

Transgenic Overexpression of HDAC9 Promotes Adipocyte Hypertrophy, Insulin Resistance and Hepatic Steatosis in Aging Mice

 , , , , ,
, , , , ,  ,
, 

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. HDAC9 TG Mice
2.2. Genotyping
2.3. RT-PCR
2.4. Western Blot
2.5. Plasma Insulin Measurement
2.6. Body Composition Measurements
2.7. Insulin Tolerance Test
2.8. Glucose Tolerance Test
2.9. Histology
2.10. Noninvasive Blood Pressure Measurement
2.11. Pulse Wave Velocity
2.12. Statistics
3. Results
3.1. Generation and Validation of HDAC9 TG Mice
3.2. General Features of HDAC9 TG Mice
3.3. Impaired Insulin Sensitivity and Glucose Tolerance in Aging HDAC9 TG Mice
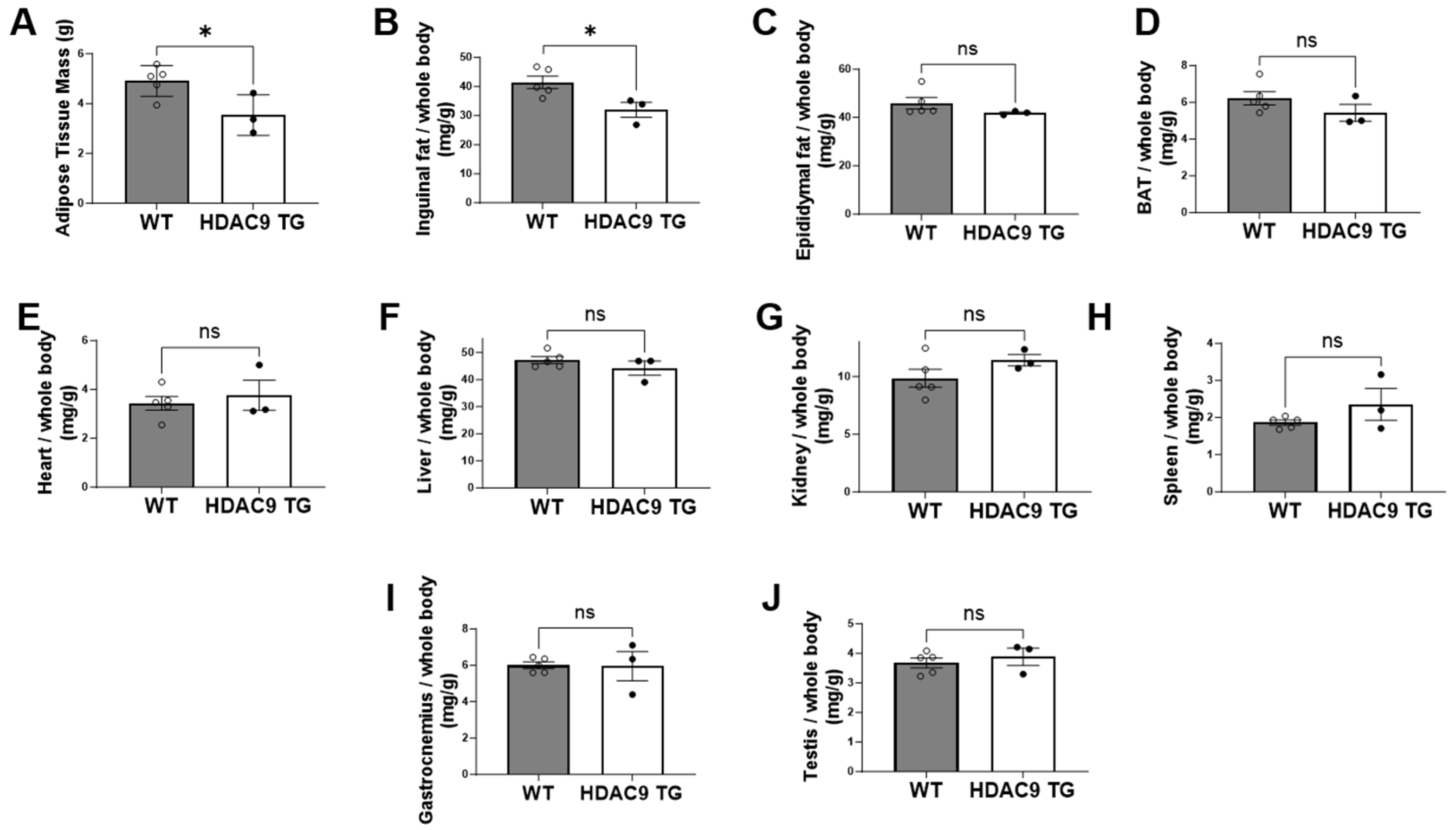
3.4. Reduced Inguinal Adipose Tissue in HDAC9 TG Mice
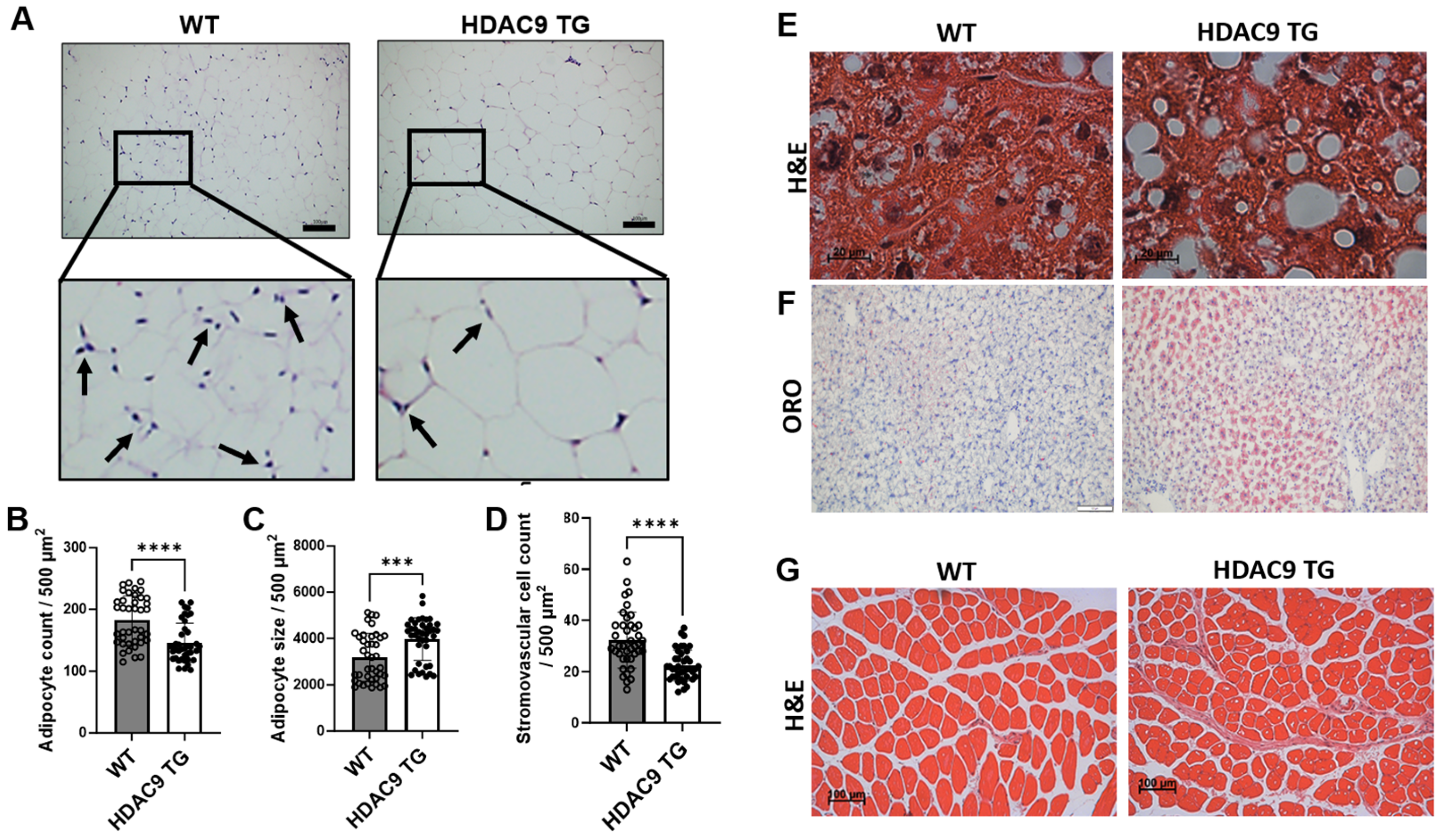
3.5. Morphological Features of Adipose Tissue, Liver and Skeletal Muscle in HDAC9 TG Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Haberland, M.; Montgomery, R.L.; Olson, E.N. The many roles of histone deacetylases in development and physiology: Implications for disease and therapy. Nat. Rev. Genet. 2009, 10, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Parra, M. Class IIa HDACs—New insights into their functions in physiology and pathology. FEBS J. 2015, 282, 1736–1744. [Google Scholar] [CrossRef] [PubMed]
- Brancolini, C.; Di Giorgio, E.; Formisano, L.; Gagliano, T. Quis Custodiet Ipsos Custodes (Who Controls the Controllers)? Two decades of studies on HDAC9. Life 2021, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, R.A.; Weeks, K.L. Histone deacetylases in cardiovascular and metabolic diseases. J. Mol. Cell. Cardiol. 2019, 130, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Croteau, S.; Hardy, P. Histone deacetylase (HDAC) 9: Versatile biological functions and emerging roles in human cancer. Cell. Oncol. 2021, 44, 997–1017. [Google Scholar] [CrossRef] [PubMed]
- White, U. Adipose tissue expansion in obesity, health, and disease. Front. Cell Dev. Biol. 2023, 11, 1188844. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding adipocyte differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [CrossRef] [PubMed]
- Isakson, P.; Hammarstedt, A.; Gustafson, B.; Smith, U. Impaired preadipocyte differentiation in human abdominal obesity: Role of Wnt, tumor necrosis factor-alpha, and inflammation. Diabetes 2009, 58, 1550–1557. [Google Scholar] [CrossRef] [PubMed]
- Laforest, S.; Labrecque, J.; Michaud, A.; Cianflone, K.; Tchernof, A. Adipocyte size as a determinant of metabolic disease and adipose tissue dysfunction. Crit. Rev. Clin. Lab. Sci. 2015, 52, 301–313. [Google Scholar] [CrossRef]
- Chatterjee, T.K.; Idelman, G.; Blanco, V.; Blomkalns, A.L.; Piegore, M.G., Jr.; Weintraub, D.S.; Kumar, S.; Rajsheker, S.; Manka, D.; Rudich, S.M.; et al. Histone deacetylase 9 is a negative regulator of adipogenic differentiation. J. Biol. Chem. 2011, 286, 27836–27847. [Google Scholar] [CrossRef]
- Chatterjee, T.K.; Basford, J.E.; Knoll, E.; Tong, W.S.; Blanco, V.; Blomkalns, A.L.; Rudich, S.; Lentsch, A.B.; Hui, D.Y.; Weintraub, N.L. HDAC9 knockout mice are protected from adipose tissue dysfunction and systemic metabolic disease during high-fat feeding. Diabetes 2014, 63, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, N.; Dong, M.; Guo, M.; Zhao, Y.; Zhuo, Z.; Zhang, C.; Chi, X.; Pan, Y.; Jiang, J.; et al. The metabolic regulator histone deacetylase 9 contributes to glucose homeostasis abnormality induced by hepatitis C virus infection. Diabetes 2015, 64, 4088–4098. [Google Scholar] [CrossRef] [PubMed]
- Lenoir, O.; Flosseau, K.; Ma, F.X.; Blondeau, B.; Mai, A.; Bassel-Duby, R.; Ravassard, P.; Olson, E.N.; Haumaitre, C.; Scharfmann, R. Specific control of pancreatic endocrine beta- and delta-cell mass by class IIa histone deacetylases HDAC4, HDAC5, and HDAC9. Diabetes 2011, 60, 2861–2871. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Xu, T.; Wu, X. Generation of genetically engineered mice by the piggyBac transposon system. Methods Mol. Biol. 2014, 1194, 171–185. [Google Scholar] [CrossRef] [PubMed]
- Goo, B.; Ahmadieh, S.; Zarzour, A.; Yiew, N.K.H.; Kim, D.; Shi, H.; Greenway, J.; Cave, S.; Nguyen, J.; Aribindi, S.; et al. Sex-dependent role of adipose tissue HDAC9 in diet-induced obesity and metabolic dysfunction. Cells 2022, 11, 2698. [Google Scholar] [CrossRef] [PubMed]
- Yiew, N.K.H.; Greenway, C.; Zarzour, A.; Ahmadieh, S.; Goo, B.; Kim, D.; Benson, T.W.; Ogbi, M.; Tang, Y.L.; Chen, W.; et al. Enhancer of zeste homolog 2 (EZH2) regulates adipocyte lipid metabolism independent of adipogenic differentiation: Role of apolipoprotein E. J. Biol. Chem. 2019, 294, 8577–8591. [Google Scholar] [CrossRef] [PubMed]
- The International Stroke Genetics Consortium (ISGC); the Wellcome Trust Case Control Consortium 2 (WTCCC2); Bellenguez, C.; Bevan, S.; Gschwendtner, A.; Spencer, C.C.; Burgess, A.I.; Pirinen, M.; Jackson, C.A.; Traylor, M.; et al. Genome-wide association study identifies a variant in HDAC9 associated with large vessel ischemic stroke. Nat. Genet. 2012, 44, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Pande, S.V.; Parvin, R. Carnitine-acylcarnitine translocase-mediated transport of fatty acids into mitochondria: Its involvement in the control of fatty acid oxidation in liver. In Carnitine Biosynthesis, Metabolism, and Functions; Elsevier: Amsterdam, The Netherlands, 1980; Volume 61, pp. 43–157. [Google Scholar]
- Ahmadieh, S.; Goo, B.; Zarzour, A.; Kim, D.; Shi, H.; Veerapaneni, P.; Chouhaita, R.; Yiew, N.K.H.; Dominguez Gonzalez, C.; Chakravartty, A.; et al. Impact of housing temperature on adipose tissue HDAC9 expression and adipogenic differentiation in high fat-fed mice. Obesity 2023, 32, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Rong, S.; Repa, J.J.; St Clair, R.; Parks, J.S.; Mishra, N. Histone deacetylase 9 represses cholesterol efflux and alternatively activated macrophages in atherosclerosis development. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1871–1879. [Google Scholar] [CrossRef]
- Tchkonia, T.; Morbeck, D.E.; Von Zglinicki, T.; Van Deursen, J.; Lustgarten, J.; Scrable, H.; Khosla, S.; Jensen, M.D.; Kirkland, J.L. Fat tissue, aging, and cellular senescence. Aging Cell 2010, 9, 667–684. [Google Scholar] [CrossRef]
- Oñate, B.; Vilahur, G.; Ferrer-Lorente, R.; Ybarra, J.; Díez-Caballero, A.; Ballesta-López, C.; Moscatiello, F.; Herrero, J.; Badimon, L. The subcutaneous adipose tissue reservoir of functionally active stem cells is reduced in obese patients. FASEB J. 2012, 26, 4327–4336. [Google Scholar] [CrossRef] [PubMed]
- Rawshani, A.; Eliasson, B.; Rawshani, A.; Henninger, J.; Mardinoglu, A.; Carlsson, Å.; Sohlin, M.; Ljungberg, M.; Hammarstedt, A.; Rosengren, A.; et al. Adipose tissue morphology, imaging and metabolomics predicting cardiometabolic risk and family history of type 2 diabetes in non-obese men. Sci. Rep. 2020, 10, 9973. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-C.; Hsieh, P.-S. The role of adipocyte hypertrophy and hypoxia in the development of obesity-associated adipose tissue inflammation and insulin resistance. In Adiposity—Omics and Molecular Understanding; Gordeladze, J.O., Ed.; InTech: Rijeka, Croatia, 2017. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veerapaneni, P.; Goo, B.; Ahmadieh, S.; Shi, H.; Kim, D.S.; Ogbi, M.; Cave, S.; Chouhaita, R.; Cyriac, N.; Fulton, D.J.; et al. Transgenic Overexpression of HDAC9 Promotes Adipocyte Hypertrophy, Insulin Resistance and Hepatic Steatosis in Aging Mice. Biomolecules 2024, 14, 494. https://doi.org/10.3390/biom14040494
Veerapaneni P, Goo B, Ahmadieh S, Shi H, Kim DS, Ogbi M, Cave S, Chouhaita R, Cyriac N, Fulton DJ, et al. Transgenic Overexpression of HDAC9 Promotes Adipocyte Hypertrophy, Insulin Resistance and Hepatic Steatosis in Aging Mice. Biomolecules. 2024; 14(4):494. https://doi.org/10.3390/biom14040494
Chicago/Turabian StyleVeerapaneni, Praneet, Brandee Goo, Samah Ahmadieh, Hong Shi, David S. Kim, Mourad Ogbi, Stephen Cave, Ronnie Chouhaita, Nicole Cyriac, David J. Fulton, and et al. 2024. "Transgenic Overexpression of HDAC9 Promotes Adipocyte Hypertrophy, Insulin Resistance and Hepatic Steatosis in Aging Mice" Biomolecules 14, no. 4: 494. https://doi.org/10.3390/biom14040494
APA StyleVeerapaneni, P., Goo, B., Ahmadieh, S., Shi, H., Kim, D. S., Ogbi, M., Cave, S., Chouhaita, R., Cyriac, N., Fulton, D. J., Verin, A. D., Chen, W., Lei, Y., Lu, X.-Y., Kim, H. W., & Weintraub, N. L. (2024). Transgenic Overexpression of HDAC9 Promotes Adipocyte Hypertrophy, Insulin Resistance and Hepatic Steatosis in Aging Mice. Biomolecules, 14(4), 494. https://doi.org/10.3390/biom14040494