

Downregulation of miR-1388 Regulates the Expression of Antiviral Genes via Tumor Necrosis Factor Receptor (TNFR)-Associated Factor 3 Targeting Following poly(I:C) Stimulation in Silver Carp (Hypophthalmichthys molitrix)

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Silver Carp and Treatment
2.2. Cell Culture, Macrophage Isolation and poly(I:C) Exposure
2.3. Plasmid Construction and Transfection
2.4. miRNA Mimics and Inhibitors
2.5. RNA Interference
2.6. RNA Extraction and Real-Time Quantitative PCR
2.7. Dual-Luciferase Reporter Assays
2.8. Western Blotting
2.9. Statistical Analysis
3. Results
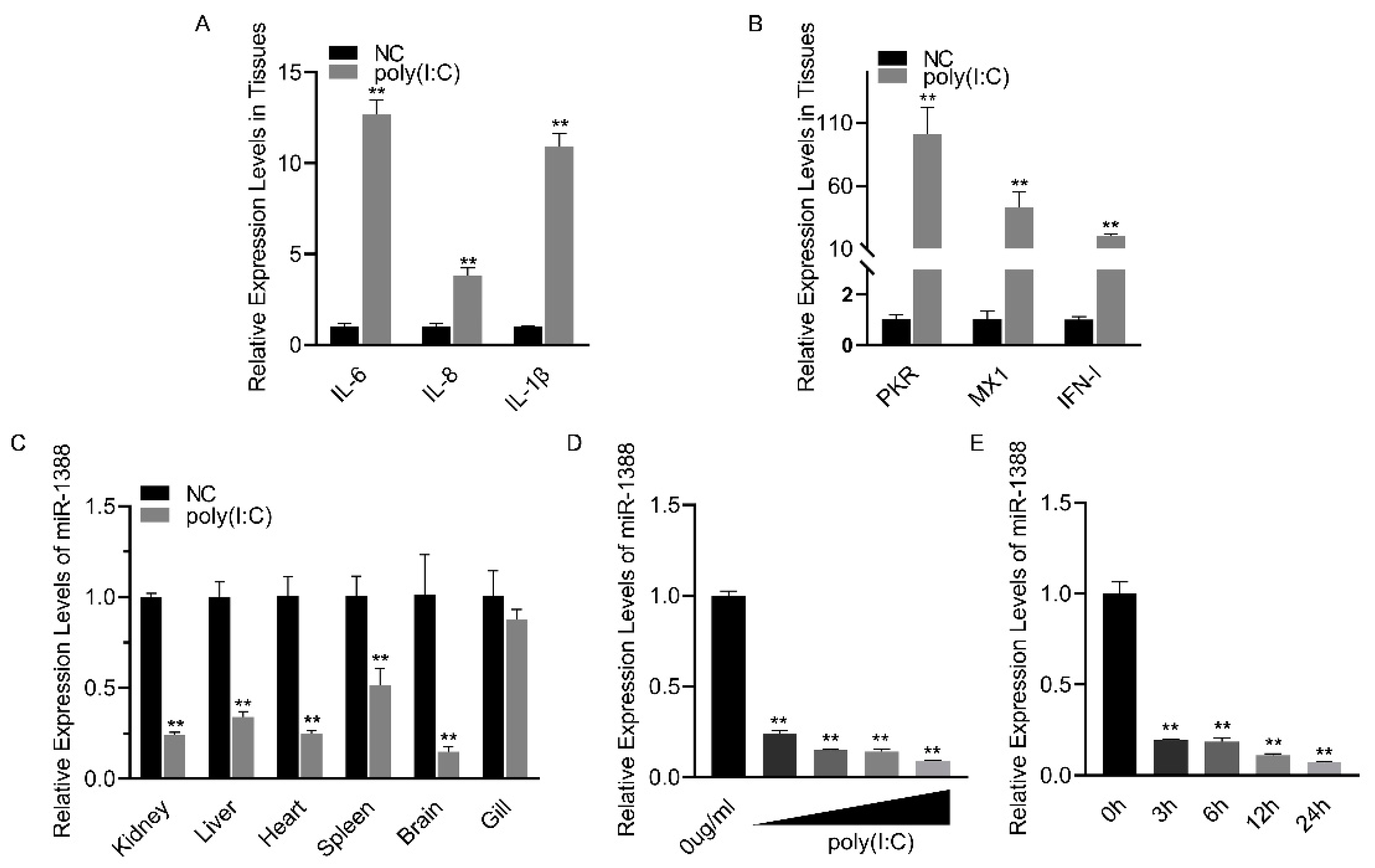
3.1. Poly(I:C) Stimulation Induces Downregulation of miR-1388 Expression
3.2. miR-1388 Has an Effect on the Antiviral Genes Stimulated by poly(I:C)
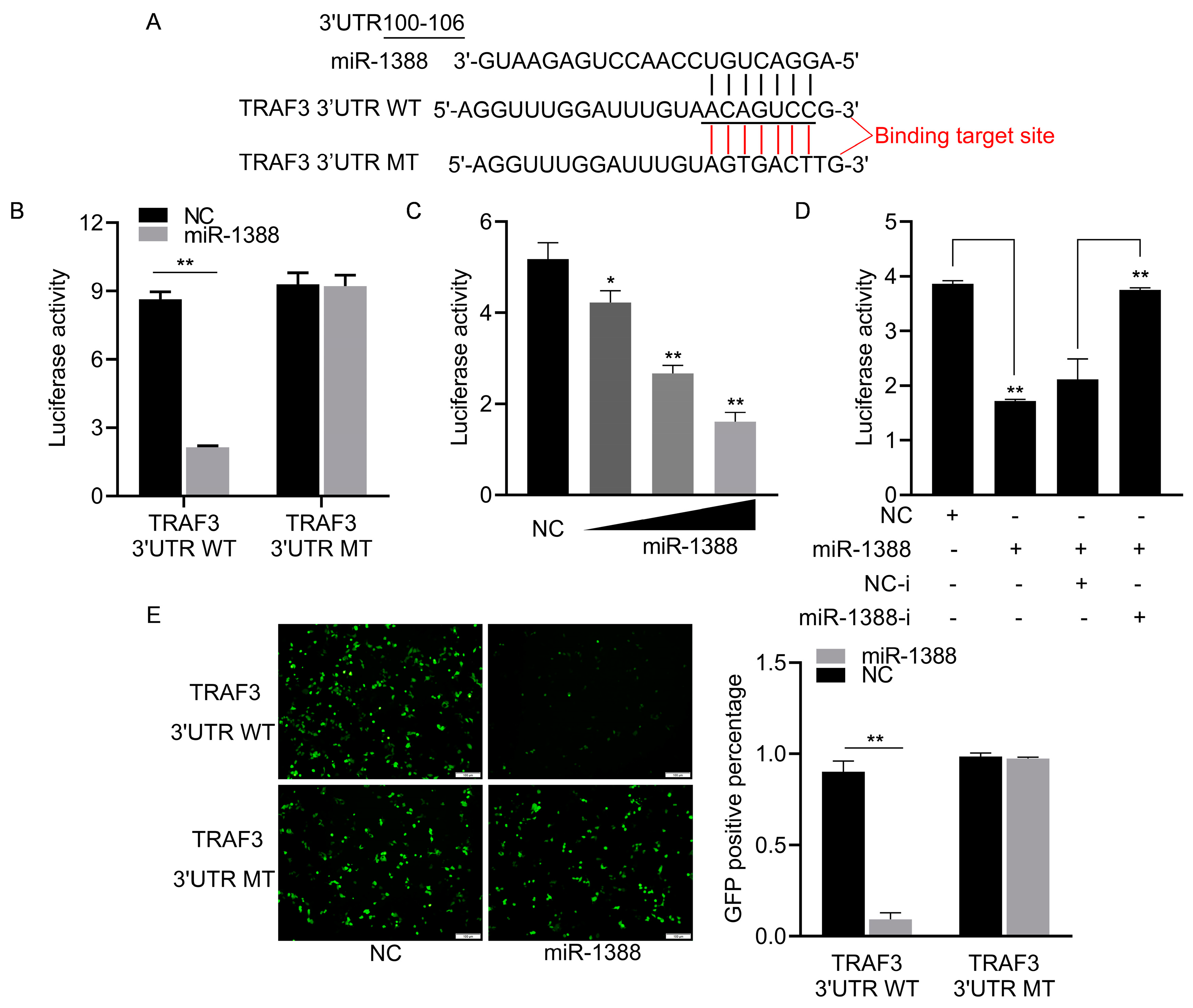
3.3. miR-1388 Showed a Targeting Relationship with TRAF3
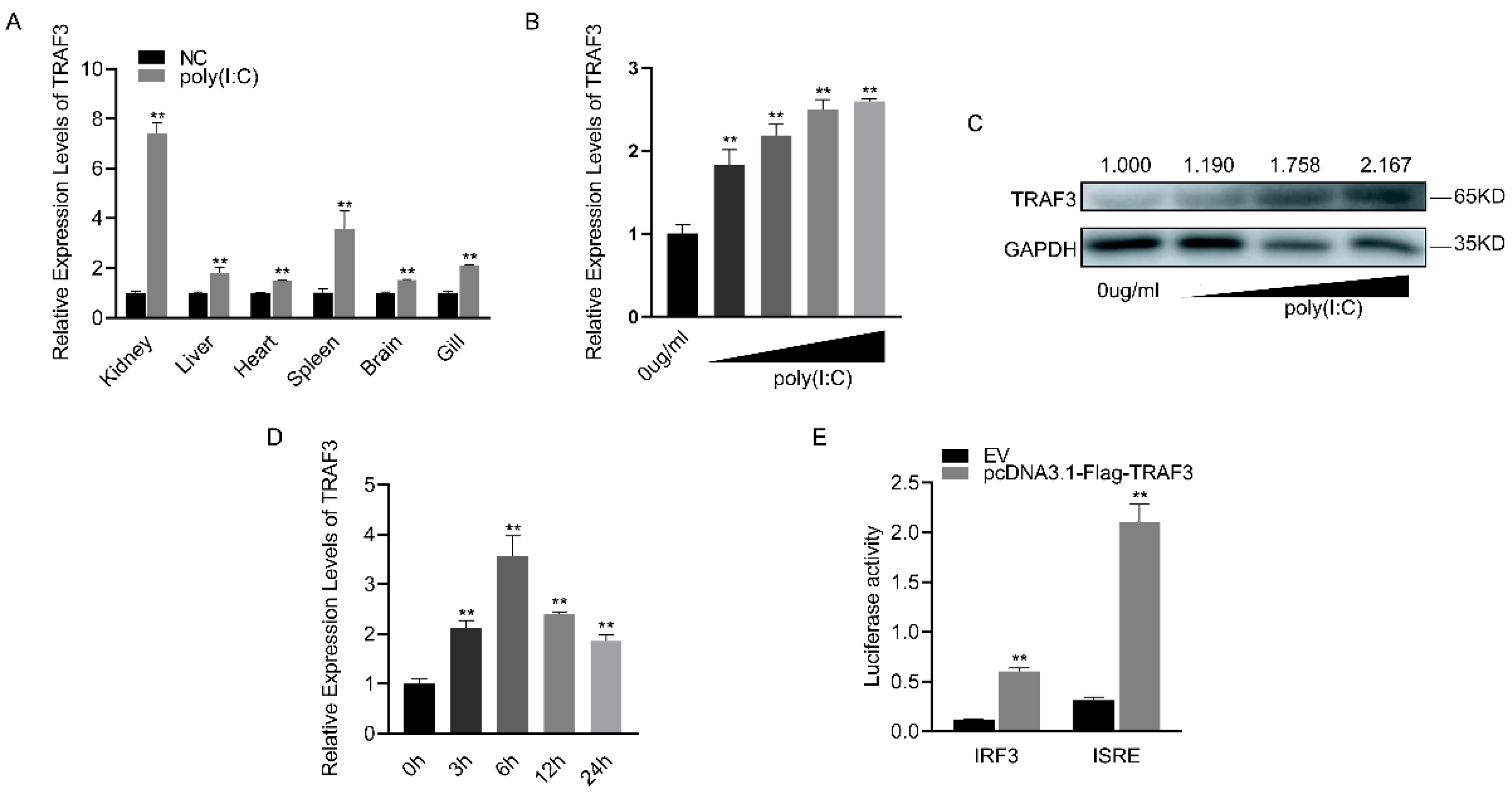
3.4. TRAF3’s Role in Innate Immune Response and IRF3-Mediated Signaling Pathway Activation
3.5. Correlation between miR-1388 Expression and TRAF3 Levels
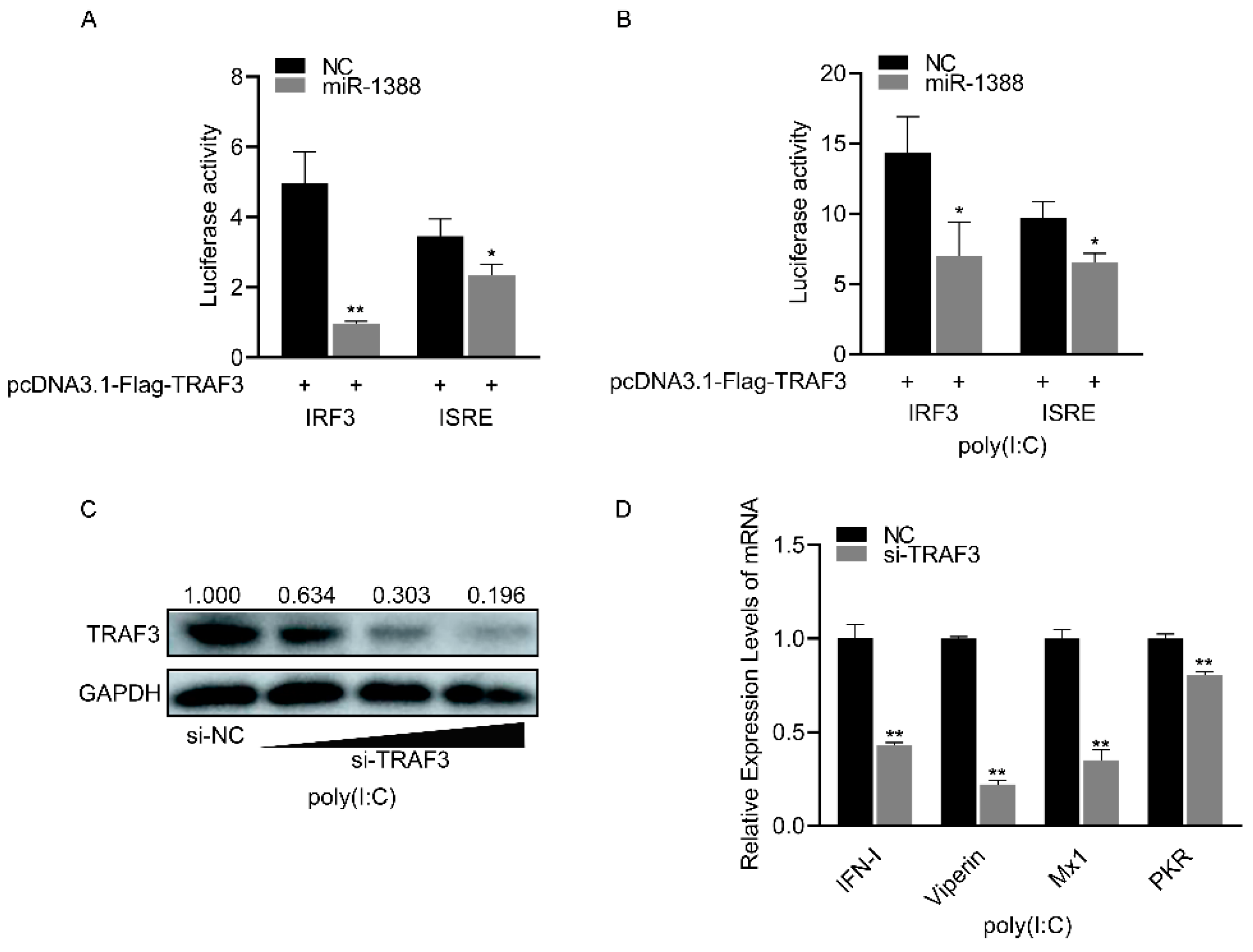
3.6. miR-1388 Regulates Antiviral Genes Primarily through Its Target Gene TRAF3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ruangkiattikul, N.; Rys, D.; Abdissa, K.; Rohde, M.; Semmler, T.; Tegtmeyer, P.-K.; Kalinke, U.; Schwarz, C.; Lewin, A.; Goethe, R. Type I interferon induced by TLR2-TLR4-MyD88-TRIF-IRF3 controls Mycobacterium abscessus subsp. abscessus persistence in murine macrophages via nitric oxide. Int. J. Med. Microbiol. 2019, 309, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.-P.; Zhang, H.-Y.; Liao, B.-W.; Tong, Z.; Xu, Z.-S.; Wang, Y.-Y.; Yang, Y. SRP54 Negatively Regulates IFN-Beta Production and Antiviral Response by Targeting RIG-I and MDA5. Virol. Sin. 2021, 36, 231–240. [Google Scholar] [CrossRef]
- Li, S.-F.; Shao, J.-J.; Zhao, F.-R.; Gong, M.-J.; Xie, Y.-L.; Chang, H.-Y.; Zhang, Y.-G. Antiviral activity of porcine interferon delta 8 against foot-and-mouth disease virus in vitro. Int. Immunopharmacol. 2018, 59, 47–52. [Google Scholar] [CrossRef]
- Mantlo, E.; Bukreyeva, N.; Maruyama, J.; Paessler, S.; Huang, C. Antiviral activities of type I interferons to SARS-CoV-2 infection. Antivir. Res. 2020, 179, 104811. [Google Scholar] [CrossRef]
- Zhou, P.; Liu, D.; Zhang, Q.; Wu, W.; Chen, D.; Luo, R. Antiviral effects of duck type I and type III interferons against Duck Tembusu virus in vitro and in vivo. Vet. Microbiol. 2023, 287, 109889. [Google Scholar] [CrossRef] [PubMed]
- He, J.Q.; Oganesyan, G.; Saha, S.K.; Zarnegar, B.; Cheng, G. TRAF3 and Its Biological Function. In TNF Receptor Associated Factors (TRAFs); Wu, H., Ed.; Springer: New York, NY, USA, 2007; pp. 48–59. [Google Scholar]
- Chen, F.; Chen, L.; Li, Y.; Sang, H.; Zhang, C.; Yuan, S.; Yang, J. TRAF3 Positively Regulates Host Innate Immune Resistance to Influenza A Virus Infection. Front. Cell. Infect. Microbiol. 2022, 12, 839625. [Google Scholar] [CrossRef]
- de Diego, R.P.; Sancho-Shimizu, V.; Lorenzo, L.; Puel, A.; Plancoulaine, S.; Picard, C.; Herman, M.; Cardon, A.; Durandy, A.; Bustamante, J.; et al. Human TRAF3 adaptor molecule deficiency leads to impaired Toll-like receptor 3 response and susceptibility to herpes simplex encephalitis. Immunity 2010, 33, 400–411. [Google Scholar] [CrossRef]
- Rae, W.; Sowerby, J.M.; Verhoeven, D.; Youssef, M.; Kotagiri, P.; Savinykh, N.; Coomber, E.L.; Boneparth, A.; Chan, A.; Gong, C.; et al. Immunodeficiency, autoimmunity, and increased risk of B cell malignancy in humans with TRAF3 mutations. Sci. Immunol. 2022, 7, eabn3800. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Jin, S.; Xian, H.; Zhao, Z.; Wang, L.; Wu, Y.; Zhou, L.; Li, M.; Cui, J. Metabolic enzyme UAP1 mediates IRF3 pyrophosphorylation to facilitate innate immune response. Mol. Cell 2023, 83, 298–313.e8. [Google Scholar] [CrossRef]
- Häcker, H.; Tseng, P.-H.; Karin, M. Expanding TRAF function: TRAF3 as a tri-faced immune regulator. Nat. Rev. Immunol. 2011, 11, 457–468. [Google Scholar] [CrossRef]
- Xiang, Y.; Zhang, W.; Jia, P.; Lu, X.; Liu, W.; Yi, M.; Jia, K. E3 Ubiquitin Ligase RNF114 Inhibits Innate Immune Response to Red-Spotted Grouper Nervous Necrosis Virus Infection in Sea Perch by Targeting MAVS and TRAF3 to Mediate Their Degradation. J. Immunol. 2021, 206, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Iwagami, Y.; Zou, J.; Zhang, H.; Cao, K.; Ji, C.; Kim, M.; Huang, C.-K. Alcohol-mediated miR-34a modulates hepatocyte growth and apoptosis. J. Cell. Mol. Med. 2018, 22, 3987–3995. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-H.; Hwang, J.-S.; Oh, S.-H.; Shin, Y.-J. MiR-302a Regenerates Human Corneal Endothelial Cells against IFN-γ-Induced Cell Death. Cells 2023, 12, 36. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Chu, L.; Li, Y.; Wang, Q.; Zhu, J.; Wang, C.; Cui, S. miR-23a/b suppress cGAS-mediated innate and autoimmunity. Cell. Mol. Immunol. 2021, 18, 1235–1248. [Google Scholar] [CrossRef] [PubMed]
- Abdellaoui, N.; Kim, D.E.; Jeon, Y.H.; Kim, M.S. Antiviral effect of miR-155 in Epithelioma papulosum cyprini (EPC) cells against viral hemorrhagic septicemia virus (VHSV) infection. Fish Shellfish Immunol. 2023, 140, 108937. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Chu, Q.; Cui, J. Rhabdovirus-Inducible MicroRNA-210 Modulates Antiviral Innate Immune Response via Targeting STING/MITA in Fish. J. Immunol. 2018, 201, 982–994. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Hemida, M.G.; Qiu, Y.; Hanson, P.J.; Zhang, H.M.; Yang, D. MiR-126 promotes coxsackievirus replication by mediating cross-talk of ERK1/2 and Wnt/β-catenin signal pathways. Cell. Mol. Life Sci. 2013, 70, 4631–4644. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Guan, J.; Li, R.; Li, X.; Niu, J.; Shang, R.; Qi, J.; Wang, X. miR-1388 regulates the expression of nectin2l in Paralichthys olivaceus. Comp. Biochem. Physiol. Part D Genom. Proteom. 2018, 28, 9–16. [Google Scholar] [CrossRef]
- Liu, H.; Xie, J.-F.; Yu, H.; Ma, Z.; Yu, Y.-Y.; Yang, Y. The early response expression profiles of miRNA-mRNA in farmed yellow catfish (Pelteobagrus fulvidraco) challenged with Edwardsiella tarda infection. Dev. Comp. Immunol. 2021, 119, 104018. [Google Scholar] [CrossRef]
- Xu, Q.; Cai, C.; Hu, X.; Liu, Y.; Guo, Y.; Hu, P.; Chen, Z.; Peng, S.; Zhang, D.; Jiang, S.; et al. Evolutionary suppression of erythropoiesis via the modulation of TGF-β signalling in an Antarctic icefish. Mol. Ecol. 2015, 24, 4664–4678. [Google Scholar] [CrossRef]
- Wang, X.; Chen, X.; Sun, X.; Ao, J. Comparative analysis of miRNAs and mRNAs in large yellow croaker head kidney cells (LYCK) provided novel insights into the redox regulation of fish. Sci. Total Environ. 2024, 918, 170503. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, W.-F.; Huang, C.-X.; Li, X.-H.; Liu, H.; Wang, H.-L. miR-731 modulates the zebrafish heart morphogenesis via targeting Calcineurin/Nfatc3a pathway. Biochim. Biophys. Acta (BBA) Gen. Subj. 2022, 1866, 130133. [Google Scholar] [CrossRef]
- Ma, Z.; Yang, J.; Zhang, Q.; Xu, C.; Wei, J.; Sun, L.; Wang, D.; Tao, W. miR-133b targets tagln2 and functions in tilapia oogenesis. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2021, 256, 110637. [Google Scholar] [CrossRef]
- Liu, M.; Tang, H.; Gao, K.; Zhang, X.; Yang, Z.; Gao, Y.; Shan, X. Identification and Characterization of Immune-Associated MicroRNAs in Silver Carp (Hypophthalmichthys molitrix) Responding to Aeromonas veronii and LPS Stimulation. Animals 2024, 14, 285. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Song, X.; Xie, X.; Li, W.; Lu, L.; Chen, S.; Wu, H.; Feng, H. TRAF3 enhances STING-mediated antiviral signaling during the innate immune activation of black carp. Dev. Comp. Immunol. 2018, 88, 83–93. [Google Scholar] [CrossRef]
- Wu, S.; Sun, M.; Zhang, L.; Kang, S.; Liao, J.; Zhu, Z.; Chen, H.; Xu, Z.; Xu, L.; Zhang, X.; et al. Grouper TRAF3 inhibits nodavirus infection by regulating the STING-mediated antiviral signaling pathway. Fish Shellfish Immunol. 2022, 123, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Yu, T.; Gan, S.; Wang, Y.; Pei, Y.; Zhao, Q.; Pei, S.; Hao, S.; Yuan, J.; Xu, J.; et al. TRIM24 facilitates antiviral immunity through mediating K63-linked TRAF3 ubiquitination. J. Exp. Med. 2020, 217, e20192083. [Google Scholar] [CrossRef]
- Xu, Y.; Cheng, G.; Baltimore, D. Targeted disruption of TRAF3 leads to postnatal lethality and defective T-dependent immune responses. Immunity 1996, 5, 407–415. [Google Scholar] [CrossRef]
- Häcker, H.; Redecke, V.; Blagoev, B.; Kratchmarova, I.; Hsu, L.-C.; Wang, G.G.; Kamps, M.P.; Raz, E.; Wagner, H.; Häcker, G.; et al. Specificity in Toll-like receptor signalling through distinct effector functions of TRAF3 and TRAF6. Nature 2006, 439, 204–207. [Google Scholar] [CrossRef]
- Oganesyan, G.; Saha, S.K.; Guo, B.; He, J.Q.; Shahangian, A.; Zarnegar, B.; Perry, A.; Cheng, G. Critical role of TRAF3 in the Toll-like receptor-dependent and -independent antiviral response. Nature 2006, 439, 208–211. [Google Scholar] [CrossRef]
- Lin, M.; Ji, X.; Lv, Y.; Cui, D.; Xie, J. The Roles of TRAF3 in Immune Responses. Dis. Markers 2023, 2023, 7787803. [Google Scholar] [CrossRef]
- Thakur, A.; Kumar, M. AntiVIRmiR: A repository of host antiviral miRNAs and their expression along with experimentally validated viral miRNAs and their targets. Front. Genet. 2022, 13, 971852. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, X.; Yao, Y.; Zhu, Y.; Zheng, X.; Liu, F.; Feng, W. Inducible miR-150 Inhibits Porcine Reproductive and Respiratory Syndrome Virus Replication by Targeting Viral Genome and Suppressor of Cytokine Signaling 1. Viruses 2022, 14, 1485. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Ke, X.; Wang, M.; He, S.; Li, Q.; Zheng, C.; Zhang, Z.; Liu, Y.; Wang, H. Human MicroRNA hsa-miR-296-5p Suppresses Enterovirus 71 Replication by Targeting the Viral Genome. J. Virol. 2013, 87, 5645–5656. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xia, S.; Yang, X.; Chen, H.; Li, F.; Liu, F.; Chen, Z. Human Cytomegalovirus Encoded miR-US25-1-5p Attenuates CD147/EMMPRIN-Mediated Early Antiviral Response. Viruses 2017, 9, 365. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lin, S.; Zhao, Z.; Xu, P.; Gao, K.; Qian, H.; Zhang, Z.; Guo, X. Functional analysis of a putative Bombyx mori cypovirus miRNA BmCPV-miR-10 and its effect on virus replication. Insect Mol. Biol. 2021, 30, 552–565. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Zhou, Q.; Wang, H.; Li, Q.; Chen, Z.; Lin, Y.; Yi, L.; Jiang, S.; Chen, C.; Deng, Y. MiRNA-338-3p Inhibits Neuroinflammation in the Corpus Callosum of LCV-LPS Rats Via STAT1 Signal Pathway. Cell. Mol. Neurobiol. 2023, 43, 3669–3692. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.; Lii, Y.E.; Chellappan, P.; Lei, L.; Peralta, K.; Jiang, C.; Guo, J.; Coaker, G.; Jin, H. miRNA863-3p sequentially targets negative immune regulator ARLPKs and positive regulator SERRATE upon bacterial infection. Nat. Commun. 2016, 7, 11324. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Zheng, W.; Sun, Y.; Xu, T. microRNA-489 negatively modulates RIG-I signaling pathway via targeting TRAF6 in miiuy croaker after poly(I:C) stimulation. Fish Shellfish Immunol. 2021, 113, 61–68. [Google Scholar] [CrossRef]
- Andreassen, R.; Høyheim, B. miRNAs associated with immune response in teleost fish. Dev. Comp. Immunol. 2017, 75, 77–85. [Google Scholar] [CrossRef]
- Gao, W.; Chang, R.; Sun, Y.; Xu, T. MicroRNA-2187 Modulates the NF-κB and IRF3 Pathway in Teleost Fish by Targeting TRAF6. Front. Immunol. 2021, 12, 647202. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cheng, Y.; Wang, L.; Sun, A.; Lin, Z.; Zhu, W.; Wang, Z.; Ma, J.; Wang, H.; Yan, Y.; et al. Chicken miR-126-5p negatively regulates antiviral innate immunity by targeting TRAF3. Vet. Res. 2022, 53, 82. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chu, Q.; Chang, R.; Xu, T. Inducible MicroRNA-217 Inhibits NF-κB– and IRF3-Driven Immune Responses in Lower Vertebrates through Targeting TAK1. J. Immunol. 2020, 205, 1620–1632. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.; Zheng, W.; Sun, Y.; Xu, T. microRNA-1388-5p inhibits NF-κB signaling pathway in miiuy croaker through targeting IRAK1. Dev. Comp. Immunol. 2021, 119, 104025. [Google Scholar] [CrossRef] [PubMed]
- Oshiumi, H. Circulating Extracellular Vesicles Carry Immune Regulatory miRNAs and Regulate Vaccine Efficacy and Local Inflammatory Response After Vaccination. Front. Immunol. 2021, 12, 685344. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Hazama, S.; Iida, M.; Tsunedomi, R.; Takenouchi, H.; Nakajima, M.; Tokumitsu, Y.; Kanekiyo, S.; Shindo, Y.; Tomochika, S.; et al. miR-125b-1 and miR-378a are predictive biomarkers for the efficacy of vaccine treatment against colorectal cancer. Cancer Sci. 2017, 108, 2229–2238. [Google Scholar] [CrossRef] [PubMed]
- Barnes, D.; Vignuzzi, M.K.M.F.; Saksela, K.V.M.F.; Andino, R.S.K.F.; Andino, R. Harnessing endogenous miRNAs to control virus tissue tropism as a strategy for developing attenuated virus vaccines. Cell Host Microbe 2008, 4, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Yang, T.; Liu, X.; Xiong, F.; Luo, J.; Yi, Y.; Fan, J.; Chen, Z.; Tan, W.-S. MiRNA Targeted NP Genome of Live Attenuated Influenza Vaccines Provide Cross-Protection against a Lethal Influenza Virus Infection. Vaccines 2020, 8, 65. [Google Scholar] [CrossRef]
- Yee, T.I.P.; Poh, L.C. Development of Novel miRNA-based Vaccines and Antivirals against Enterovirus 71. Curr. Pharm. Des. 2016, 22, 6694–6700. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, K.; Liu, M.; Tang, H.; Ma, Z.; Pan, H.; Zhang, X.; Inam, M.; Shan, X.; Gao, Y.; Wang, G. Downregulation of miR-1388 Regulates the Expression of Antiviral Genes via Tumor Necrosis Factor Receptor (TNFR)-Associated Factor 3 Targeting Following poly(I:C) Stimulation in Silver Carp (Hypophthalmichthys molitrix). Biomolecules 2024, 14, 694. https://doi.org/10.3390/biom14060694
Gao K, Liu M, Tang H, Ma Z, Pan H, Zhang X, Inam M, Shan X, Gao Y, Wang G. Downregulation of miR-1388 Regulates the Expression of Antiviral Genes via Tumor Necrosis Factor Receptor (TNFR)-Associated Factor 3 Targeting Following poly(I:C) Stimulation in Silver Carp (Hypophthalmichthys molitrix). Biomolecules. 2024; 14(6):694. https://doi.org/10.3390/biom14060694
Chicago/Turabian StyleGao, Kun, Meng Liu, Huan Tang, Zhenhua Ma, Hanyu Pan, Xiqing Zhang, Muhammad Inam, Xiaofeng Shan, Yunhang Gao, and Guiqin Wang. 2024. "Downregulation of miR-1388 Regulates the Expression of Antiviral Genes via Tumor Necrosis Factor Receptor (TNFR)-Associated Factor 3 Targeting Following poly(I:C) Stimulation in Silver Carp (Hypophthalmichthys molitrix)" Biomolecules 14, no. 6: 694. https://doi.org/10.3390/biom14060694