas adiponectin administration delays steatohepatitis progression [220]. Leptin levels were also elevated in patients with MAFLD and may mediate liver fibrosis by affecting Kupffer cells [221]. AT inflammation may occur earlier than liver tissue inflammation in MAFLD pathogenesis [222]. PPAR-γ agonists exert an ameliorative effect on hepatic steatosis in patients with MAFLD by inducing the anti-inflammatory phenotype of macrophages [223].
11.8. Tumor
AT plays a crucial role in the development and progression of various tumors. Adipocytes facilitate tumor metastasis, provide energy for rapid tumor growth, and produce signaling molecules that stimulate cell invasion and migration [224]. In ovarian cancer, human omental adipocytes promote cancer cell homing, migration and invasion by secreting adipokines such as IL-8 [225]. In breast cancer, FFAs generated from adipocyte lipolysis are transferred to cancer cells, fueling their fatty acid metabolism and enhancing tumor growth [226]. Similarly, in melanoma, adipocyte-derived lipids contribute to tumor progression through fatty acid transporter protein [227]. Moreover, obesity-associated alterations in inflammation and adipokine signaling, including elevated leptin levels and reduced adiponectin, further promote tumor growth and metastasis [228].
12. Mechanisms and Drugs Targeting Adipose Tissue Plasticity in Obesity Treatment
During obesity, to accommodate excess energy, AT undergoes diverse changes, including adipocyte hypertrophy, hyperplasia, and remodeling of the vascular system and ECM. This allows sufficient AT expansion to coordinate the mobilization of various nutrients [229]. However, after a certain limit is exceeded, several hazards such as persistent inflammation, fibrosis, and hypoxia ensue. The adverse effects of obesity originate in AT pathological remodeling [175]. Many treatments have been reported for various aspects of adipose plasticity (Figure 6).

Figure 6.
Mechanisms and drugs targeting adipose tissue plasticity in obesity treatment. Mechanisms including the regulation of adipocyte production and AT expansion increase in the BAT mass and activity, regulation of adipocyte apoptosis, improvement in adipose tissue angiogenesis, and improvement in inflammation and fibrosis in the adipose tissues. The drugs associated with each of the therapeutic mechanisms are listed. AT: adipose tissue. BAT: brown adipose tissue. TZD: thiazolidinediones. DAT: decellularized adipose tissues. GKA: glucokinase agonists.
12.1. Regulation of Adipocyte Production and Adipose Tissue Expansion
The failure of adipose precursor cells to form new adipocytes is among the major causes of AT dysfunction and is also closely related to obesity development [1]. PPARγ is a primary regulator of adipogenesis, and the PPARγ agonist TZD drives sirtuin 1-dependent PPARγ deacetylation, facilitating the interaction of PPARγ with PRDM16 and inducing metabolic and cellular remodeling. This thus improves AT’s ability to sequester fatty acids [230,231,232]. Decellularized adipose tissue (DAT) does not contain cellular structures but is rich in collagen, which thus creates a good microenvironment for adipogenesis. DAT is considered an ideal derived scaffold for AT by interacting with different types of MSCs for eventual adipose regeneration [233]. Enabling massive SAT expansion has been shown to counteract excessive caloric intake-associated IR and to improve lipid metabolism [132].
12.2. Increase in BAT Mass and Activity
BAT atrophy and increased visceral fat are observed in people with obesity; therefore, increasing BAT mass or activity is considered a potential strategy for obesity treatment [234]. BAT transplantation significantly prevents weight gain, reduces total fat weight, and increases oxygen consumption in mice with obesity, hence improving IR in them [235], which may be partially mediated by IL-6 [236]. Berberine [237], the water extract of Caulis Spatholobi [238], and ginseng extract [239] can improve obesity through BAT activation. The latter two can induce an increase in anti-obesity-associated bacteria by modulating gut microbiota composition. Although β3-AR agonists such as mirabellum can activate BAT, they have cardiovascular side effects and their use should be considered with caution [240]. Activated BAT can prevent obesity and T2DM by absorbing and utilizing glucose and lipids, improving IR, and reducing blood glucose, lipids, and total fat mass [241].
The molecules and regulatory pathways associated with WAT browning are new targets for obesity treatment [242]. Resveratrol, a natural antiaging polyphenol, increases the activation of the SIRT1 signaling pathway in AT to promote WAT browning [243] and is correlated with adipocyte apoptosis [244]. The GLP-1 receptor agonist liraglutide increases the expression of BAT marker genes in the SAT of diabetic rats, promotes SAT browning, and reduces VAT weight [245]. Glucokinase, an emerging antidiabetic target, and glucokinase agonists (GKA), such as the newly marketed dorzagliatin, can be used in routine diabetes treatment. GKA is safe, promotes insulin and GLP-1 secretion, and has potential weight loss effects [246]. The central browning agent nicotine may act through the κ opioid receptor in the lateral hypothalamus [247]. Retinoic acid [248], menthol [249], allicin [250], and pentamethylquercetin [251] have been found to promote WAT browning while exhibiting efficacy in reducing body weight and improving insulin sensitivity. However, differences in the browning capacity of individual fat depots may limit the therapeutic value of browning agents; moreover, VAT is less susceptible to browning agents than SAT [240].
Brown-like adipocytes, designed by CRISPR-Cas9 technology, and tested in mice for obesity and metabolic syndrome treatment have exhibited good efficacy and may offer therapeutic opportunities at the cellular level. Specifically, human brown-like (HUMBLE) cells were created by modifying human white preadipocytes with CRISPR/Cas9-SAM-gRNA to activate UCP-1 expression. The transplantation of HUMBLE cells into obese mice resulted in sustained improvements in glucose tolerance, insulin sensitivity, and increased energy expenditure [252].
12.3. Regulation of Adipocyte Apoptosis
Reducing adipocyte number by regulating adipocyte apoptosis can be complementary to other therapeutic options for obesity [253]. Green tea activates apoptotic signaling in adipocytes, with its active component possibly being (−)-epigallocatechin-3-gallate (EGCG). A reduction in the mass of individual fat depots was observed in EGCG-fed mice [254]. Lipodystrophy has been described in the treatment of patients with AIDS using the anti-HIV protease inhibitor ritonavir. This may be related to the apoptosis-inducing effect of ritonavir [255].
12.4. Improvement of Adipose Tissue Angiogenesis
Promoting angiogenesis can effectively improve local hypoxia, fibrosis, and inflammation caused by rapid AT expansion, but excessive angiogenesis may lead to unhealthy WAT expansion [256]. Therefore, whether angiogenesis inhibitors or promoters should be chosen to treat obesity remains controversial. Exenatide, a GLP-1 receptor agonist, increases AT angiogenesis through VEGF A in people with obesity and diabetes, improves AT chronic hypoxia, and alleviates obesity and obesity-related metabolic disorders [257]. Nifedipine, an antihypertensive drug, can improve obesity-impaired angiogenesis by inhibiting oxidative stress and increasing the number of endothelial progenitor cells. This thus improves AT hypoxia, increases mitochondrial thermogenesis, and halts obesity development [258]. Metformin has been found to have a role in obesity treatment. However, the exact mechanism remains unclear, and the inhibition of AT angiogenic activity is among the possible mechanisms [259].
12.5. Improvement of Inflammation in Adipose Tissue
Obesity promotes an inflammatory environment within the AT and exacerbates fibrosis. Furthermore, obesity alters ASC composition and function. Applying good lifestyle modifications, such as exercise and caloric restriction, may suppress or even reverse this class of pathological changes [260]. The regulation of different inflammation-associated pathways in macrophage polarization, thereby inhibiting M1 polarization and reducing inflammatory factor release, may be a potential strategy for obesity treatment [108]. By inhibiting M1 macrophage activation in AT, berberine may improve IR induced by chronic inflammation in AT, which in turn reduces serum TNF-α and IL-6 levels [261]. The TNF-α antagonist etanercept used for the treatment of obesity and related metabolic complications may alleviate obesity-induced inflammation and reduce fasting glucose levels by modulating adiponectin levels. Nevertheless, the wide application of etanercept is limited because of its possible side effects such as increased susceptibility to infection [262].
13. Discussion
Obesity is a highly prevalent and harmful condition and is associated with a wide range of complications. Obesity is ultimately caused by abnormal fat metabolism, and altered AT plasticity is a sign of abnormal fat metabolism. We here first provide the complete definition of AT plasticity. At the same time, we emphasize that a threshold of AT plasticity exists, which may be related to the presence of MHO and MUO. This AT plasticity allows the body to adapt rapidly to various alterations in energy requirements, and the underlying process is regulated by multiple factors [263].
AT expansion manifests itself as adipocyte enlargement or an increase in adipocyte number, which begins in fetal life and continues throughout the life cycle [264]. Changes in AT species are an essential part of AT plasticity, and far more studies have investigated the AT browning process than the whitening process. In addition to traditional fat depots, a natural class of beige adipose depots, called thigh adipose tissue (tAT), was recently identified and characterized in mice maintaining a beige adipose morphology at room temperature. tAT exhibits significantly higher BAT marker expression and a considerably greater browning potential than iWAT, and its UCP1 content is 2600-fold higher than that of iWAT under HFD stimuli, which may be a more suitable model for studying AT plasticity [265].
A unique type of AT known as “pink adipocytes” received its name due to its pink color during pregnancy and lactation. These cells are alveolar in nature and contain large cytoplasmic lipid droplets. In appearance, they closely resemble adipocytes and can transdifferentiate from WAT. A new concept, termed “pinking,” refers to the process of white-to-pink transdifferentiation or adipoepithelial conversion [266]. Perilipin1 is expressed in white adipocytes but not in pink adipocytes, while perilipin2 is characteristic of pink adipocytes and is absent in white adipocytes [267]. Lineage-tracing experiments have also confirmed the transdifferentiation of WAT into pink adipocytes [266]. During lactation, lipids are transferred from adipocytes to alveolar cells, which secrete fat as a component of milk. After lactation, the cytoplasm of lean adipocytes is refilled with lipids, allowing them to regain the typical WAT morphology [85]. Cells with intermediate features of pink and brown adipocytes have been identified in the dorsal mammary glands of post-lactation mice, where BAT in the interscapular region is located near the alveoli. These cells display several ultrastructural markers of brown adipocytes, such as multilocular lipid droplets and elevated UCP1 expression, indicating the transition from pink to brown adipocytes [268,269].
MSCs from the mesoderm are now widely considered a source of APCs [270,271]. Adipose single-cell mapping has confirmed the complex APC heterogeneity in mouse and human adipose depots [2]. APC profiling has revealed that most brown adipocytes originate from myogenic Myf5+ MSCs, whereas white adipocytes from adipogenic Myf5− MSCs. Indeed, both brown and white adipocytes originate from multiple lineages, which is possibly related to their plasticity [272]. The regulatory factors involved in preadipocyte differentiation and adipocyte dedifferentiation have been mentioned previously. Obesity induces both structural and functional changes in mitochondria within human MSCs, including decreases in mitochondrial matrix density and fatty acid metabolite levels [273]. Adipogenic regulatory cells (Aregs), which are a type of preadipocyte, were recently identified in mice and humans. Aregs are characterized by elevated Cd142 and Abcg1 expression and exert antiadipogenic effects on committed progenitor cells through a paracrine mechanism [274].
The threshold of AT plasticity simply means that the degree of AT changeability is limited by many aspects. The plasticity of AT distribution may be of wide interest due to its lipid-lowering effects. Collagen and proteoglycans are crucial ECM components in AT. They are present at an appropriate density in the healthy state, while their excessive accumulation leads to severe fibrosis, thereby promoting AT dysfunction [275]. AT over-expansion and the inability of angiogenesis to meet its demands lead to hypoxia and senescent cell accumulation, aggravating AT fibrosis and inflammation, and uncontrolled lipid spill over, producing lipotoxicity [92]. The cause–effect relationship is complex, and identifying the initiating factor and the outcome is difficult. The relationship between the presence of MUO and MHO and AT plasticity needs to be further explored clinically.
14. Prospect
AT has been a focus area of energy metabolism research for a long time, and recent advancements, such as scRNA-seq and spatial transcriptomics, have deepened our understanding of complex AT dynamics [260,276]. The WAT-on-Chip system has created the first human-based, autologous, and immunocompetent AT model in vitro. This system uses organ-on-chip technology to almost reproduce WAT heterogeneity and is expected to be a powerful tool in metabolism and related fields [277]. AT plasticity research will become a hot spot in the future, and more drugs related to AT plasticity regulation will be invented and developed. Therapies related to ADSC regulation and differentiation will receive attention because of their abundant source and high self-renewal potential. AT plasticity, as an active changeability, is irreplaceable in disease development. The multidimensional dissection of AT plasticity facilitates an in-depth understanding of adaptive changes in AT in response to systemic physiological or pathological changes and provides multiple potential targets for obesity treatment strategies to be applied in clinical practice.
15. Conclusions
In conclusion, AT plasticity is a complex and adaptive process that plays a central role in growth, obesity, organismal protection, and homeostasis. This plasticity involves dynamic changes in adipocyte size, number, lipid storage, and cellular composition, all of which are crucial for the tissue’s response to physiological and pathological stresses. Understanding the mechanisms behind AT remodeling offers valuable insights into its contributions to various diseases. By exploring AT plasticity, we can identify new potential therapeutic targets for managing obesity and other metabolic disorders, ultimately advancing relevant clinical applications.
Author Contributions
Conceptualization, Y.-Y.M. and H.-D.Z.; formal analysis, Y.-Y.M., Y.-X.H., S.-N.X., H.-L.J., H.-X.W., J.-M.C., F.X., H.-D.L., Y.W., Y.-Z.L., Y.-L.Y. and H.-D.Z.; investigation, Y.-Y.M., Y.-X.H., S.-N.X., H.-L.J., H.-X.W., J.-M.C., F.X., H.-D.L., Y.W., Y.-Z.L., Y.-L.Y. and H.-D.Z.; resources, L.L., Y.-H.B. and H.-D.Z.; writing—original draft preparation, Y.-Y.M.; writing—review and editing, Y.-Y.M., J.-M.C., Y.-L.Y. and H.-D.Z.; visualization, Y.-Y.M. and Y.-Z.L.; supervision, L.L., Y.-H.B. and H.-D.Z.; project administration, H.-D.Z.; funding acquisition, H.-D.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Natural Scientific Foundation of China [grant numbers: 82170900, 81970762], Hunan Administration of Traditional Chinese Medicine and Hunan Province high-level health Talents “225” project, and Graduate Student Innovation Program of Central South University (2024ZZTS0553).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- De Fano, M.; Bartolini, D.; Tortoioli, C.; Vermigli, C.; Malara, M.; Galli, F.; Murdolo, G. Adipose Tissue Plasticity in Response to Pathophysiological Cues: A Connecting Link between Obesity and Its Associated Comorbidities. Int. J. Mol. Sci. 2022, 23, 5511. [Google Scholar] [CrossRef] [PubMed]
- Emont, M.P.; Jacobs, C.; Essene, A.L.; Pant, D.; Tenen, D.; Colleluori, G.; Di Vincenzo, A.; Jørgensen, A.M.; Dashti, H.; Stefek, A.; et al. A single-cell atlas of human and mouse white adipose tissue. Nature 2022, 603, 926–933. [Google Scholar] [CrossRef] [PubMed]
- Cypess, A.M. Reassessing Human Adipose Tissue. N. Engl. J. Med. 2022, 386, 768–779. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.F.; Roesler, A.; Kazak, L. Regulation of adipocyte thermogenesis: Mechanisms controlling obesity. FEBS J. 2020, 287, 3370–3385. [Google Scholar] [CrossRef] [PubMed]
- Loyd, C.; Obici, S. Brown fat fuel use and regulation of energy homeostasis. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 368–372. [Google Scholar] [CrossRef]
- Schreiber, R.; Diwoky, C.; Schoiswohl, G.; Feiler, U.; Wongsiriroj, N.; Abdellatif, M.; Kolb, D.; Hoeks, J.; Kershaw, E.E.; Sedej, S.; et al. Cold-Induced Thermogenesis Depends on ATGL-Mediated Lipolysis in Cardiac Muscle, but Not Brown Adipose Tissue. Cell Metab. 2017, 26, 753–763.e7. [Google Scholar] [CrossRef]
- Carrière, A.; Lagarde, D.; Jeanson, Y.; Portais, J.-C.; Galinier, A.; Ader, I.; Casteilla, L. The emerging roles of lactate as a redox substrate and signaling molecule in adipose tissues. J. Physiol. Biochem. 2020, 76, 241–250. [Google Scholar] [CrossRef]
- Kopelman, P.G. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar] [CrossRef]
- GBD 2015 Obesity Collaborators; Afshin, A.; Forouzanfar, M.H.; Reitsma, M.B.; Sur, P.; Estep, K.; Lee, A.; Marczak, L.; Mokdad, A.H.; Moradi-Lakeh, M.; et al. Health Effects of Overweight and Obesity in 195 Countries over 25 Years. N. Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, R.; Rainer, P.Y.; Rumpler, M.; Russeil, J.; Zachara, M.; Pezoldt, J.; van Mierlo, G.; Gardeux, V.; Saelens, W.; Alpern, D.; et al. A human omentum-specific mesothelial-like stromal population inhibits adipogenesis through IGFBP2 secretion. Cell Metab. 2024, 36, 1566–1585.e9. [Google Scholar] [CrossRef]
- Sun, W.; Modica, S.; Dong, H.; Wolfrum, C. Plasticity and heterogeneity of thermogenic adipose tissue. Nat. Metab. 2021, 3, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Sharma, A.K.; Wolfrum, C. Novel insights into adipose tissue heterogeneity. Rev. Endocr. Metab. Disord. 2022, 23, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Song, T.; Kuang, S. Adipocyte dedifferentiation in health and diseases. Clin. Sci. 2019, 133, 2107–2119. [Google Scholar] [CrossRef] [PubMed]
- Stenkula, K.G.; Erlanson-Albertsson, C. Adipose cell size: Importance in health and disease. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R284–R295. [Google Scholar] [CrossRef] [PubMed]
- Skurk, T.; Alberti-Huber, C.; Herder, C.; Hauner, H. Relationship between Adipocyte Size and Adipokine Expression and Secretion. J. Clin. Endocrinol. Metab. 2007, 92, 1023–1033. [Google Scholar] [CrossRef] [PubMed]
- Villarroya, F.; Cereijo, R.; Gavaldà-Navarro, A.; Villarroya, J.; Giralt, M. Inflammation of brown/beige adipose tissues in obesity and metabolic disease. J. Intern. Med. 2018, 284, 492–504. [Google Scholar] [CrossRef]
- Weyer, C.; Foley, J.E.; Bogardus, C.; Tataranni, P.A.; Pratley, R.E. Enlarged subcutaneous abdominal adipocyte size, but not obesity itself, predicts Type II diabetes independent of insulin resistance. Diabetologia 2000, 43, 1498–1506. [Google Scholar] [CrossRef]
- Celi, F.S. Human Brown Adipose Tissue Plasticity: Hormonal and Environmental Manipulation. In Hormones, Metabolism and the Benefits of Exercise; Spiegelman, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Chen, Y.-R.; Xiao, F.; Tang, H.-N.; Wang, T.; Zhou, Y.-H.; Iqbal, J.; Yang, S.-B.; Li, L.; Zhou, H. Plasticity of adipose tissues in response to fasting and refeeding declines with aging in mice. Aging 2023, 15, 4319–4333. [Google Scholar] [CrossRef]
- Tchernof, A.; Després, J.-P. Pathophysiology of Human Visceral Obesity: An Update. Physiol. Rev. 2013, 93, 359–404. [Google Scholar] [CrossRef]
- Tchernof, A.; Desmeules, A.; Richard, C.; Laberge, P.; Daris, M.; Mailloux, J.; Rhéaume, C.; Dupont, P. Ovarian Hormone Status and Abdominal Visceral Adipose Tissue Metabolism. J. Clin. Endocrinol. Metab. 2004, 89, 3425–3430. [Google Scholar] [CrossRef]
- Halberg, N.; Khan, T.; Trujillo, M.E.; Wernstedt-Asterholm, I.; Attie, A.D.; Sherwani, S.; Wang, Z.V.; Landskroner-Eiger, S.; Dineen, S.; Magalang, U.J.; et al. Hypoxia-Inducible Factor 1α Induces Fibrosis and Insulin Resistance in White Adipose Tissue. Mol. Cell. Biol. 2009, 29, 4467–4483. [Google Scholar] [CrossRef]
- Zhang, Y.; Xie, L.; Gunasekar, S.K.; Tong, D.; Mishra, A.; Gibson, W.J.; Wang, C.; Fidler, T.; Marthaler, B.; Klingelhutz, A.; et al. SWELL1 is a regulator of adipocyte size, insulin signalling and glucose homeostasis. Nat. Cell Biol. 2017, 19, 504–517. [Google Scholar] [CrossRef] [PubMed]
- Bahceci, M.; Gokalp, D.; Bahceci, S.; Tuzcu, A.; Atmaca, S.; Arikan, S. The correlation between adiposity and adiponectin, tumor necrosis factor α, interleukin-6 and high sensitivity C-reactive protein levels. Is adipocyte size associated with inflammation in adults? J. Endocrinol. Investig. 2007, 30, 210–214. [Google Scholar] [CrossRef]
- Jernås, M.; Palming, J.; Sjöholm, K.; Jennische, E.; Svensson, P.-A.; Gabrielsson, B.G.; Levin, M.; Sjögren, A.; Rudemo, M.; Lystig, T.C.; et al. Separation of human adipocytes by size: Hypertrophic fat cells display distinct gene expression. FASEB J. 2006, 20, 1540–1542. [Google Scholar] [CrossRef]
- Keller, P.; Petrie, J.T.; De Rose, P.; Gerin, I.; Wright, W.S.; Chiang, S.-H.; Nielsen, A.R.; Fischer, C.P.; Pedersen, B.K.; MacDougald, O.A. Fat-specific Protein 27 Regulates Storage of Triacylglycerol. J. Biol. Chem. 2008, 283, 14355–14365. [Google Scholar] [CrossRef]
- He, Y.; Taub, A.B.; Yu, L.; Yao, Y.; Zhang, R.; Zahr, T.; Aaron, N.; LeSauter, J.; Fan, L.; Liu, L.; et al. PPARγ Acetylation Orchestrates Adipose Plasticity and Metabolic Rhythms. Adv. Sci. 2023, 10, e2204190. [Google Scholar] [CrossRef]
- White, U.; Ravussin, E. Dynamics of adipose tissue turnover in human metabolic health and disease. Diabetologia 2019, 62, 17–23. [Google Scholar] [CrossRef]
- Tang, Q.Q.; Lane, M.D. Adipogenesis: From Stem Cell to Adipocyte. Annu. Rev. Biochem. 2012, 81, 715–736. [Google Scholar] [CrossRef]
- Lu, Q.; Li, M.; Zou, Y.; Cao, T. Induction of adipocyte hyperplasia in subcutaneous fat depot alleviated type 2 diabetes symptoms in obese mice. Obesity 2014, 22, 1623–1631. [Google Scholar] [CrossRef]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. PPARγ signaling and metabolism: The good, the bad and the future. Nat. Med. 2013, 19, 557–566. [Google Scholar] [CrossRef]
- Arner, P.; Andersson, D.P.; Wirén, M.; Hoffstedt, J.; Näslund, E.; Thorell, A.; Rydén, M. Variations in the Size of the Major Omentum Are Primarily Determined by Fat Cell Number. J. Clin. Endocrinol. Metab. 2013, 98, E897–E901. [Google Scholar] [CrossRef]
- Pellegrinelli, V.; Carobbio, S.; Vidal-Puig, A. Adipose tissue plasticity: How fat depots respond differently to pathophysio-logical cues. Diabetologia 2016, 59, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Davis, R.J. Cell Signaling and Stress Responses. Cold Spring Harb. Perspect. Biol. 2016, 8, a006072. [Google Scholar] [CrossRef]
- Niemann, B.; Niemann, B.; Haufs-Brusberg, S.; Haufs-Brusberg, S.; Puetz, L.; Puetz, L.; Feickert, M.; Feickert, M.; Jaeckstein, M.Y.; Jaeckstein, M.Y.; et al. Apoptotic brown adipocytes enhance energy expenditure via extracellular inosine. Nature 2022, 609, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Zhang, X.; Yu, L.; Xu, Y.; Zhou, Y.; Qian, S.; Cao, X.; Ye, X.; Yang, J.; Jia, W.; et al. NF-κB regulates brown adipocyte function through suppression of ANT2. Acta Pharm. Sin. B 2022, 12, 1186–1197. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.M.S.; Castro, R.E.; Machado, M.V.; Evangelista, T.; Silvestre, A.; Costa, A.; Coutinho, J.; Carepa, F.; Cortez-Pinto, H.; Rodrigues, C.M.P. Apoptosis and insulin resistance in liver and peripheral tissues of morbidly obese patients is associated with different stages of non-alcoholic fatty liver disease. Diabetologia 2011, 54, 1788–1798. [Google Scholar] [CrossRef]
- Arner, E.; Westermark, P.O.; Spalding, K.L.; Britton, T.; Rydén, M.; Frisén, J.; Bernard, S.; Arner, P. Adipocyte Turnover: Relevance to Human Adipose Tissue Morphology. Diabetes 2010, 59, 105–109. [Google Scholar] [CrossRef]
- Sakers, A.; De Siqueira, M.K.; Seale, P.; Villanueva, C.J. Adipose-tissue plasticity in health and disease. Cell 2022, 185, 419–446. [Google Scholar] [CrossRef]
- Peng, Y.; Zhao, L.; Li, M.; Liu, Y.; Shi, Y.; Zhang, J. Plasticity of Adipose Tissues: Interconversion among White, Brown, and Beige Fat and Its Role in Energy Homeostasis. Biomolecules 2024, 14, 483. [Google Scholar] [CrossRef] [PubMed]
- Uldry, M.; Yang, W.; St-Pierre, J.; Lin, J.; Seale, P.; Spiegelman, B.M. Complementary action of the PGC-1 coactivators in mitochondrial biogenesis and brown fat differentiation. Cell Metab. 2006, 3, 333–341. [Google Scholar] [CrossRef]
- Seale, P.; Bjork, B.; Yang, W.; Kajimura, S.; Chin, S.; Kuang, S.; Scimè, A.; Devarakonda, S.; Conroe, H.M.; Erdjument-Bromage, H.; et al. PRDM16 controls a brown fat/skeletal muscle switch. Nature 2008, 454, 961–967. [Google Scholar] [CrossRef]
- Wang, W.; Kissig, M.; Rajakumari, S.; Huang, L.; Lim, H.-W.; Won, K.-J.; Seale, P. Ebf2 is a selective marker of brown and beige adipogenic precursor cells. Proc. Natl. Acad. Sci. USA 2014, 111, 14466–14471. [Google Scholar] [CrossRef]
- Qian, S.-W.; Tang, Y.; Li, X.; Liu, Y.; Zhang, Y.-Y.; Huang, H.-Y.; Xue, R.-D.; Yu, H.-Y.; Guo, L.; Gao, H.-D.; et al. BMP4-mediated brown fat-like changes in white adipose tissue alter glucose and energy homeostasis. Proc. Natl. Acad. Sci. USA 2013, 110, E798–E807. [Google Scholar] [CrossRef]
- Tseng, Y.-H.; Kokkotou, E.; Schulz, T.J.; Huang, T.L.; Winnay, J.N.; Taniguchi, C.M.; Tran, T.T.; Suzuki, R.; Espinoza, D.O.; Yamamoto, Y.; et al. New role of bone morphogenetic protein 7 in brown adipogenesis and energy expenditure. Nature 2008, 454, 1000–1004. [Google Scholar] [CrossRef]
- Cai, J.; Quan, Y.; Zhu, S.; Lin, J.; Zhang, Q.; Liu, J.; Liang, Z.; Liao, Y.; Jiang, W.; He, Y.; et al. The browning and mobilization of subcutaneous white adipose tissue supports efficient skin repair. Cell Metab. 2024, 36, 1287–1301.e7. [Google Scholar] [CrossRef]
- Shi, L.; Tao, Z.; Zheng, L.; Yang, J.; Hu, X.; Scott, K.; de Kloet, A.; Krause, E.; Collins, J.F.; Cheng, Z. FoxO1 regulates adipose transdifferentiation and iron influx by mediating Tgfβ1 signaling pathway. Redox Biol. 2023, 63, 102727. [Google Scholar] [CrossRef]
- Yadav, H.; Quijano, C.; Kamaraju, A.K.; Gavrilova, O.; Malek, R.; Chen, W.; Zerfas, P.; Zhigang, D.; Wright, E.C.; Stuelten, C.; et al. Protection from Obesity and Diabetes by Blockade of TGF-β/Smad3 Signaling. Cell Metab. 2011, 14, 67–79. [Google Scholar] [CrossRef]
- Bi, P.; Shan, T.; Liu, W.; Yue, F.; Yang, X.; Liang, X.-R.; Wang, J.; Li, J.; Carlesso, N.; Liu, X.; et al. Inhibition of Notch signaling promotes browning of white adipose tissue and ameliorates obesity. Nat. Med. 2014, 20, 911–918. [Google Scholar] [CrossRef]
- Nanduri, R. Epigenetic Regulators of White Adipocyte Browning. Epigenomes 2021, 5, 3. [Google Scholar] [CrossRef]
- Lorente-Cebrián, S.; González-Muniesa, P.; Milagro, F.I.; Martínez, J.A. MicroRNAs and other non-coding RNAs in adipose tissue and obesity: Emerging roles as biomarkers and therapeutic targets. Clin. Sci. 2019, 133, 23–40. [Google Scholar] [CrossRef]
- Zhang, H.; Guan, M.; Townsend, K.L.; Huang, T.L.; An, D.; Yan, X.; Xue, R.; Schulz, T.J.; Winnay, J.; Mori, M.; et al. Micro RNA-455 regulates brown adipogenesis via a novel HIF 1an-AMPK—PGC 1α signaling network. Embo Rep. 2015, 16, 1378–1393. [Google Scholar] [CrossRef]
- He, L.; Tang, M.; Xiao, T.; Liu, H.; Liu, W.; Li, G.; Zhang, F.; Xiao, Y.; Zhou, Z.; Liu, F.; et al. Obesity-Associated miR-199a/214 Cluster Inhibits Adipose Browning via PRDM16–PGC-1α Transcriptional Network. Diabetes 2018, 67, 2585–2600. [Google Scholar] [CrossRef]
- Gaudet, A.D.; Fonken, L.K.; Gushchina, L.V.; Aubrecht, T.G.; Maurya, S.K.; Periasamy, M.; Nelson, R.J.; Popovich, P.G. miR-155 Deletion in Female Mice Prevents Diet-Induced Obesity. Sci. Rep. 2016, 6, 22862. [Google Scholar] [CrossRef]
- Fischer, C.; Seki, T.; Lim, S.; Nakamura, M.; Andersson, P.; Yang, Y.; Honek, J.; Wang, Y.; Gao, Y.; Chen, F.; et al. A miR-327–FGF10–FGFR2-mediated autocrine signaling mechanism controls white fat browning. Nat. Commun. 2017, 8, 2079. [Google Scholar] [CrossRef]
- Lv, Y.-F.; Yu, J.; Sheng, Y.-L.; Huang, M.; Kong, X.-C.; Di, W.-J.; Liu, J.; Zhou, H.; Liang, H.; Ding, G.-X. Glucocorticoids Suppress the Browning of Adipose Tissue via miR-19b in Male Mice. Endocrinology 2018, 159, 310–322. [Google Scholar] [CrossRef]
- Kim, S.; Park, J.W.; Lee, M.G.; Nam, K.H.; Park, J.H.; Oh, H.; Lee, J.; Han, J.; Yi, S.A.; Han, J. Reversine promotes browning of white adipocytes by suppressing miR-133a. J. Cell. Physiol. 2019, 234, 3800–3813. [Google Scholar] [CrossRef]
- Fu, T.; Seok, S.; Choi, S.; Huang, Z.; Suino-Powell, K.; Xu, H.E.; Kemper, B.; Kemper, J.K. MicroRNA 34a Inhibits Beige and Brown Fat Formation in Obesity in Part by Suppressing Adipocyte Fibroblast Growth Factor 21 Signaling and SIRT1 Function. Mol. Cell. Biol. 2014, 34, 4130–4142. [Google Scholar] [CrossRef]
- Shi, F.; Collins, S. Second messenger signaling mechanisms of the brown adipocyte thermogenic program: An integrative perspective. Horm. Mol. Biol. Clin. Investig. 2017, 31, 20170062. [Google Scholar] [CrossRef]
- Collins, S. β-adrenoceptor signaling networks in adipocytes for recruiting stored fat and energy expenditure. Front. Endocrinol. 2011, 2, 102. [Google Scholar] [CrossRef]
- Lass, A.; Zimmermann, R.; Oberer, M.; Zechner, R. Lipolysis—A highly regulated multi-enzyme complex mediates the catabolism of cellular fat stores. Prog. Lipid Res. 2011, 50, 14–27. [Google Scholar] [CrossRef]
- Cannon, B.; Nedergaard, J. Brown Adipose Tissue: Function and Physiological Significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef]
- Leiva, M.; Matesanz, N.; Pulgarín-Alfaro, M.; Nikolic, I.; Sabio, G. Uncovering the Role of p38 Family Members in Adipose Tissue Physiology. Front. Endocrinol. 2020, 11, 572089. [Google Scholar] [CrossRef]
- Wang, S.; Wang, X.; Ye, Z.; Xu, C.; Zhang, M.; Ruan, B.; Wei, M.; Jiang, Y.; Zhang, Y.; Wang, L.; et al. Curcumin promotes browning of white adipose tissue in a norepinephrine-dependent way. Biochem. Biophys. Res. Commun. 2015, 466, 247–253. [Google Scholar] [CrossRef]
- Chen, Y.; Ikeda, K.; Yoneshiro, T.; Scaramozza, A.; Tajima, K.; Wang, Q.; Kim, K.; Shinoda, K.; Sponton, C.H.; Brown, Z.; et al. Thermal stress induces glycolytic beige fat formation via a myogenic state. Nature 2019, 565, 180–185. [Google Scholar] [CrossRef]
- Martínez-Sánchez, N.; Moreno-Navarrete, J.M.; Contreras, C.; Rial-Pensado, E.; Fernø, J.; Nogueiras, R.; Diéguez, C.; Fernández-Real, J.-M.; López, M. Thyroid hormones induce browning of white fat. J. Endocrinol. 2017, 232, 351–362. [Google Scholar] [CrossRef]
- Lin, J.Z.; Martagón, A.J.; Cimini, S.L.; Gonzalez, D.D.; Tinkey, D.W.; Biter, A.; Baxter, J.D.; Webb, P.; Gustafsson, J.; Hartig, S.M.; et al. Pharmacological Activation of Thyroid Hormone Receptors Elicits a Functional Conversion of White to Brown Fat. Cell Rep. 2015, 13, 1528–1537. [Google Scholar] [CrossRef]
- Finan, B.; Clemmensen, C.; Zhu, Z.; Stemmer, K.; Gauthier, K.; Müller, L.; Moreth, K.; Neff, F.; Perez-Tilve, D.; Fischer, K.; et al. Chemical Hybridization of Glucagon and Thyroid Hormone Optimizes Therapeutic Impact for Metabolic Disease. Cell 2016, 167, 843–857.e14. [Google Scholar] [CrossRef]
- Mahajan, R.D.; Patra, S.K. Irisin, a Novel Myokine Responsible for Exercise Induced Browning of White Adipose Tissue. Indian J. Clin. Biochem. 2013, 28, 102–103. [Google Scholar] [CrossRef]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Castillo-Quan, J.I. From white to brown fat through the PGC-1α-dependent myokine irisin: Implications for diabetes and obesity. Dis. Model. Mech. 2012, 5, 293–295. [Google Scholar] [CrossRef]
- Cao, L.; Choi, E.Y.; Liu, X.; Martin, A.; Wang, C.; Xu, X.; During, M.J. White to Brown Fat Phenotypic Switch Induced by Genetic and Environmental Activation of a Hypothalamic-Adipocyte Axis. Cell Metab. 2011, 14, 324–338. [Google Scholar] [CrossRef]
- Lizcano, F. The Beige Adipocyte as a Therapy for Metabolic Diseases. Int. J. Mol. Sci. 2019, 20, 5058. [Google Scholar] [CrossRef]
- Foltz, I.N.; Hu, S.; King, C.; Wu, X.; Yang, C.; Wang, W.; Weiszmann, J.; Stevens, J.; Chen, J.S.; Nuanmanee, N.; et al. Treating Diabetes and Obesity with an FGF21-Mimetic Antibody Activating the βKlotho/FGFR1c Receptor Complex. Sci. Transl. Med. 2012, 4, 162ra153. [Google Scholar] [CrossRef]
- Lee, P.; Brychta, R.J.; Linderman, J.; Smith, S.; Chen, K.Y.; Celi, F.S. Mild cold exposure modulates fibroblast growth factor 21 (FGF21) diurnal rhythm in humans: Relationship between FGF21 levels, lipolysis, and cold-induced thermogenesis. J. Clin. Endocrinol. Metab. 2013, 98, 98–102. [Google Scholar] [CrossRef]
- Morón-Ros, S.; Uriarte, I.; Berasain, C.; Avila, M.A.; Sabater-Masdeu, M.; Moreno-Navarrete, J.M.; Fernández-Real, J.M.; Giralt, M.; Villarroya, F.; Gavaldà-Navarro, A. FGF15/19 is required for adipose tissue plasticity in response to thermogenic adaptations. Mol. Metab. 2021, 43, 101113. [Google Scholar] [CrossRef]
- Bartelt, A.; Bruns, O.T.; Reimer, R.; Hohenberg, H.; Ittrich, H.; Peldschus, K.; Kaul, M.G.; Tromsdorf, U.I.; Weller, H.; Waurisch, C.; et al. Brown adipose tissue activity controls triglyceride clearance. Nat. Med. 2011, 17, 200–205. [Google Scholar] [CrossRef]
- Abdullahi, A.; Knuth, C.M.; Auger, C.; Sivayoganathan, T.; Parousis, A.; Jeschke, M.G. Adipose browning response to burn trauma is impaired with aging. J. Clin. Investig. 2021, 6, 143451. [Google Scholar] [CrossRef]
- Li, G.; Xie, C.; Lu, S.; Nichols, R.G.; Tian, Y.; Li, L.; Patel, D.; Ma, Y.; Brocker, C.N.; Yan, T.; et al. Intermittent Fasting Promotes White Adipose Browning and Decreases Obesity by Shaping the Gut Microbiota. Cell Metab. 2017, 26, 672–685.e4. [Google Scholar] [CrossRef]
- Sahuri-Arisoylu, M.; Brody, L.P.; Parkinson, J.R.; Parkes, H.; Navaratnam, N.; Miller, A.D.; Thomas, E.L.; Frost, G.; Bell, J.D. Reprogramming of hepatic fat accumulation and ‘browning’ of adipose tissue by the short-chain fatty acid acetate. Int. J. Obes. 2016, 40, 955–963. [Google Scholar] [CrossRef]
- Petersen, C.; Nielsen, M.D.; Andersen, E.S.; Basse, A.L.; Isidor, M.S.; Markussen, L.K.; Viuff, B.M.; Lambert, I.H.; Hansen, J.B.; Pedersen, S.F. MCT1 and MCT4 Expression and Lactate Flux Activity Increase During White and Brown Adipogenesis and Impact Adipocyte Metabolism. Sci. Rep. 2017, 7, 13101. [Google Scholar] [CrossRef]
- Rabiee, A. Beige Fat Maintenance; Toward a Sustained Metabolic Health. Front. Endocrinol. 2020, 11, 634. [Google Scholar] [CrossRef] [PubMed]
- Roh, H.C.; Tsai, L.T.; Shao, M.; Tenen, D.; Shen, Y.; Kumari, M.; Lyubetskaya, A.; Jacobs, C.; Dawes, B.; Gupta, R.K.; et al. Warming Induces Significant Reprogramming of Beige, but Not Brown, Adipocyte Cellular Identity. Cell Metab. 2018, 27, 1121–1137.e5. [Google Scholar] [CrossRef]
- Morigny, P.; Boucher, J.; Arner, P.; Langin, D. Lipid and glucose metabolism in white adipocytes: Pathways, dysfunction and therapeutics. Nat. Rev. Endocrinol. 2021, 17, 276–295. [Google Scholar] [CrossRef] [PubMed]
- Giordano, A.; Smorlesi, A.; Frontini, A.; Barbatelli, G.; Cinti, S. Mechanisms in endocrinology: White, brown and pink adipocytes: The extraordinary plasticity of the adipose organ. Eur. J. Endocrinol. 2014, 170, R159–R171. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wang, J.; Dai, H.; Duan, Y.; An, Y.; Shi, L.; Lv, Y.; Li, H.; Wang, C.; Ma, Q.; et al. Brown and beige adipose tissue: A novel therapeutic strategy for obesity and type 2 diabetes mellitus. Adipocyte 2021, 10, 48–65. [Google Scholar] [CrossRef] [PubMed]
- Bordicchia, M.; Liu, D.; Amri, E.-Z.; Ailhaud, G.; Dessì-Fulgheri, P.; Zhang, C.; Takahashi, N.; Sarzani, R.; Collins, S. Cardiac natriuretic peptides act via p38 MAPK to induce the brown fat thermogenic program in mouse and human adipocytes. J. Clin. Investig. 2012, 122, 1022–1036. [Google Scholar] [CrossRef]
- Liang, X.; Pan, J.; Cao, C.; Zhang, L.; Zhao, Y.; Fan, Y.; Li, K.; Tao, C.; Wang, Y.; Liang, X.; et al. Transcriptional Response of Subcutaneous White Adipose Tissue to Acute Cold Exposure in Mice. Int. J. Mol. Sci. 2019, 20, 3968. [Google Scholar] [CrossRef] [PubMed]
- Merlin, J.; Sato, M.; Nowell, C.; Pakzad, M.; Fahey, R.; Gao, J.; Dehvari, N.; Summers, R.J.; Bengtsson, T.; Evans, B.A.; et al. The PPARγ agonist rosiglitazone promotes the induction of brite adipocytes, increasing β-adrenoceptor-mediated mitochondrial function and glucose uptake. Cell. Signal. 2018, 42, 54–66. [Google Scholar] [CrossRef]
- Krause, K. Novel Aspects of White Adipose Tissue Browning by Thyroid Hormones. Exp. Clin. Endocrinol. Diabetes 2020, 128, 446–449. [Google Scholar] [CrossRef]
- Cairó, M.; Villarroya, J. The role of autophagy in brown and beige adipose tissue plasticity. J. Physiol. Biochem. 2020, 76, 213–226. [Google Scholar] [CrossRef]
- Crewe, C.; An, Y.A.; Scherer, P.E. The ominous triad of adipose tissue dysfunction: Inflammation, fibrosis, and impaired angiogenesis. J. Clin. Investig. 2017, 127, 74–82. [Google Scholar] [CrossRef]
- Corvera, S.; Gealekman, O. Adipose tissue angiogenesis: Impact on obesity and type-2 diabetes. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2014, 1842, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, A.B.; Massihnia, D.; Grisafi, F.; Castiglia, M.; Toia, F.; Montesano, L.; Russo, A.; Moschella, F.; Cordova, A. Adipose tissue, angiogenesis and angio-MIR under physiological and pathological conditions. Eur. J. Cell Biol. 2019, 98, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Hammarstedt, A.; Gogg, S.; Hedjazifar, S.; Nerstedt, A.; Smith, U. Impaired Adipogenesis and Dysfunctional Adipose Tissue in Human Hypertrophic Obesity. Physiol. Rev. 2018, 98, 1911–1941. [Google Scholar] [CrossRef] [PubMed]
- Briot, A.; Decaunes, P.; Volat, F.; Belles, C.; Coupaye, M.; Ledoux, S.; Bouloumié, A. Senescence Alters PPARγ (Peroxisome Proliferator–Activated Receptor Gamma)-Dependent Fatty Acid Handling in Human Adipose Tissue Microvascular Endothelial Cells and Favors Inflammation. Arter. Thromb. Vasc. Biol. 2018, 38, 1134–1146. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.W.; Heo, S.C.; Jeong, G.O.; Yoon, J.W.; Mo, W.M.; Lee, M.J.; Jang, I.-H.; Kwon, S.M.; Lee, J.S.; Kim, J.H. Tumor necrosis factor-α-activated mesenchymal stem cells promote endothelial progenitor cell homing and angiogenesis. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2013, 1832, 2136–2144. [Google Scholar] [CrossRef] [PubMed]
- Jetten, N.; Verbruggen, S.; Gijbels, M.J.; Post, M.J.; De Winther, M.P.J.; Donners, M.M.P.C. Anti-inflammatory M2, but not pro-inflammatory M1 macrophages promote angiogenesis in vivo. Angiogenesis 2014, 17, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Three-Dimensional Volume Fluorescence-Imaging of Vascular Plasticity in Adipose Tissues—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/29914852/ (accessed on 29 October 2022).
- Meng, X.; Qian, X.; Ding, X.; Wang, W.; Yin, X.; Zhuang, G.; Zeng, W. Eosinophils regulate intra-adipose axonal plasticity. Proc. Natl. Acad. Sci. USA 2022, 119, e2112281119. [Google Scholar] [CrossRef]
- Blaszkiewicz, M.; Wood, E.; Koizar, S.; Willows, J.; Anderson, R.; Tseng, Y.-H.; Godwin, J.; Townsend, K.L. The involvement of neuroimmune cells in adipose innervation. Mol. Med. 2020, 26, 126. [Google Scholar] [CrossRef]
- Cao, Y.; Wang, H.; Zeng, W. Whole-tissue 3D imaging reveals intra-adipose sympathetic plasticity regulated by NGF-TrkA signal in cold-induced beiging. Protein Cell 2018, 9, 527–539. [Google Scholar] [CrossRef]
- Wang, P.; Loh, K.H.; Wu, M.; Morgan, D.A.; Schneeberger, M.; Yu, X.; Chi, J.; Kosse, C.; Kim, D.; Rahmouni, K.; et al. A leptin–BDNF pathway regulating sympathetic innervation of adipose tissue. Nature 2020, 583, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Mishra, G.; Townsend, K.L. The metabolic and functional roles of sensory nerves in adipose tissues. Nat. Metab. 2023, 5, 1461–1474. [Google Scholar] [CrossRef] [PubMed]
- Blaszkiewicz, M.; Willows, J.W.; Dubois, A.L.; Waible, S.; DiBello, K.; Lyons, L.L.; Johnson, C.P.; Paradie, E.; Banks, N.; Motyl, K.; et al. Neuropathy and neural plasticity in the subcutaneous white adipose depot. PLoS ONE 2019, 14, e0221766. [Google Scholar] [CrossRef] [PubMed]
- Jaitin, D.A.; Adlung, L.; Thaiss, C.A.; Weiner, A.; Li, B.; Descamps, H.; Lundgren, P.; Bleriot, C.; Liu, Z.; Deczkowska, A.; et al. Lipid-Associated Macrophages Control Metabolic Homeostasis in a Trem2-Dependent Manner. Cell 2019, 178, 686–698.e14. [Google Scholar] [CrossRef] [PubMed]
- Sárvári, A.K.; Van Hauwaert, E.L.; Markussen, L.K.; Gammelmark, E.; Marcher, A.-B.; Ebbesen, M.F.; Nielsen, R.; Brewer, J.R.; Madsen, J.G.S.; Mandrup, S. Plasticity of Epididymal Adipose Tissue in Response to Diet-Induced Obesity at Single-Nucleus Resolution. Cell Metab. 2021, 33, 437–453.e5. [Google Scholar] [CrossRef]
- Liang, W.; Qi, Y.; Yi, H.; Mao, C.; Meng, Q.; Wang, H.; Zheng, C. The Roles of Adipose Tissue Macrophages in Human Disease. Front. Immunol. 2022, 13, 908749. [Google Scholar] [CrossRef]
- Blüher, M.; Bashan, N.; Shai, I.; Harman-Boehm, I.; Tarnovscki, T.; Avinaoch, E.; Stumvoll, M.; Dietrich, A.; Klöting, N.; Rudich, A. Activated Ask1-MKK4-p38MAPK/JNK stress signaling pathway in human omental fat tissue may link mac-rophage infiltration to whole-body Insulin sensitivity. J. Clin. Endocrinol. Metab. 2009, 94, 2507–2515. [Google Scholar] [CrossRef]
- Park, K.S. Aucubin, a naturally occurring iridoid glycoside inhibits TNF-α-induced inflammatory responses through suppression of NF-κB activation in 3T3-L1 adipocytes. Cytokine 2013, 62, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.; Tang, Y.; Tang, Q.-Q. Adipose tissue plasticity and the pleiotropic roles of BMP signaling. J. Biol. Chem. 2021, 296, 100678. [Google Scholar] [CrossRef]
- Glass, C.K.; Olefsky, J.M. Inflammation and Lipid Signaling in the Etiology of Insulin Resistance. Cell Metab. 2012, 15, 635–645. [Google Scholar] [CrossRef]
- Odegaard, J.I.; Ricardo-Gonzalez, R.R.; Eagle, A.R.; Vats, D.; Morel, C.R.; Goforth, M.H.; Subramanian, V.; Mukundan, L.; Ferrante, A.W.; Chawla, A. Alternative M2 Activation of Kupffer Cells by PPARδ Ameliorates Obesity-Induced Insulin Resistance. Cell Metab. 2008, 7, 496–507. [Google Scholar] [CrossRef]
- Li, C.; Ying, W.; Huang, Z.; Brehm, T.; Morin, A.; Vella, A.T.; Zhou, B. IRF6 Regulates Alternative Activation by Suppressing PPARγ in Male Murine Macrophages. Endocrinology 2017, 158, 2837–2847. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, K.; Parker, J.L.; Ouchi, N.; Higuchi, A.; Vita, J.A.; Gokce, N.; Pedersen, A.A.; Kalthoff, C.; Tullin, S.; Sams, A.; et al. Adiponectin Promotes Macrophage Polarization toward an Anti-inflammatory Phenotype. J. Biol. Chem. 2010, 285, 6153–6160. [Google Scholar] [CrossRef] [PubMed]
- Kumari, M.; Heeren, J.; Scheja, L. Regulation of immunometabolism in adipose tissue. Semin. Immunopathol. 2018, 40, 189–202. [Google Scholar] [CrossRef]
- Abdullahi, A.P.; Auger, C.; Stanojcic, M.P.; Patsouris, D.; Parousis, A.M.; Epelman, S.; Jeschke, M.G. Alternatively Activated Macrophages Drive Browning of White Adipose Tissue in Burns. Ann. Surg. 2019, 269, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.D.; Qiu, Y.; Cui, X.; Goh, Y.P.S.; Mwangi, J.; David, T.; Mukundan, L.; Brombacher, F.; Locksley, R.M.; Chawla, A. Alternatively activated macrophages produce catecholamines to sustain adaptive thermogenesis. Nature 2011, 480, 104–108. [Google Scholar] [CrossRef]
- Rao, R.R.; Long, J.Z.; White, J.P.; Svensson, K.J.; Lou, J.; Lokurkar, I.; Jedrychowski, M.P.; Ruas, J.L.; Wrann, C.D.; Lo, J.C.; et al. Meteorin-like Is a Hormone that Regulates Immune-Adipose Interactions to Increase Beige Fat Thermogenesis. Cell 2014, 157, 1279–1291. [Google Scholar] [CrossRef] [PubMed]
- Caslin, H.L.; Bhanot, M.; Bolus, W.R.; Hasty, A.H. Adipose tissue macrophages: Unique polarization and bioenergetics in obesity. Immunol. Rev. 2020, 295, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Winer, D.A.; Winer, S.; Shen, L.; Wadia, P.P.; Yantha, J.; Paltser, G.; Tsui, H.; Wu, P.; Davidson, M.G.; Alonso, M.N.; et al. B cells promote insulin resistance through modulation of T cells and production of pathogenic IgG antibodies. Nat. Med. 2011, 17, 610–617. [Google Scholar] [CrossRef]
- Nishimura, S.; Manabe, I.; Nagasaki, M.; Eto, K.; Yamashita, H.; Ohsugi, M.; Otsu, M.; Hara, K.; Ueki, K.; Sugiura, S.; et al. CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med. 2009, 15, 914–920. [Google Scholar] [CrossRef]
- Winer, S.; Chan, Y.; Paltser, G.; Truong, D.; Tsui, H.; Bahrami, J.; Dorfman, R.; Wang, Y.; Zielenski, J.; Mastronardi, F.; et al. Normalization of obesity-associated insulin resistance through immunotherapy. Nat. Med. 2009, 15, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Cipolletta, D.; Feuerer, M.; Li, A.; Kamei, N.; Lee, J.; Shoelson, S.E.; Benoist, C.; Mathis, D. PPAR-γ is a major driver of the accumulation and phenotype of adipose tissue Treg cells. Nature 2012, 486, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Lynch, L.; Michelet, X.; Zhang, S.; Brennan, P.J.; Moseman, A.; Lester, C.; Besra, G.; Vomhof-Dekrey, E.E.; Tighe, M.; Koay, H.-F.; et al. Regulatory iNKT cells lack expression of the transcription factor PLZF and control the homeostasis of Treg cells and macrophages in adipose tissue. Nat. Immunol. 2015, 16, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.M. Subcutaneous and visceral adipose tissue: Structural and functional differences. Obes. Rev. 2010, 11, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Pramyothin, P.; Karastergiou, K.; Fried, S.K. Deconstructing the roles of glucocorticoids in adipose tissue biology and the development of central obesity. Biochim. Biophys. Acta (BBA) 2014, 1842, 473–481. [Google Scholar] [CrossRef]
- Mancuso, P.; Mancuso, P.; Bouchard, B.; Bouchard, B. The Impact of Aging on Adipose Function and Adipokine Synthesis. Front. Endocrinol. 2019, 10, 137. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, J.C.; Champagne, C.M.; de Jonge, L.; Xie, H.; Smith, S.R. Increased visceral fat and decreased energy expenditure during the menopausal transition. Int. J. Obes. 2008, 32, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Dumesic, D.A.; Akopians, A.L.; Madrigal, V.K.; Ramirez, E.; Margolis, D.J.; Sarma, M.K.; Thomas, A.M.; Grogan, T.R.; Haykal, R.; Schooler, T.A.; et al. Hyperandrogenism Accompanies Increased Intra-Abdominal Fat Storage in Normal Weight Polycystic Ovary Syndrome Women. J. Clin. Endocrinol. Metab. 2016, 101, 4178–4188. [Google Scholar] [CrossRef]
- Wang, G.; Song, A.; Bae, M.; Wang, Q.A. Adipose Tissue Plasticity in Aging. Compr. Physiol. 2022, 12, 4119–4132. [Google Scholar] [CrossRef]
- Kim, J.-Y.; van de Wall, E.; Laplante, M.; Azzara, A.; Trujillo, M.E.; Hofmann, S.M.; Schraw, T.; Durand, J.L.; Li, H.; Li, G.; et al. Obesity-associated improvements in metabolic profile through expansion of adipose tissue. J. Clin. Investig. 2007, 117, 2621–2637. [Google Scholar] [CrossRef]
- Kim, Y.J.; Ryu, R.; Choi, J.-Y.; Choi, M.-S. Platycodon grandiflorus Root Ethanol Extract Induces Lipid Excretion, Lipolysis, and Thermogenesis in Diet-Induced Obese Mice. J. Med. Food 2019, 22, 1100–1109. [Google Scholar] [CrossRef]
- Zamboni, M.; Rossi, A.P.; Fantin, F.; Zamboni, G.; Chirumbolo, S.; Zoico, E.; Mazzali, G. Adipose tissue, diet and aging. Mech. Ageing Dev. 2014, 136–137, 129–137. [Google Scholar] [CrossRef]
- Eckel-Mahan, K.; Latre, A.R.; Kolonin, M.G. Adipose Stromal Cell Expansion and Exhaustion: Mechanisms and Consequences. Cells 2020, 9, 863. [Google Scholar] [CrossRef] [PubMed]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Ge, G.; Yang, P.; Wang, L.; Qiao, Y.; Pan, G.; Yang, H.; Bai, J.; Cui, W.; Geng, D. An Update on Adipose-Derived Stem Cells for Regenerative Medicine: Where Challenge Meets Opportunity. Adv. Sci. 2023, 10, e2207334. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Han, N.; Zhang, P.; Jiang, B. Local transplantation of osteogenic pre-differentiated autologous adipose-derived mesenchymal stem cells may accelerate non-union fracture healing with limited pro-metastatic potency. Int. J. Clin. Exp. Med. 2015, 8, 1406–1410. [Google Scholar]
- Tontonoz, P.; Hu, E.; Spiegelman, B.M. Stimulation of adipogenesis in fibroblasts by PPARγ2, a lipid-activated transcription factor. Cell 1994, 79, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Darlington, G.J.; Ross, S.E.; MacDougald, O.A. The Role of C/EBP Genes in Adipocyte Differentiation. J. Biol. Chem. 1998, 273, 30057–30060. [Google Scholar] [CrossRef]
- Tang, Q.-Q.; Otto, T.C.; Lane, M.D. Mitotic clonal expansion: A synchronous process required for adipogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 44–49. [Google Scholar] [CrossRef]
- Shan, B.; Barker, C.S.; Shao, M.; Zhang, Q.; Gupta, R.K.; Wu, Y. Multilayered omics reveal sex- and depot-dependent adipose progenitor cell heterogeneity. Cell Metab. 2022, 34, 783–799.e7. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, S. Role of kruppel-like transcription factors in adipogenesis. Dev. Biol. 2013, 373, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Birsoy, K.; Chen, Z.; Friedman, J. Transcriptional Regulation of Adipogenesis by KLF4. Cell Metab. 2008, 7, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Choy, L.; Derynck, R. Transforming Growth Factor-β Inhibits Adipocyte Differentiation by Smad3 Interacting with CCAAT/Enhancer-binding Protein (C/EBP) and Repressing C/EBP Transactivation Function. J. Biol. Chem. 2003, 278, 9609–9619. [Google Scholar] [CrossRef]
- BMP Signaling Pathway Is Required for Commitment of C3H10T1/2 Pluripotent Stem Cells to the Adipocyte Lineage—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/19620713/ (accessed on 17 October 2022).
- Lijnen, H.; Maquoi, E.; Hansen, L.; Van Hoef, B.; Frederix, L.; Collen, D. Matrix Metalloproteinase Inhibition Impairs Adipose Tissue Development in Mice. Arter. Thromb. Vasc. Biol. 2002, 22, 374–379. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Kano, K.; Kondo, D.; Fukuda, N.; Iribe, Y.; Tanaka, N.; Matsubara, Y.; Sakuma, T.; Satomi, A.; Otaki, M.; et al. Mature adipocyte-derived dedifferentiated fat cells exhibit multilineage potential. J. Cell. Physiol. 2008, 215, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Plikus, M.V.; Guerrero-Juarez, C.F.; Ito, M.; Li, Y.R.; Dedhia, P.H.; Zheng, Y.; Shao, M.; Gay, D.L.; Ramos, R.; Hsi, T.-C.; et al. Regeneration of fat cells from myofibroblasts during wound healing. Science 2017, 355, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Bochet, L.; Lehuédé, C.; Dauvillier, S.; Wang, Y.Y.; Dirat, B.; Laurent, V.; Dray, C.; Guiet, R.; Maridonneau-Parini, I.; Le Gonidec, S.; et al. Adipocyte-derived fibroblasts promote tumor progression and contribute to the desmoplastic reaction in breast cancer. Cancer Res. 2013, 73, 5657–5668. [Google Scholar] [CrossRef]
- Bi, P.; Yue, F.; Karki, A.; Castro, B.; Wirbisky, S.E.; Wang, C.; Durkes, A.; Elzey, B.D.; Andrisani, O.M.; Bidwell, C.A.; et al. Notch activation drives adipocyte dedifferentiation and tumorigenic transformation in mice. J. Exp. Med. 2016, 213, 2019–2037. [Google Scholar] [CrossRef] [PubMed]
- Zoico, E.; Darra, E.; Rizzatti, V.; Budui, S.; Franceschetti, G.; Mazzali, G.; Rossi, A.P.; Fantin, F.; Menegazzi, M.; Cinti, S.; et al. Adipocytes WNT5a mediated dedifferentiation: A possible target in pancreatic cancer microenvironment. Oncotarget 2016, 7, 20223–20235. [Google Scholar] [CrossRef]
- Marangoni, R.G.; Korman, B.D.; Wei, J.; Wood, T.A.; Graham, L.V.; Whitfield, M.L.; Scherer, P.E.; Tourtellotte, W.G.; Varga, J. Myofibroblasts in Murine Cutaneous Fibrosis Originate from Adiponectin-Positive Intradermal Progenitors. Arthritis Rheumatol. 2015, 67, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- Ohgo, S.; Hasegawa, S.; Hasebe, Y.; Mizutani, H.; Nakata, S.; Akamatsu, H. Bleomycin inhibits adipogenesis and accelerates fibrosis in the subcutaneous adipose layer through TGF-β1. Exp. Dermatol. 2013, 22, 769–771. [Google Scholar] [CrossRef] [PubMed]
- Halaas, J.L.; Gajiwala, K.S.; Maffei, M.; Cohen, S.L.; Chait, B.T.; Rabinowitz, D.; Lallone, R.L.; Burley, S.K.; Friedman, J.M. Weight-Reducing Effects of the Plasma Protein Encoded by the obese Gene. Science 1995, 269, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, I.S.; O’Rahilly, S. Mutations in ligands and receptors of the leptin–melanocortin pathway that lead to obesity. Nat. Clin. Pract. Endocrinol. Metab. 2008, 4, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Stern, J.H.; Rutkowski, J.M.; Scherer, P.E. Adiponectin, Leptin, and Fatty Acids in the Maintenance of Metabolic Homeostasis through Adipose Tissue Crosstalk. Cell Metab. 2016, 23, 770–784. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Pirzgalska, R.M.; Pereira, M.M.; Kubasova, N.; Barateiro, A.; Seixas, E.; Lu, Y.-H.; Kozlova, A.; Voss, H.; Martins, G.G.; et al. Sympathetic Neuro-adipose Connections Mediate Leptin-Driven Lipolysis. Cell 2015, 163, 84–94. [Google Scholar] [CrossRef]
- Li, R.-M.; Chen, S.-Q.; Zeng, N.-X.; Zheng, S.-H.; Guan, L.; Liu, H.-M.; Zhou, L.-Q.; Xu, J.-W. Browning of Abdominal Aorta Perivascular Adipose Tissue Inhibits Adipose Tissue Inflammation. Metab. Syndr. Relat. Disord. 2017, 15, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose Expression of Tumor Necrosis Factor-α: Direct Role in Obesity-Linked Insulin Resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Steppan, C.M.; Bailey, S.T.; Bhat, S.; Brown, E.J.; Banerjee, R.R.; Wright, C.M.; Patel, H.R.; Ahima, R.S.; Lazar, M.A. The hormone resistin links obesity to diabetes. Nature 2001, 409, 307–312. [Google Scholar] [CrossRef]
- Yang, Q.; Graham, T.E.; Mody, N.; Preitner, F.; Peroni, O.D.; Zabolotny, J.M.; Kotani, K.; Quadro, L.; Kahn, B.B. Serum retinol binding protein 4 contributes to insulin resistance in obesity and type 2 diabetes. Nature 2005, 436, 356–362. [Google Scholar] [CrossRef]
- Gesmundo, I.; Pardini, B.; Gargantini, E.; Gamba, G.; Birolo, G.; Fanciulli, A.; Banfi, D.; Congiusta, N.; Favaro, E.; Deregibus, M.C.; et al. Adipocyte-derived extracellular vesicles regulate survival and function of pancreatic β cells. J. Clin. Investig. 2021, 6, 141962. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, A.; Lazic, M.; Armando, A.M.; Phillips, S.A.; Katebian, R.; Maraka, S.; Quehenberger, O.; Sears, D.D.; Feldstein, A.E. Circulating adipocyte-derived extracellular vesicles are novel markers of metabolic stress. J. Mol. Med. 2016, 94, 1241–1253. [Google Scholar] [CrossRef]
- Ferrante, S.C.; Nadler, E.P.; Pillai, D.K.; Hubal, M.J.; Wang, Z.; Wang, J.M.; Gordish-Dressman, H.; Koeck, E.; Sevilla, S.; Wiles, A.A.; et al. Adipocyte-derived exosomal miRNAs: A novel mechanism for obesity-related disease. Pediatr. Res. 2015, 77, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Huang-Doran, I.; Zhang, C.-Y.; Vidal-Puig, A. Extracellular Vesicles: Novel Mediators of Cell Communication In Metabolic Disease. Trends Endocrinol. Metab. 2017, 28, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Storti, G.; Scioli, M.G.; Kim, B.-S.; Terriaca, S.; Fiorelli, E.; Orlandi, A.; Cervelli, V. Mesenchymal Stem Cells in Adipose Tissue and Extracellular Vesicles in Ovarian Cancer Patients: A Bridge toward Metastatic Diffusion or a New Therapeutic Opportunity? Cells 2021, 10, 2117. [Google Scholar] [CrossRef]
- Röszer, T. Metabolic impact of adipose tissue macrophages in the early postnatal life. J. Leukoc. Biol. 2022, 112, 1515–1524. [Google Scholar] [CrossRef]
- McMorrow, A.M.; Connaughton, R.M.; Lithander, F.E.; Roche, H.M. Adipose tissue dysregulation and metabolic consequences in childhood and adolescent obesity: Potential impact of dietary fat quality. Proc. Nutr. Soc. 2015, 74, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M. Metabolically Healthy Obesity. Endocr. Rev. 2020, 41, bnaa004. [Google Scholar] [CrossRef]
- Carobbio, S.; Pellegrinelli, V.; Vidal-Puig, A. Adipose Tissue Function and Expandability as Determinants of Lipotoxicity and the Metabolic Syndrome. Adv. Exp. Med. Biol. 2017, 960, 161–196. [Google Scholar] [CrossRef]
- Rydén, M.; Andersson, D.P.; Bergström, I.B.; Arner, P. Adipose tissue and metabolic alterations: Regional differences in fat cell size and number matter, but differently: A cross-sectional study. J. Clin. Endocrinol. Metab. 2014, 99, E1870–E1876. [Google Scholar] [CrossRef] [PubMed]
- Lacasa, D.; Taleb, S.; Keophiphath, M.; Miranville, A.; Clement, K. Macrophage-Secreted Factors Impair Human Adipogenesis: Involvement of Proinflammatory State in Preadipocytes. Endocrinology 2007, 148, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Kusminski, C.M.; Scherer, P.E. Adipose tissue remodeling and obesity. J. Clin. Investig. 2011, 121, 2094–2101. [Google Scholar] [CrossRef] [PubMed]
- Iacobini, C.; Pugliese, G.; Blasetti Fantauzzi, C.; Federici, M.; Menini, S. Metabolically healthy versus metabolically unhealthy obesity. Metabolism 2019, 92, 51–60. [Google Scholar] [CrossRef]
- Yaghootkar, H.; Lotta, L.A.; Tyrrell, J.; Smit, R.A.; Jones, S.E.; Donnelly, L.; Beaumont, R.; Campbell, A.; Tuke, M.A.; Hayward, C.; et al. Genetic Evidence for a Link between Favorable Adiposity and Lower Risk of Type 2 Diabetes, Hypertension, and Heart Disease. Diabetes 2016, 65, 2448–2460. [Google Scholar] [CrossRef] [PubMed]
- Ou, M.-Y.; Zhang, H.; Tan, P.-C.; Zhou, S.-B.; Li, Q.-F. Adipose tissue aging: Mechanisms and therapeutic implications. Cell Death Dis. 2022, 13, 300. [Google Scholar] [CrossRef] [PubMed]
- Fitzgibbons, T.P.; Kogan, S.; Aouadi, M.; Hendricks, G.M.; Straubhaar, J.; Czech, M.P. Similarity of mouse perivascular and brown adipose tissues and their resistance to diet-induced inflammation. Am. J. Physiol.-Heart Circ. Physiol. 2011, 301, H1425–H1437. [Google Scholar] [CrossRef]
- Sakamoto, T. Inflammation induced by RAW macrophages suppresses UCP1 mRNA induction via ERK activation in 10T1/2 adipocytes. Am. J. Physiol. Cell. Physiol. 2013, 304, 729–738. [Google Scholar] [CrossRef]
- Pellegrinelli, V.; Heuvingh, J.; du Roure, O.; Rouault, C.; Devulder, A.; Klein, C.; Lacasa, M.; Clément, E.; Lacasa, D.; Clément, K. Human adipocyte function is impacted by mechanical cues. J. Pathol. 2014, 233, 183–195. [Google Scholar] [CrossRef]
- Abdennour, M.; Reggio, S.; Le Naour, G.; Liu, Y.; Poitou, C.; Aron-Wisnewsky, J.; Charlotte, F.; Bouillot, J.-L.; Torcivia, A.; Sasso, M.; et al. Association of Adipose Tissue and Liver Fibrosis with Tissue Stiffness in Morbid Obesity: Links with Diabetes and BMI Loss After Gastric Bypass. J. Clin. Endocrinol. Metab. 2014, 99, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Tchoukalova, Y.; Koutsari, C.; Jensen, M. Committed subcutaneous preadipocytes are reduced in human obesity. Diabetologia 2007, 50, 151–157. [Google Scholar] [CrossRef]
- Haque, W.A.; Oral, E.A.; Dietz, K.; Bowcock, A.M.; Agarwal, A.K.; Garg, A. Risk Factors for Diabetes in Familial Partial Lipodystrophy, Dunnigan Variety. Diabetes Care 2003, 26, 1350–1355. [Google Scholar] [CrossRef] [PubMed]
- Wong, V.W.-S.; Wong, G.L.-H.; Yeung, D.K.-W.; Abrigo, J.M.; Kong, A.P.-S.; Chan, R.S.-M.; Chim, A.M.-L.; Shen, J.; Ho, C.-S.; Woo, J.; et al. Fatty Pancreas, Insulin Resistance, and β-Cell Function: A Population Study Using Fat-Water Magnetic Resonance Imaging. Am. J. Gastroenterol. 2014, 109, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.J.; Valencia, A.; Startzell, M.; Cochran, E.; Walter, P.J.; Garraffo, H.M.; Cai, H.; Gharib, A.M.; Ouwerkerk, R.; Courville, A.B.; et al. Metreleptin-mediated improvements in insulin sensitivity are independent of food intake in humans with lipodystrophy. J. Clin. Investig. 2018, 128, 3504–3516. [Google Scholar] [CrossRef] [PubMed]
- Fischer-Posovszky, P.; Hebestreit, H.; Hofmann, A.K.; Strauss, G.; Möller, P.; Debatin, K.-M.; Wabitsch, M. Role of CD95-Mediated Adipocyte Loss in Autoimmune Lipodystrophy. J. Clin. Endocrinol. Metab. 2006, 91, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Torriani, M.; Srinivasa, S.; Fitch, K.V.; Thomou, T.; Wong, K.; Petrow, E.; Kahn, C.R.; Cypess, A.M.; Grinspoon, S.K. Dysfunctional Subcutaneous Fat with Reduced Dicer and Brown Adipose Tissue Gene Expression in HIV-Infected Patients. J. Clin. Endocrinol. Metab. 2016, 101, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic Adipose Tissue Inflammation Linking Obesity to Insulin Resistance and Type 2 Diabetes. Front. Physiol. 2020, 10, 1607. [Google Scholar] [CrossRef]
- Petersen, M.C.; Shulman, G.I. Mechanisms of Insulin Action and Insulin Resistance. Physiol. Rev. 2018, 98, 2133–2223. [Google Scholar] [CrossRef]
- Lin, Y.; Bai, M.; Wang, S.; Chen, L.; Li, Z.; Li, C.; Cao, P.; Chen, Y. Lactate Is a Key Mediator That Links Obesity to Insulin Resistance via Modulating Cytokine Production from Adipose Tissue. Diabetes 2022, 71, 637–652. [Google Scholar] [CrossRef]
- Wang, J.; Li, L.; Zhang, Z.; Zhang, X.; Zhu, Y.; Zhang, C.; Bi, Y. Extracellular vesicles mediate the communication of adipose tissue with brain and promote cognitive impairment associated with insulin resistance. Cell Metab. 2022, 34, 1264–1279.e8. [Google Scholar] [CrossRef]
- Pellegrinelli, V.; Rodriguez-Cuenca, S.; Rouault, C.; Figueroa-Juarez, E.; Schilbert, H.; Virtue, S.; Moreno-Navarrete, J.M.; Bidault, G.; Vázquez-Borrego, M.C.; Dias, A.R.; et al. Dysregulation of macrophage PEPD in obesity determines adipose tissue fibro-inflammation and insulin resistance. Nat. Metab. 2022, 4, 476–494. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Sun, W.; Shen, Y.; Baláz, M.; Balázová, L.; Ding, L.; Löffler, M.; Hamilton, B.; Klöting, N.; Blüher, M.; et al. Identification of a regulatory pathway inhibiting adipogenesis via RSPO2. Nat. Metab. 2022, 4, 90–105. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.G.; Choi, W.; Oh, T.J.; Cha, H.-N.; Hwang, I.; Lee, Y.K.; Lee, S.Y.; Shin, H.; Lim, A.; Ryu, D.; et al. Inhibiting serotonin signaling through HTR2B in visceral adipose tissue improves obesity-related insulin resistance. J. Clin. Investig. 2021, 131, e145331. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, B.; Gasek, N.S.; Zhou, Y.; Cohn, R.L.; Martin, D.E.; Zuo, W.; Flynn, W.F.; Guo, C.; Jellison, E.R.; et al. Targeting p21Cip1 highly expressing cells in adipose tissue alleviates insulin resistance in obesity. Cell Metab. 2022, 34, 75–89.e8. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Morreale, H.F. Polycystic ovary syndrome: Definition, aetiology, diagnosis and treatment. Nat. Rev. Endocrinol. 2018, 14, 270–284. [Google Scholar] [CrossRef] [PubMed]
- Rudnicka, E.; Suchta, K.; Grymowicz, M.; Calik-Ksepka, A.; Smolarczyk, K.; Duszewska, A.M.; Smolarczyk, R.; Meczekalski, B. Chronic Low Grade Inflammation in Pathogenesis of PCOS. Int. J. Mol. Sci. 2021, 22, 3789. [Google Scholar] [CrossRef]
- Silvestris, E.; de Pergola, G.; Rosania, R.; Loverro, G. Obesity as disruptor of the female fertility. Reprod. Biol. Endocrinol. 2018, 16, 22. [Google Scholar] [CrossRef] [PubMed]
- Psilopanagioti, A.; Papadaki, H.; Kranioti, E.F.; Alexandrides, T.K.; Varakis, J.N. Expression of Adiponectin and Adiponectin Receptors in Human Pituitary Gland and Brain. Neuroendocrinology 2009, 89, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-H.; Lee, K.; Moon, Y.S.; Sul, H.S. A Cysteine-rich Adipose Tissue-specific Secretory Factor Inhibits Adipocyte Differentiation. J. Biol. Chem. 2001, 276, 11252–11256. [Google Scholar] [CrossRef]
- Spicer, L.J.; Schreiber, N.B.; Lagaly, D.V.; Aad, P.Y.; Douthit, L.B.; Grado-Ahuir, J.A. Effect of resistin on granulosa and theca cell function in cattle. Anim. Reprod. Sci. 2011, 124, 19–27. [Google Scholar] [CrossRef]
- Escobar-Morreale, H.F.; Luque-Ramírez, M.; González, F. Circulating inflammatory markers in polycystic ovary syndrome: A systematic review and metaanalysis. Fertil. Steril. 2011, 95, 1048–1058.e2. [Google Scholar] [CrossRef]
- González, F.; Considine, R.V.; Abdelhadi, O.A.; Acton, A.J. Inflammation Triggered by Saturated Fat Ingestion Is Linked to Insulin Resistance and Hyperandrogenism in Polycystic Ovary Syndrome. J. Clin. Endocrinol. Metab. 2020, 105, E2152–E2167. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, R.L.; Ryan, K.J. Hyperandrogenism, insulin resistance, and acanthosis nigricans syndrome: A common endocrinopathy with distinct pathophysiologic features. Am. J. Obstet. Gynecol. 1983, 147, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, K.; Mathur, R.; Purwar, N.; Mathur, S.K.; Mathur, D.K. Hyperandrogenism, Insulin Resistance, and Acanthosis Nigricans (HAIR-AN) Syndrome Reflects Adipose Tissue Dysfunction (“Adiposopathy” or “Sick Fat”) in Asian Indian Girls. Dermatology 2021, 237, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Gołacki, J.; Matuszek, M.; Matyjaszek-Matuszek, B. Link between Insulin Resistance and Obesity—From Diagnosis to Treatment. Diagnostics 2022, 12, 1681. [Google Scholar] [CrossRef] [PubMed]
- Gómez, A.P.; Memije, M.V.; Tamayo, M.T.; Carreón, A.R. Skin disorders in overweight and obese patients and their relationship with insulin. Actas Dermosifiliogr. 2014, 105, 178–185. [Google Scholar] [CrossRef]
- Kutbay, N. An Unusual Case of Acquired Partial Lipodystrophy Presenting with Acanthosis Nigricans. Acta Endocrinol. 2019, 15, 129–130. [Google Scholar] [CrossRef] [PubMed]
- Verdecchia, F.; Akcan, N.; Dastamani, A.; Morgan, K.; Semple, R.K.; Shah, P. Unusual Glycemic Presentations in a Child with a Novel Heterozygous Intragenic INSR Deletion. Horm. Res. Paediatr. 2020, 93, 396–401. [Google Scholar] [CrossRef]
- Neeland, I.J.; Ross, R.; Després, J.-P.; Matsuzawa, Y.; Yamashita, S.; Shai, I.; Seidell, J.; Magni, P.; Santos, R.D.; Arsenault, B.; et al. Visceral and ectopic fat, atherosclerosis, and cardiometabolic disease: A position statement. Lancet Diabetes Endocrinol. 2019, 7, 715–725. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Wang, P.; Zhang, S.-Y.; Dong, Y.; Zeng, G.; Yan, Y.; Sun, L.; Wu, Q.; Liu, H.; et al. Adipocyte Hypoxia-Inducible Factor 2α Suppresses Atherosclerosis by Promoting Adipose Ceramide Catabolism. Cell Metab. 2019, 30, 937–951.e5. [Google Scholar] [CrossRef]
- Hu, H.; Garcia-Barrio, M.; Jiang, Z.-S.; Chen, Y.E.; Chang, L. Roles of Perivascular Adipose Tissue in Hypertension and Atherosclerosis. Antioxid. Redox Signal. 2021, 34, 736–749. [Google Scholar] [CrossRef]
- Mancio, J.; Oikonomou, E.K.; Antoniades, C. Perivascular adipose tissue and coronary atherosclerosis. Heart 2018, 104, 1654–1662. [Google Scholar] [CrossRef] [PubMed]
- Adachi, Y.; Ueda, K.; Nomura, S.; Ito, K.; Katoh, M.; Katagiri, M.; Yamada, S.; Hashimoto, M.; Zhai, B.; Numata, G.; et al. Beiging of perivascular adipose tissue regulates its inflammation and vascular remodeling. Nat. Commun. 2022, 13, 5117. [Google Scholar] [CrossRef] [PubMed]
- Mu, W.; Qian, S.; Song, Y.; Yang, L.; Song, S.; Yang, Q.; Liu, H.; Liu, Y.; Pan, D.; Tang, Y.; et al. BMP4-mediated browning of perivascular adipose tissue governs an anti-inflammatory program and prevents atherosclerosis. Redox Biol. 2021, 43, 101979. [Google Scholar] [CrossRef]
- Eslam, M.; Newsome, P.N.; Sarin, S.K.; Anstee, Q.M.; Targher, G.; Romero-Gomez, M.; Zelber-Sagi, S.; Wong, V.W.-S.; Dufour, J.-F.; Schattenberg, J.M.; et al. A new definition for metabolic dysfunction-associated fatty liver disease: An international expert consensus statement. J. Hepatol. 2020, 73, 202–209. [Google Scholar] [CrossRef]
- Alharthi, J.; Latchoumanin, O.; George, J.; Eslam, M. Macrophages in metabolic associated fatty liver disease. World J. Gastroenterol. 2020, 26, 1861–1878. [Google Scholar] [CrossRef] [PubMed]
- Kazankov, K.; Jørgensen, S.M.D.; Thomsen, K.L.; Møller, H.J.; Vilstrup, H.; George, J.; Schuppan, D.; Grønbæk, H. The role of macrophages in nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 145–159. [Google Scholar] [CrossRef]
- Fukushima, J.; Kamada, Y.; Matsumoto, H.; Yoshida, Y.; Ezaki, H.; Takemura, T.; Saji, Y.; Igura, T.; Tsutsui, S.; Kihara, S.; et al. Adiponectin prevents progression of steatohepatitis in mice by regulating oxidative stress and Kupffer cell phenotype polarization. Hepatol. Res. 2009, 39, 724–738. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Leclercq, I.; Brymora, J.M.; Xu, N.; Ramezani–Moghadam, M.; London, R.M.; Brigstock, D.; George, J. Kupffer Cells Mediate Leptin-Induced Liver Fibrosis. Gastroenterology 2009, 137, 713–723.e1. [Google Scholar] [CrossRef] [PubMed]
- Stanton, M.C.; Chen, S.-C.; Jackson, J.V.; Rojas-Triana, A.; Kinsley, D.; Cui, L.; Fine, J.S.; Greenfeder, S.; Bober, L.A.; Jenh, C.-H. Inflammatory Signals shift from adipose to liver during high fat feeding and influence the development of steatohepatitis in mice. J. Inflamm. 2011, 8, 8. [Google Scholar] [CrossRef]
- Luo, W.; Xu, Q.; Wang, Q.; Wu, H.; Hua, J. Effect of modulation of PPAR-γ activity on Kupffer cells M1/M2 polarization in the development of non-alcoholic fatty liver disease. Sci. Rep. 2017, 7, srep44612. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, C.; Miserocchi, G.; De Vita, A.; Spadazzi, C.; Cocchi, C.; Vanni, S.; Gabellone, S.; Martinelli, G.; Ranallo, N.; Bongiovanni, A.; et al. Lipids and adipocytes involvement in tumor progression with a focus on obesity and diet. Obes. Rev. 2024, e13833, online ahead of print. [Google Scholar] [CrossRef]
- Nieman, K.M.; Kenny, H.A.; Penicka, C.V.; Ladanyi, A.; Buell-Gutbrod, R.; Zillhardt, M.R.; Romero, I.L.; Carey, M.S.; Mills, G.B.; Hotamisligil, G.S.; et al. Adipocytes promote ovarian cancer metastasis and provide energy for rapid tumor growth. Nat. Med. 2011, 17, 1498–1503. [Google Scholar] [CrossRef]
- Balaban, S.; Shearer, R.F.; Lee, L.S.; van Geldermalsen, M.; Schreuder, M.; Shtein, H.C.; Cairns, R.; Thomas, K.C.; Fazakerley, D.J.; Grewal, T.; et al. Adipocyte lipolysis links obesity to breast cancer growth: Adipocyte-derived fatty acids drive breast cancer cell proliferation and migration. Cancer Metab. 2017, 5, 1. [Google Scholar] [CrossRef]
- Zhang, M.; Di Martino, J.S.; Bowman, R.L.; Campbell, N.R.; Baksh, S.C.; Simon-Vermot, T.; Kim, I.S.; Haldeman, P.; Mondal, C.; Yong-Gonzales, V.; et al. Adipocyte-Derived Lipids Mediate Melanoma Progression via FATP Proteins. Cancer Discov. 2018, 8, 1006–1025. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Morley, T.S.; Kim, M.; Clegg, D.J.; Scherer, P.E. Obesity and cancer—Mechanisms underlying tumour progression and recurrence. Nat. Rev. Endocrinol. 2014, 10, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y. Angiogenesis and vascular functions in modulation of obesity, adipose metabolism, and insulin sensitivity. Cell Metab. 2013, 18, 478–489. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-H.; Mottillo, E.P.; Granneman, J.G. Adipose tissue plasticity from WAT to BAT and in between. Biochim. Biophys. Acta 2014, 1842, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Qiang, L.; Wang, L.; Kon, N.; Zhao, W.; Lee, S.; Zhang, Y.; Rosenbaum, M.; Zhao, Y.; Gu, W.; Farmer, S.R.; et al. Brown Remodeling of White Adipose Tissue by SirT1-Dependent Deacetylation of Ppary. Cell 2012, 150, 620–632. [Google Scholar] [CrossRef]
- Ohno, H.; Shinoda, K.; Spiegelman, B.M.; Kajimura, S. PPARγ agonists Induce a White-to-Brown Fat Conversion through Stabilization of PRDM16 Protein. Cell Metab. 2012, 15, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Lai, X.-R.; Lu, J.-Q.; Tang, L.-Z.; Zhang, J.-R.; Liu, H.-W. Decellularized adipose tissue: A key factor in promoting fat regeneration by recruiting and inducing mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2021, 541, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Cypess, A.M.; Lehman, S.; Williams, G.; Tal, I.; Rodman, D.; Goldfine, A.B.; Kuo, F.C.; Palmer, E.L.; Tseng, Y.-H.; Doria, A.; et al. Identification and Importance of Brown Adipose Tissue in Adult Humans. N. Engl. J. Med. 2009, 360, 1509–1517. [Google Scholar] [CrossRef]
- Liu, X.; Zheng, Z.; Zhu, X.; Meng, M.; Li, L.; Shen, Y.; Chi, Q.; Wang, D.; Zhang, Z.; Li, C.; et al. Brown adipose tissue transplantation improves whole-body energy metabolism. Cell Res. 2013, 23, 851–854. [Google Scholar] [CrossRef] [PubMed]
- Stanford, K.I.; Middelbeek, R.J.; Townsend, K.L.; An, D.; Nygaard, E.B.; Hitchcox, K.M.; Markan, K.R.; Nakano, K.; Hirshman, M.F.; Tseng, Y.-H.; et al. Brown adipose tissue regulates glucose homeostasis and insulin sensitivity. J. Clin. Investig. 2013, 123, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, H.; Li, B.; Meng, X.; Wang, J.; Zhang, Y.; Yao, S.; Ma, Q.; Jin, L.; Yang, J.; et al. Berberine activates thermogenesis in white and brown adipose tissue. Nat. Commun. 2014, 5, 5493. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, J.; He, X.; Sheng, Y.; Yang, C.; Li, H.; Xu, J.; Xu, W.; Huang, K. Caulis Spatholobi Ameliorates Obesity through Activating Brown Adipose Tissue and Modulating the Composition of Gut Microbiota. Int. J. Mol. Sci. 2019, 20, 5150. [Google Scholar] [CrossRef]
- Quan, L.-H.; Zhang, C.; Dong, M.; Jiang, J.; Xu, H.; Yan, C.; Liu, X.; Zhou, H.; Zhang, H.; Chen, L.; et al. Myristoleic acid produced by enterococci reduces obesity through brown adipose tissue activation. Gut 2020, 69, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Cheong, L.Y.; Xu, A. Intercellular and inter-organ crosstalk in browning of white adipose tissue: Molecular mechanism and therapeutic complications. J. Mol. Cell Biol. 2021, 13, 466–479. [Google Scholar] [CrossRef]
- Singh, A.K.; Aryal, B.; Chaube, B.; Rotllan, N.; Varela, L.; Horvath, T.L.; Suárez, Y.; Fernández-Hernando, C. Brown adipose tissue derived ANGPTL4 controls glucose and lipid metabolism and regulates thermogenesis. Mol. Metab. 2018, 11, 59–69. [Google Scholar] [CrossRef]
- Giordano, A.; Frontini, A.; Cinti, S. Convertible visceral fat as a therapeutic target to curb obesity. Nat. Rev. Drug Discov. 2016, 15, 405–424. [Google Scholar] [CrossRef]
- Liao, W.; Yin, X.; Li, Q.; Zhang, H.; Liu, Z.; Zheng, X.; Zheng, L.; Feng, X. Resveratrol-Induced White Adipose Tissue Browning in Obese Mice by Remodeling Fecal Microbiota. Molecules 2018, 23, 3356. [Google Scholar] [CrossRef] [PubMed]
- Mader, I.; Wabitsch, M.; Debatin, K.; Fischer-Posovszky, P.; Fulda, S. Identification of a novel proapoptotic function of resveratrol in fat cells: SIRT1-independent sensitization to TRAIL-induced apoptosis. FASEB J. 2010, 24, 1997–2009. [Google Scholar] [CrossRef] [PubMed]
- The Key Role of a Glucagon-Like Peptide-1 Receptor Agonist in Body Fat Redistribution—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/30530905/ (accessed on 9 October 2022).
- Zhu, D.; Li, X.; Ma, J.; Zeng, J.; Gan, S.; Dong, X.; Yang, J.; Lin, X.; Cai, H.; Song, W.; et al. Dorzagliatin in drug-naïve patients with type 2 diabetes: A randomized, double-blind, placebo-controlled phase 3 trial. Nat. Med. 2022, 28, 965–973. [Google Scholar] [CrossRef]
- Seoane-Collazo, P.; Liñares-Pose, L.; Rial-Pensado, E.; Romero-Picó, A.; Moreno-Navarrete, J.M.; Martínez-Sánchez, N.; Garrido-Gil, P.; Iglesias-Rey, R.; Morgan, D.A.; Tomasini, N.; et al. Central nicotine induces browning through hypothalamic κ opioid receptor. Nat. Commun. 2019, 10, 4037. [Google Scholar] [CrossRef]
- Bonet, M.L.; Ribot, J.; Palou, A. Lipid metabolism in mammalian tissues and its control by retinoic acid. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2012, 1821, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Sakellariou, P.; Valente, A.; Carrillo, A.E.; Metsios, G.S.; Nadolnik, L.; Jamurtas, A.Z.; Koutedakis, Y.; Boguszewski, C.; Andrade, C.M.B.; Svensson, P.-A.; et al. Chronic l-menthol-induced browning of white adipose tissue hypothesis: A putative therapeutic regime for combating obesity and improving metabolic health. Med. Hypotheses 2016, 93, 21–26. [Google Scholar] [CrossRef]
- Lee, C.G.; Rhee, D.K.; Kim, B.O.; Um, S.H.; Pyo, S. Allicin induces beige-like adipocytes via KLF15 signal cascade. J. Nutr. Biochem. 2019, 64, 13–24. [Google Scholar] [CrossRef]
- Han, Y.; Wu, J.-Z.; Shen, J.-Z.; Chen, L.; He, T.; Jin, M.-W.; Liu, H. Pentamethylquercetin induces adipose browning and exerts beneficial effects in 3T3-L1 adipocytes and high-fat diet-fed mice. Sci. Rep. 2017, 7, 1123. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Lundh, M.; Fu, A.; Kriszt, R.; Huang, T.L.; Lynes, M.D.; Leiria, L.O.; Shamsi, F.; Darcy, J.; Greenwood, B.P.; et al. CRISPR-engineered human brown-like adipocytes prevent diet-induced obesity and ameliorate metabolic syn-drome in mice. Sci. Transl. Med. 2020, 12, eaaz8664. [Google Scholar] [CrossRef] [PubMed]
- Herold, C.; Rennekampff, H.O.; Engeli, S. Apoptotic pathways in adipose tissue. Apoptosis 2013, 18, 911–916. [Google Scholar] [CrossRef]
- Lee, M.S.; Kim, C.T.; Kim, Y. Green tea (-)-epigallocatechin-3-gallate reduces body weight with regulation of multiple genes expres-sion in adipose tissue of diet-induced obese mice. Ann. Nutr. Metab. 2009, 54, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Grigem, S.; Fischer-Posovszky, P.; Debatin, K.M.; Loizon, E.; Vidal, H.; Wabitsch, M. The effect of the HIV protease inhibitor ritonavir on proliferation, differentiation, lipogenesis, gene expression and apoptosis of human preadipocytes and adipocytes. Horm. Metab. Res. 2005, 37, 602–609. [Google Scholar] [CrossRef]
- Liao, Z.-Z.; Ran, L.; Qi, X.-Y.; Wang, Y.-D.; Wang, Y.-Y.; Yang, J.; Liu, J.-H.; Xiao, X.-H. Adipose endothelial cells mastering adipose tissues metabolic fate. Adipocyte 2022, 11, 108–119. [Google Scholar] [CrossRef]
- Xian, Y.; Chen, Z.; Deng, H.; Cai, M.; Liang, H.; Xu, W.; Weng, J.; Xu, F. Exenatide mitigates inflammation and hypoxia along with improved angiogenesis in obese fat tissue. J. Endocrinol. 2019, 242, 79–89. [Google Scholar] [CrossRef]
- Kito, T.; Shibata, R.; Kondo, M.; Yamamoto, T.; Suzuki, H.; Ishii, M.; Murohara, T. Nifedipine ameliorates ischemia-induced revascularization in diet-induced obese mice. Am. J. Hypertens. 2012, 25, 401–406. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, G.; Lin, F.; Wan, Q.; Wu, J.; Luo, M. Mechanisms of action of metformin and its regulatory effect on microRNAs related to angiogenesis. Pharmacol. Res. 2021, 164, 105390. [Google Scholar] [CrossRef] [PubMed]
- Duerre, D.J.; Galmozzi, A. Deconstructing Adipose Tissue Heterogeneity One Cell at a Time. Front. Endocrinol. 2022, 13, 847291. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Liang, S.; Guo, C.; Yu, X.; Zhao, J.; Zhang, H.; Shang, W. Inhibition of M1 macrophage activation in adipose tissue by berberine improves insulin resistance. Life Sci. 2016, 166, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Stanley, T.L.; Zanni, M.V.; Johnsen, S.; Rasheed, S.; Makimura, H.; Lee, H.; Khor, V.K.; Ahima, R.S.; Grinspoon, S.K. TNF-α Antagonism with Etanercept Decreases Glucose and Increases the Proportion of High Molecular Weight Adiponectin in Obese Subjects with Features of the Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2011, 96, E146–E150. [Google Scholar] [CrossRef]
- Braun, K.; Oeckl, J.; Westermeier, J.; Li, Y.; Klingenspor, M. Non-adrenergic control of lipolysis and thermogenesis in adipose tissues. J. Exp. Biol. 2018, 221 (Suppl. S1), jeb165381. [Google Scholar] [CrossRef] [PubMed]
- Orsso, C.E.; Colin-Ramirez, E.; Field, C.J.; Madsen, K.L.; Prado, C.M.; Haqq, A.M. Adipose Tissue Development and Expansion from the Womb to Adolescence: An Overview. Nutrients 2020, 12, 2735. [Google Scholar] [CrossRef]
- Chan, M.; Lim, Y.C.; Yang, J.; Namwanje, M.; Liu, L.; Qiang, L. Identification of a natural beige adipose depot in mice. J. Biol. Chem. 2019, 294, 6751–6761. [Google Scholar] [CrossRef] [PubMed]
- Cinti, S. Pink Adipocytes. Trends Endocrinol. Metab. 2018, 29, 651–666. [Google Scholar] [CrossRef] [PubMed]
- Prokesch, A.; Smorlesi, A.; Perugini, J.; Manieri, M.; Ciarmela, P.; Mondini, E.; Trajanoski, Z.; Kristiansen, K.; Giordano, A.; Bogner-Strauss, J.; et al. Molecular Aspects of Adipoepithelial Transdifferentiation in Mouse Mammary Gland. Stem Cells 2014, 32, 2756–2766. [Google Scholar] [CrossRef]
- Giordano, A.; Perugini, J.; Kristensen, D.M.; Sartini, L.; Frontini, A.; Kajimura, S.; Kristiansen, K.; Cinti, S. Mammary alveolar epithelial cells convert to brown adipocytes in post-lactating mice. J. Cell. Physiol. 2017, 232, 2923–2928. [Google Scholar] [CrossRef] [PubMed]
- Ricquier, D. UCP1, the mitochondrial uncoupling protein of brown adipocyte: A personal contribution and a historical perspective. Biochimie 2017, 134, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Sebo, Z.L.; Jeffery, E.; Holtrup, B.; Rodeheffer, M.S. A mesodermal fate map for adipose tissue. Development 2018, 145, dev.166801. [Google Scholar] [CrossRef] [PubMed]
- Gesta, S.; Tseng, Y.-H.; Kahn, C.R. Developmental Origin of Fat: Tracking Obesity to Its Source. Cell 2007, 131, 242–256. [Google Scholar] [CrossRef]
- Sanchez-Gurmaches, J.; Guertin, D.A. Adipocytes arise from multiple lineages that are heterogeneously and dynamically distributed. Nat. Commun. 2014, 5, 4099. [Google Scholar] [CrossRef] [PubMed]
- Eirin, A.; Thaler, R.; Glasstetter, L.M.; Xing, L.; Zhu, X.-Y.; Osborne, A.C.; Mondesir, R.; Bhagwate, A.V.; Lerman, A.; van Wijnen, A.J.; et al. Obesity-driven mitochondrial dysfunction in human adipose tissue-derived mesenchymal stem/stromal cells involves epigenetic changes. Cell Death Dis. 2024, 15, 387. [Google Scholar] [CrossRef] [PubMed]
- Schwalie, P.C.; Dong, H.; Zachara, M.; Russeil, J.; Alpern, D.; Akchiche, N.; Caprara, C.; Sun, W.; Schlaudraff, K.-U.; Soldati, G.; et al. A stromal cell population that inhibits adipogenesis in mammalian fat depots. Nature 2018, 559, 103–108. [Google Scholar] [CrossRef]
- Marcelin, G.; Silveira, A.L.M.; Martins, L.B.; Ferreira, A.V.; Clément, K. Deciphering the cellular interplays underlying obesity-induced adipose tissue fibrosis. J. Clin. Investig. 2019, 129, 4032–4040. [Google Scholar] [CrossRef] [PubMed]
- Bäckdahl, J.; Franzén, L.; Massier, L.; Li, Q.; Jalkanen, J.; Gao, H.; Andersson, A.; Bhalla, N.; Thorell, A.; Rydén, M.; et al. Spatial mapping reveals human adipocyte subpopulations with distinct sensitivities to insulin. Cell Metab. 2021, 33, 1869–1882.e6. [Google Scholar] [CrossRef] [PubMed]
- Rogal, J.; Roosz, J.; Teufel, C.; Cipriano, M.; Xu, R.; Eisler, W.; Weiss, M.; Schenke-Layland, K.; Loskill, P. Autologous Human Immunocompetent White Adipose Tissue-on-Chip. Adv. Sci. 2022, 9, e2104451. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).