
Alterations in the Hypothalamic–Pituitary–Adrenal Axis as a Response to Experimental Autoimmune Encephalomyelitis in Dark Agouti Rats of Both Sexes

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. EAE Induction and Evaluation
2.3. Blood and Tissue Collection
2.4. Histological Staining and Volumetric Measurements
2.5. Immunohistochemistry
2.6. Steroid Extraction
2.7. Hormone Measurements
2.8. RNA Isolation and qPCR Analysis
2.9. Protein Isolation and Western Blot
2.10. Statistical Analysis and Data Presentation
3. Results
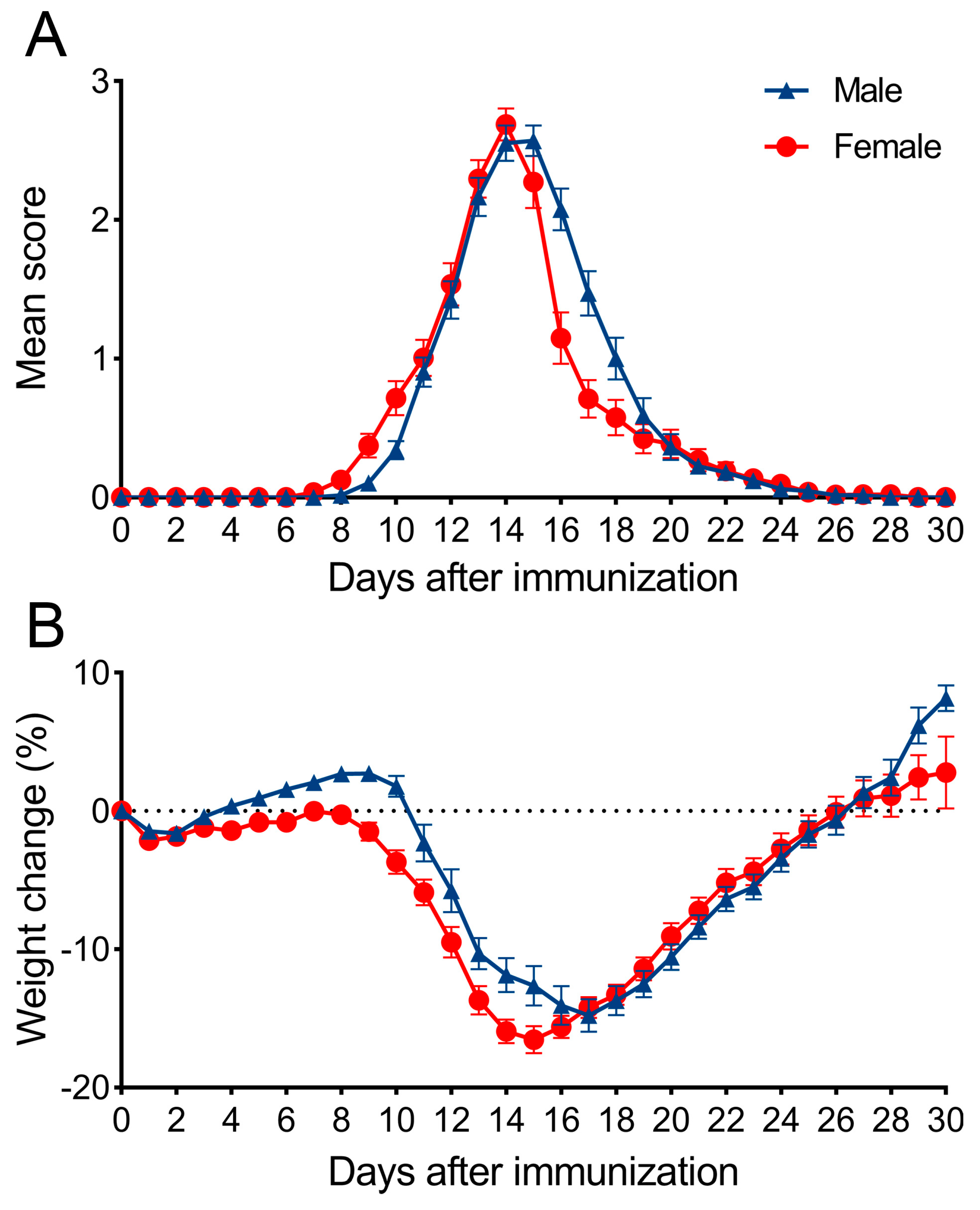
3.1. Characterization of EAE Course and Parameters in Male and Female DA Rats
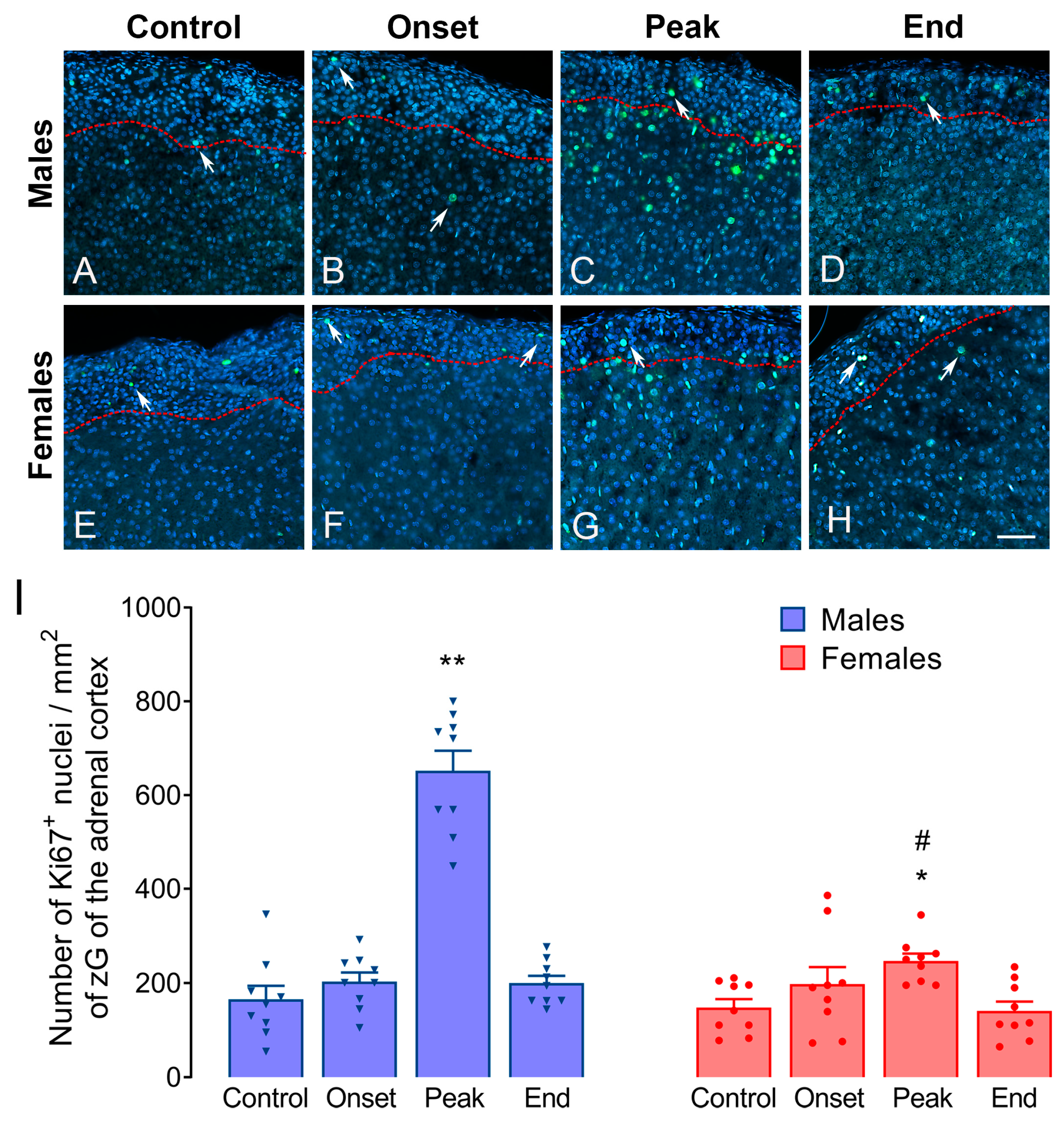
3.2. Adrenal Weight and Volumetric Measures in Male and Female Rats during EAE
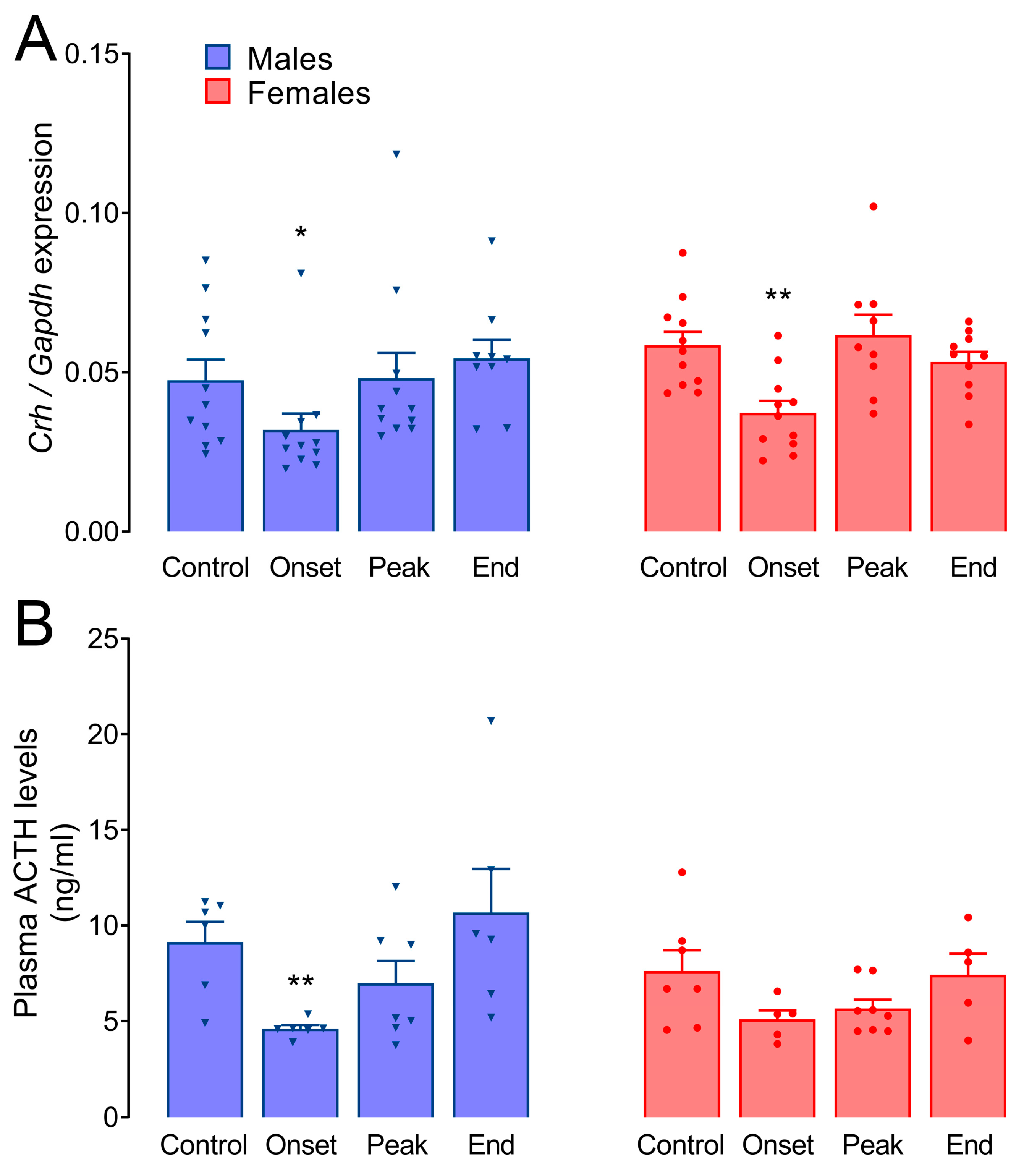
3.3. Crh Expression in the Hypothalamus and Circulating ACTH Levels during EAE
3.4. Corticosterone Levels in Males and Females during EAE
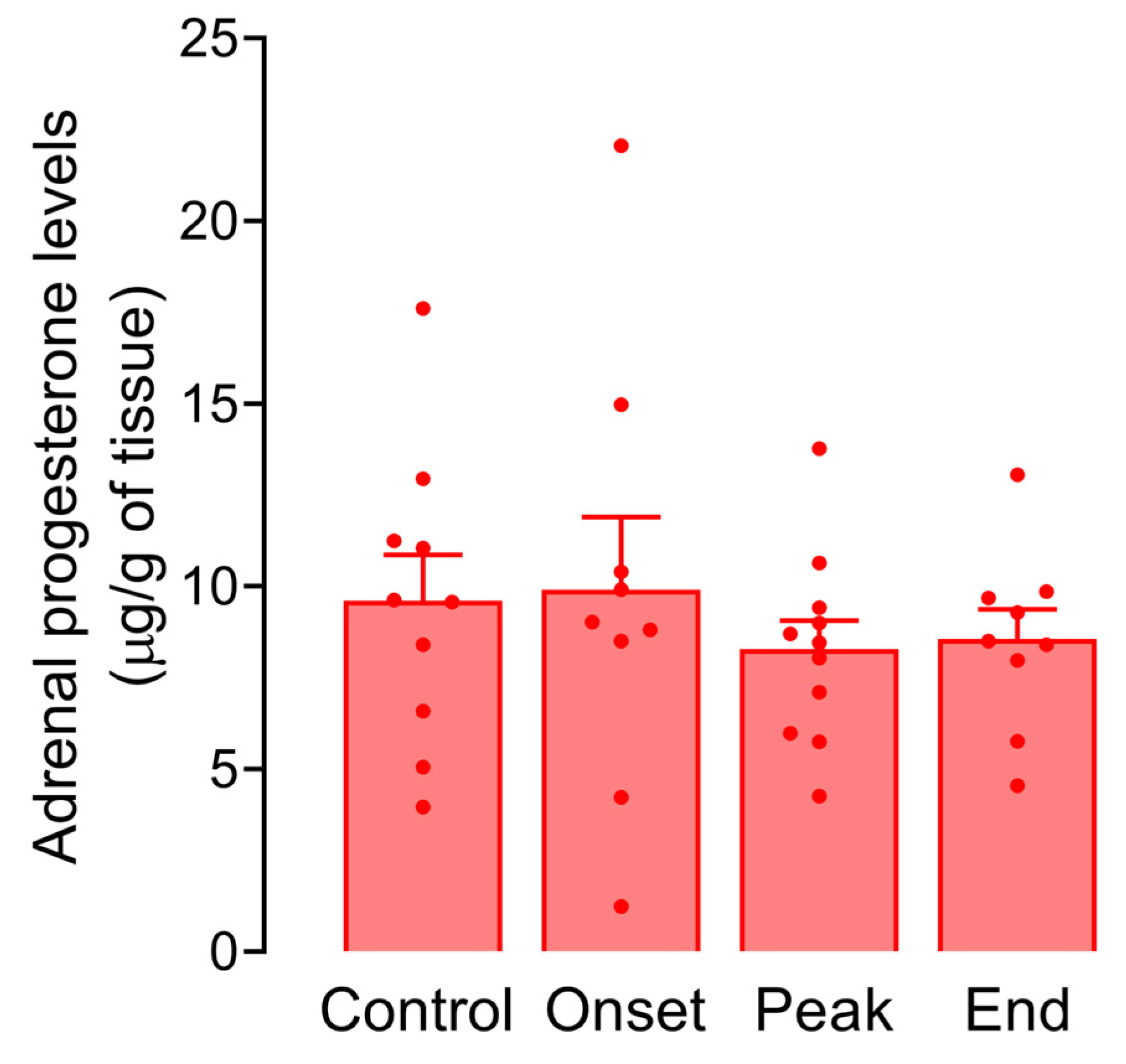
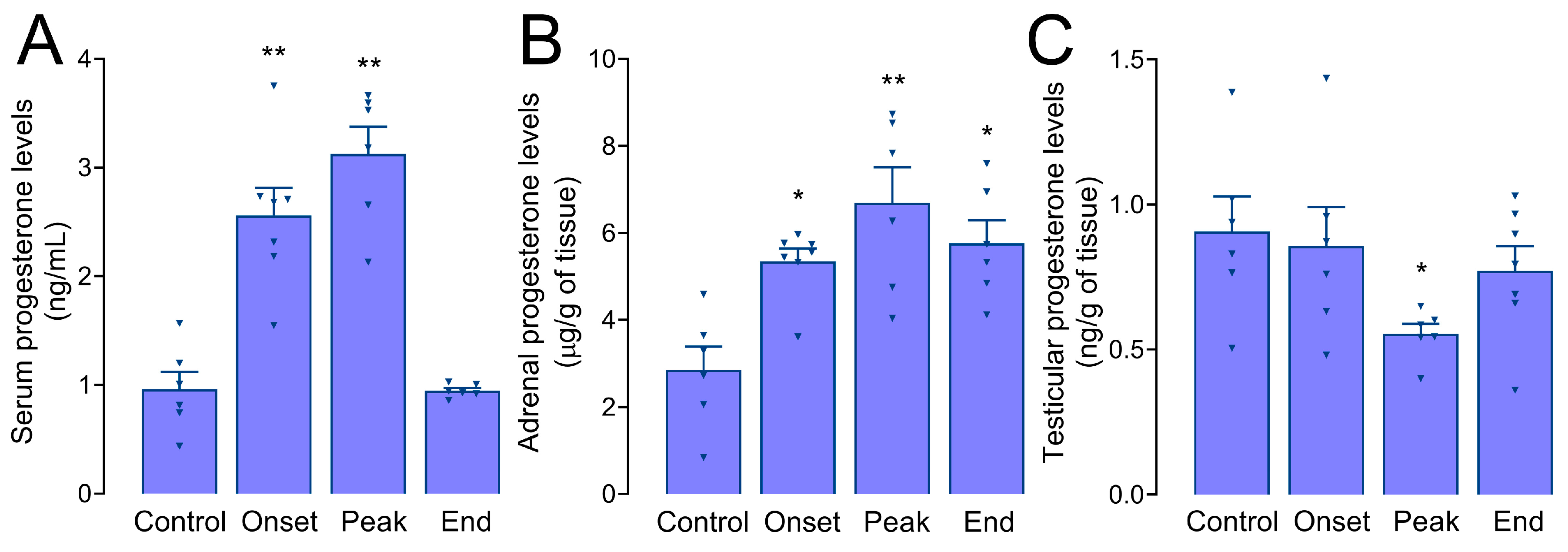
3.5. Progesterone Levels in Serum, Adrenals, and Testes of Male Rats during EAE
3.6. Gene Expression of Adrenal Steroidogenic Pathway Components during EAE
3.7. Protein Levels of StAR, P450scc, and 3β-HSD in Adrenal Glands
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Thompson, A.J.; Baranzini, S.E.; Geurts, J.; Hemmer, B.; Ciccarelli, O. Multiple sclerosis. Lancet 2018, 391, 1622–1636. [Google Scholar] [CrossRef] [PubMed]
- Coyle, P.K. What Can We Learn from Sex Differences in MS? J. Pers. Med. 2021, 11, 1006. [Google Scholar] [CrossRef]
- Voskuhl, R. Sex differences in autoimmune diseases. Biol. Sex Differ. 2011, 2, 1. [Google Scholar] [CrossRef] [PubMed]
- Duma, D.; Collins, J.B.; Chou, J.W.; Cidlowski, J.A. Sexually dimorphic actions of glucocorticoids provide a link to inflammatory diseases with gender differences in prevalence. Sci. Signal. 2010, 3, ra74. [Google Scholar] [CrossRef]
- Milosevic, A.; Janjic, M.M.; Lavrnja, I.; Savic, D.; Bozic, I.D.; Tesovic, K.; Jakovljevic, M.; Pekovic, S.; Stojilkovic, S.S.; Bjelobaba, I. The sex-specific patterns of changes in hypothalamic-pituitary-gonadal axis during experimental autoimmune encephalomyelitis. Brain Behav. Immun. 2020, 89, 233–244. [Google Scholar] [CrossRef]
- Milosevic, A.; Bjelobaba, I.; Bozic, I.D.; Lavrnja, I.; Savic, D.; Tesovic, K.; Jakovljevic, M.; Stojilkovic, S.S.; Janjic, M.M. Testicular steroidogenesis is suppressed during experimental autoimmune encephalomyelitis in rats. Sci. Rep. 2021, 11, 8996. [Google Scholar] [CrossRef]
- Milosevic, A.; Lavrnja, I.; Savic, D.; Milosevic, K.; Skuljec, J.; Bjelobaba, I.; Janjic, M.M. Rat Ovarian Function Is Impaired during Experimental Autoimmune Encephalomyelitis. Cells 2023, 12, 1045. [Google Scholar] [CrossRef] [PubMed]
- Trifunovic, S.; Stevanovic, I.; Milosevic, A.; Ristic, N.; Janjic, M.; Bjelobaba, I.; Savic, D.; Bozic, I.; Jakovljevic, M.; Tesovic, K.; et al. The Function of the Hypothalamic-Pituitary-Adrenal Axis During Experimental Autoimmune Encephalomyelitis: Involvement of Oxidative Stress Mediators. Front. Neurosci. 2021, 15, 649485. [Google Scholar] [CrossRef] [PubMed]
- Whirledge, S.; Cidlowski, J.A. Glucocorticoids, stress, and fertility. Minerva Endocrinol. 2010, 35, 109–125. [Google Scholar]
- Huitinga, I.; Erkut, Z.A.; van Beurden, D.; Swaab, D.F. The hypothalamo-pituitary-adrenal axis in multiple sclerosis. Ann. N. Y. Acad. Sci. 2003, 992, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, S.D.; Amouret, A.; Muzzi, C.; Vettorazzi, S.; Tuckermann, J.P.; Lühder, F.; Reichardt, H.M. The Role of Glucocorticoids in Inflammatory Diseases. Cells 2021, 10, 2921. [Google Scholar] [CrossRef]
- Bourke, C.H.; Harrell, C.S.; Neigh, G.N. Stress-induced sex differences: Adaptations mediated by the glucocorticoid receptor. Horm. Behav. 2012, 62, 210–218. [Google Scholar] [CrossRef]
- Bjelobaba, I.; Savic, D.; Lavrnja, I. Multiple Sclerosis and Neuroinflammation: The Overview of Current and Prospective Therapies. Curr. Pharm. Des. 2017, 23, 693–730. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Vale, W.W. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialogues Clin. Neurosci. 2006, 8, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.M.; Newell-Price, J.D.C. Chapter 15—The Adrenal Cortex. In Williams Textbook of Endocrinology, 13th ed.; Melmed, S., Polonsky, K.S., Larsen, P.R., Kronenberg, H.M., Eds.; Elsevier: Philadelphia, PA, USA, 2016; pp. 489–555. [Google Scholar] [CrossRef]
- Le Goascogne, C.; Sananès, N.; Gouézou, M.; Takemori, S.; Kominami, S.; Baulieu, E.E.; Robel, P. Immunoreactive cytochrome P-450(17 alpha) in rat and guinea-pig gonads, adrenal glands and brain. J. Reprod. Fertil. 1991, 93, 609–622. [Google Scholar] [CrossRef] [PubMed]
- van Weerden, W.M.; Bierings, H.G.; van Steenbrugge, G.J.; de Jong, F.H.; Schröder, F.H. Adrenal glands of mouse and rat do not synthesize androgens. Life Sci. 1992, 50, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.J.; Azhar, S.; Kraemer, F.B. ACTH Regulation of Adrenal SR-B1. Front. Endocrinol. 2016, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, V.; Stocco, D.M.; Clark, B.J. Current knowledge on the acute regulation of steroidogenesis. Biol. Reprod. 2018, 99, 13–26. [Google Scholar] [CrossRef]
- Mitani, F. Functional zonation of the rat adrenal cortex: The development and maintenance. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2014, 90, 163–183. [Google Scholar] [CrossRef]
- Chapman, K.; Holmes, M.; Seckl, J. 11β-hydroxysteroid dehydrogenases: Intracellular gate-keepers of tissue glucocorticoid action. Physiol. Rev. 2013, 93, 1139–1206. [Google Scholar] [CrossRef]
- Shimojo, M.; Whorwood, C.B.; Stewart, P.M. 11 beta-Hydroxysteroid dehydrogenase in the rat adrenal. J. Mol. Endocrinol. 1996, 17, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Potrebić, M.S.; Pavković, Ž.Z.; Srbovan, M.M.; Dmura, G.M.; Pešić, V.T. Changes in the Behavior and Body Weight of Mature, Adult Male Wistar Han Rats after Reduced Social Grouping and Social Isolation. J. Am. Assoc. Lab. Anim. Sci. 2022, 61, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Perović, M.; Jović, M.; Todorović, S.; Đorđević, A.M.; Milanović, D.; Kanazir, S.; Lončarević-Vasiljković, N. Neuroprotective effects of food restriction in a rat model of traumatic brain injury—the role of glucocorticoid signaling. Nutr. Neurosci. 2022, 25, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Lerario, A.M.; Finco, I.; LaPensee, C.; Hammer, G.D. Molecular Mechanisms of Stem/Progenitor Cell Maintenance in the Adrenal Cortex. Front. Endocrinol. 2017, 8, 52. [Google Scholar] [CrossRef]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Koganti, P.P.; Zhao, A.H.; Selvaraj, V. Exogenous cholesterol acquisition signaling in LH-responsive MA-10 Leydig cells and in adult mice. J. Endocrinol. 2022, 254, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Goel, N.; Workman, J.L.; Lee, T.T.; Innala, L.; Viau, V. Sex differences in the HPA axis. Compr. Physiol. 2014, 4, 1121–1155. [Google Scholar] [CrossRef]
- Seale, J.V.; Wood, S.A.; Atkinson, H.C.; Harbuz, M.S.; Lightman, S.L. Gonadal steroid replacement reverses gonadectomy-induced changes in the corticosterone pulse profile and stress-induced hypothalamic-pituitary-adrenal axis activity of male and female rats. J. Neuroendocrinol. 2004, 16, 989–998. [Google Scholar] [CrossRef]
- Seeman, T.E.; Singer, B.; Wilkinson, C.W.; McEwen, B. Gender differences in age-related changes in HPA axis reactivity. Psychoneuroendocrinology 2001, 26, 225–240. [Google Scholar] [CrossRef]
- Uhart, M.; Chong, R.Y.; Oswald, L.; Lin, P.I.; Wand, G.S. Gender differences in hypothalamic-pituitary-adrenal (HPA) axis reactivity. Psychoneuroendocrinology 2006, 31, 642–652. [Google Scholar] [CrossRef]
- Michelson, D.; Stone, L.; Galliven, E.; Magiakou, M.A.; Chrousos, G.P.; Sternberg, E.M.; Gold, P.W. Multiple sclerosis is associated with alterations in hypothalamic-pituitary-adrenal axis function. J. Clin. Endocrinol. Metab. 1994, 79, 848–853. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Lightman, S.L. The neuroendocrine axis in patients with multiple sclerosis. Brain 1997, 120 Pt 6, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Grasser, A.; Möller, A.; Backmund, H.; Yassouridis, A.; Holsboer, F. Heterogeneity of hypothalamic-pituitary-adrenal system response to a combined dexamethasone-CRH test in multiple sclerosis. Exp. Clin. Endocrinol. Diabetes 1996, 104, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Fassbender, K.; Schmidt, R.; Mössner, R.; Kischka, U.; Kühnen, J.; Schwartz, A.; Hennerici, M. Mood disorders and dysfunction of the hypothalamic-pituitary-adrenal axis in multiple sclerosis: Association with cerebral inflammation. Arch. Neurol. 1998, 55, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Then Bergh, F.; Kümpfel, T.; Trenkwalder, C.; Rupprecht, R.; Holsboer, F. Dysregulation of the hypothalamo-pituitary-adrenal axis is related to the clinical course of MS. Neurology 1999, 53, 772–777. [Google Scholar] [CrossRef]
- Melief, J.; de Wit, S.J.; van Eden, C.G.; Teunissen, C.; Hamann, J.; Uitdehaag, B.M.; Swaab, D.; Huitinga, I. HPA axis activity in multiple sclerosis correlates with disease severity, lesion type and gene expression in normal-appearing white matter. Acta Neuropathol. 2013, 126, 237–249. [Google Scholar] [CrossRef]
- Hamidovic, A.; Karapetyan, K.; Serdarevic, F.; Choi, S.H.; Eisenlohr-Moul, T.; Pinna, G. Higher Circulating Cortisol in the Follicular vs. Luteal Phase of the Menstrual Cycle: A Meta-Analysis. Front. Endocrinol. 2020, 11, 311. [Google Scholar] [CrossRef]
- Guven Yorgun, Y.; Ozakbas, S. Effect of hormonal changes on the neurological status in the menstrual cycle of patient with multiple sclerosis. Clin. Neurol. Neurosurg. 2019, 186, 105499. [Google Scholar] [CrossRef]
- Pérez-Nievas, B.G.; García-Bueno, B.; Madrigal, J.L.; Leza, J.C. Chronic immobilisation stress ameliorates clinical score and neuroinflammation in a MOG-induced EAE in Dark Agouti rats: Mechanisms implicated. J. Neuroinflamm. 2010, 7, 60. [Google Scholar] [CrossRef]
- Stefferl, A.; Linington, C.; Holsboer, F.; Reul, J.M. Susceptibility and resistance to experimental allergic encephalomyelitis: Relationship with hypothalamic-pituitary-adrenocortical axis responsiveness in the rat. Endocrinology 1999, 140, 4932–4938. [Google Scholar] [CrossRef]
- Toews, J.N.C.; Hammond, G.L.; Viau, V. Liver at the nexus of rat postnatal HPA axis maturation and sexual dimorphism. J. Endocrinol. 2021, 248, R1–R17. [Google Scholar] [CrossRef]
- Kotelevtsev, Y.; Holmes, M.C.; Burchell, A.; Houston, P.M.; Schmoll, D.; Jamieson, P.; Best, R.; Brown, R.; Edwards, C.R.; Seckl, J.R.; et al. 11beta-hydroxysteroid dehydrogenase type 1 knockout mice show attenuated glucocorticoid-inducible responses and resist hyperglycemia on obesity or stress. Proc. Natl. Acad. Sci. USA 1997, 94, 14924–14929. [Google Scholar] [CrossRef] [PubMed]
- Mullins, L.J.; Kenyon, C.J.; Bailey, M.A.; Conway, B.R.; Diaz, M.E.; Mullins, J.J. Mineralocorticoid Excess or Glucocorticoid Insufficiency: Renal and Metabolic Phenotypes in a Rat Hsd11b2 Knockout Model. Hypertension 2015, 66, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Reder, A.T.; Makowiec, R.L.; Lowy, M.T. Adrenal size is increased in multiple sclerosis. Arch. Neurol. 1994, 51, 151–154. [Google Scholar] [CrossRef]
- Quintanar-Stephano, A.; Chavira-Ramírez, R.; Kovacs, K.; Berczi, I. Neurointermediate pituitary lobectomy decreases the incidence and severity of experimental autoimmune encephalomyelitis in Lewis rats. J. Endocrinol. 2005, 184, 51–58. [Google Scholar] [CrossRef]
- Quintanar-Stephano, A.; Organista-Esparza, A.; Chavira-Ramírez, R.; Kovacs, K.; Berczi, I. Effects of neurointermediate pituitary lobectomy and desmopressin on acute experimental autoimmune encephalomyelitis in Lewis rats. Neuroimmunomodulation 2012, 19, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Ruocco, H.H.; Fernandes, G.A.; Namer, I.J.; Depaulis, A.; Levy, S. Hypothalamic response to experimental allergic encephalomyelitis: Role of substance P. Neuroimmunomodulation 2004, 11, 28–35. [Google Scholar] [CrossRef]
- Erkut, Z.A.; Hofman, M.A.; Ravid, R.; Swaab, D.F. Increased activity of hypothalamic corticotropin-releasing hormone neurons in multiple sclerosis. J. Neuroimmunol. 1995, 62, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Purba, J.S.; Raadsheer, F.C.; Hofman, M.A.; Ravid, R.; Polman, C.H.; Kamphorst, W.; Swaab, D.F. Increased number of corticotropin-releasing hormone expressing neurons in the hypothalamic paraventricular nucleus of patients with multiple sclerosis. Neuroendocrinology 1995, 62, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Harbuz, M. Neuroendocrine function and chronic inflammatory stress. Exp. Physiol. 2002, 87, 519–525. [Google Scholar] [CrossRef]
- Calzà, L.; Giardino, L.; Pozza, M.; Micera, A.; Aloe, L. Time-course changes of nerve growth factor, corticotropin-releasing hormone, and nitric oxide synthase isoforms and their possible role in the development of inflammatory response in experimental allergic encephalomyelitis. Proc. Natl. Acad. Sci. USA 1997, 94, 3368–3373. [Google Scholar] [CrossRef]
- Keller-Wood, M.E.; Dallman, M.F. Corticosteroid inhibition of ACTH secretion. Endocr. Rev. 1984, 5, 1–24. [Google Scholar] [CrossRef]
- Stolz, L.; Derouiche, A.; Devraj, K.; Weber, F.; Brunkhorst, R.; Foerch, C. Anticoagulation with warfarin and rivaroxaban ameliorates experimental autoimmune encephalomyelitis. J. Neuroinflamm. 2017, 14, 152. [Google Scholar] [CrossRef]
- Zingg, H.H.; Lefebvre, D.; Almazan, G. Regulation of vasopressin gene expression in rat hypothalamic neurons. Response to osmotic stimulation. J. Biol. Chem. 1986, 261, 12956–12959. [Google Scholar] [CrossRef]
- Jessop, D.S.; Chowdrey, H.S.; Lightman, S.L. Inhibition of rat corticotropin-releasing factor and adrenocorticotropin secretion by an osmotic stimulus. Brain Res. 1990, 523, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, G.; Lightman, S.L.; Kiss, A. Regulation of the hypothalamic-pituitary-adrenal axis during water deprivation. Endocrinology 1993, 132, 241–248. [Google Scholar] [CrossRef]
- Handa, R.J.; Weiser, M.J. Gonadal steroid hormones and the hypothalamo-pituitary-adrenal axis. Front. Neuroendocrinol. 2014, 35, 197–220. [Google Scholar] [CrossRef] [PubMed]
- Gore, A.C.; Attardi, B.; DeFranco, D.B. Glucocorticoid repression of the reproductive axis: Effects on GnRH and gonadotropin subunit mRNA levels. Mol. Cell Endocrinol. 2006, 256, 40–48. [Google Scholar] [CrossRef]
- Kalil, B.; Leite, C.M.; Carvalho-Lima, M.; Anselmo-Franci, J.A. Role of sex steroids in progesterone and corticosterone response to acute restraint stress in rats: Sex differences. Stress 2013, 16, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Krysko, K.M.; Graves, J.S.; Dobson, R.; Altintas, A.; Amato, M.P.; Bernard, J.; Bonavita, S.; Bove, R.; Cavalla, P.; Clerico, M.; et al. Sex effects across the lifespan in women with multiple sclerosis. Ther. Adv. Neurol. Disord. 2020, 13, 1756286420936166. [Google Scholar] [CrossRef]
- Caruso, D.; Melis, M.; Fenu, G.; Giatti, S.; Romano, S.; Grimoldi, M.; Crippa, D.; Marrosu, M.G.; Cavaletti, G.; Melcangi, R.C. Neuroactive steroid levels in plasma and cerebrospinal fluid of male multiple sclerosis patients. J. Neurochem. 2014, 130, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Komane, M.; Avenant, C.; Louw-du Toit, R.; Africander, D.J.; Hapgood, J.P. Differential off-target glucocorticoid activity of progestins used in endocrine therapy. Steroids 2022, 182, 108998. [Google Scholar] [CrossRef] [PubMed]
- Nacka-Aleksić, M.; Djikić, J.; Pilipović, I.; Stojić-Vukanić, Z.; Kosec, D.; Bufan, B.; Arsenović-Ranin, N.; Dimitrijević, M.; Leposavić, G. Male rats develop more severe experimental autoimmune encephalomyelitis than female rats: Sexual dimorphism and diergism at the spinal cord level. Brain Behav. Immun. 2015, 49, 101–118. [Google Scholar] [CrossRef]
- Duque Ede, A.; Munhoz, C.D. The Pro-inflammatory Effects of Glucocorticoids in the Brain. Front. Endocrinol. 2016, 7, 78. [Google Scholar] [CrossRef]
- Gomes, J.A.; Stevens, R.D.; Lewin, J.J., 3rd; Mirski, M.A.; Bhardwaj, A. Glucocorticoid therapy in neurologic critical care. Crit. Care Med. 2005, 33, 1214–1224. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences | Accession Number |
|---|---|---|
| Avp | f: AGGAGAACTACCTGCCCTCG r: AAACCCTCTCGACACTCGG | NM_016992.2 |
| Cyp11a1 | f: ACTTCCTGAGGGAGAACGGC r: TCCATGTTGCCCAGCTTCTC | NM_017286.3 |
| Cyp21a1 | f: ACGATCATCATCCCCAACAT r: CACAGCCAAAGGTCGGTATT | NM_057101.2 |
| Crh | f: AACCTCAACAGAAGTCCCGC r: ACACGCGGAAAAAGTTAGCC | NM_031019.1 |
| Gapdh | f: CAACTCCCTCAAGATTGTCAGCAA r: GGCATGGACTGTGGTCATGA | NM_017008.4 |
| Hsd11b1 | f: AAATACCTCCTCCCCGTCCT r: TTTCTCTTCCGATCCCTTTG | NM_017080.2 |
| Hsd11b2 | f: CAAACCCTTCCCCCACAG r: GGCTGGGCTTTTCTTAACAG | NM_017081.2 |
| Hsd3b1 | f: GACAGGAGCAGGAGGGTTTGTGG r: CTCCTTCTAACATTGTCACCTTGGCCT | NM_001007719.3 |
| Scarb1 | f: GCTTCTGGTGCCCATCATTTAC r: AGCTTGGCTTCTTGCAGTACC | NM_031541.1 |
| Star | f: AGCAAGGAGAGGAAGCTATGC r: GGCACCACCTTACTTAGCACT | NM_031558.3 |
| Antibody | Source and Type | Manufacturer | Dilution |
|---|---|---|---|
| β-actin | Mouse, monoclonal | Sigma 1, A5316 | 1:5000 |
| StAR | Rabbit, polyclonal | Gift from Prof. Vimal Selvaraj [27] | 1:500 |
| P450scc | Rabbit, polyclonal | Protein Tech 2, 13363-1-AP | 1:2000 |
| 3β-HSD | Mouse, monoclonal | Santa Cruz 3, sc-515120 | 1:500 |
| Anti-mouse IgG, HRP-conjugated | Donkey, polyclonal | Santa Cruz 3, sc-2314 | 1:5000 |
| Anti-rabbit IgG, HRP-conjugated | Donkey, polyclonal | Santa Cruz 3, sc-2313 | 1:5000 |
| Males | Females | |
|---|---|---|
| Day of disease onset | 11.07 ± 0.15 | 10.55 ± 0.17 # |
| Duration of symptoms (days) | 9.23 ± 0.56 | 9.44 ± 0.79 |
| Duration of paralysis (days) | 1.88 ± 0.29 | 1.41 ± 0.29 |
| Maximal severity score | 3.18 ± 0.07 | 2.97 ± 0.10 # |
| Cumulative disease severity | 14.64 ± 0.93 | 13.00 ± 1.17 |
| Adrenal Weight (mg) | Adrenal Volume (mm3) | Medulla Volume (mm3) | Cortex Volume (mm3) | zF + zR Volume (mm3) | zG Volume (mm3) | ||
|---|---|---|---|---|---|---|---|
| Males | Control | 30.82 ± 0.77 | 7.32 ± 0.74 | 0.68 ± 0.05 | 6.63 ± 0.71 | 5.55 ± 0.66 | 1.09 ± 0.06 |
| Onset | 36.75 ± 1.44 ** | 7.90 ± 0.41 | 0.68 ± 0.07 | 7.22 ± 0.38 | 6.20 ± 0.32 | 1.02 ± 0.08 | |
| Peak | 39.80 ± 1.19 ** | 10.00 ± 0.48 * | 1.04 ± 0.13 | 8.96 ± 0.36 * | 7.61 ± 0.31 * | 1.34 ± 0.06 | |
| End | 29.77 ± 0.75 | 7.21 ± 0.10 | 0.79 ± 0.05 | 6.43 ± 0.14 | 5.30 ± 0.13 | 1.12 ± 0.05 | |
| Females | Control | 41.67 ± 1.21 ## | 10.34 ± 0.37 # | 0.92 ± 0.08 | 9.42 ± 0.37 # | 7.97 ± 0.30 # | 1.45 ± 0.07 # |
| Onset | 47.40 ± 1.80 ## | 10.34 ± 0.53 | 0.72 ± 0.03 | 9.52 ± 0.32 | 8.42 ± 0.46 # | 1.27 ± 0.10 | |
| Peak | 48.73 ± 1.02 ** ## | 11.77 ± 1.33 | 0.85 ± 0.14 | 10.92 ± 1.20 | 9.37 ± 0.99 | 1.55 ± 0.28 | |
| End | 40.40 ± 2.44 ## | 10.07 ± 1.24 # | 0.90 ± 0.21 | 9.17 ± 1.03 # | 7.75 ± 0.80 # | 1.41 ± 0.27 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milosevic, A.; Milosevic, K.; Zivkovic, A.; Lavrnja, I.; Savic, D.; Bjelobaba, I.; Janjic, M.M. Alterations in the Hypothalamic–Pituitary–Adrenal Axis as a Response to Experimental Autoimmune Encephalomyelitis in Dark Agouti Rats of Both Sexes. Biomolecules 2024, 14, 1020. https://doi.org/10.3390/biom14081020
Milosevic A, Milosevic K, Zivkovic A, Lavrnja I, Savic D, Bjelobaba I, Janjic MM. Alterations in the Hypothalamic–Pituitary–Adrenal Axis as a Response to Experimental Autoimmune Encephalomyelitis in Dark Agouti Rats of Both Sexes. Biomolecules. 2024; 14(8):1020. https://doi.org/10.3390/biom14081020
Chicago/Turabian StyleMilosevic, Ana, Katarina Milosevic, Anica Zivkovic, Irena Lavrnja, Danijela Savic, Ivana Bjelobaba, and Marija M. Janjic. 2024. "Alterations in the Hypothalamic–Pituitary–Adrenal Axis as a Response to Experimental Autoimmune Encephalomyelitis in Dark Agouti Rats of Both Sexes" Biomolecules 14, no. 8: 1020. https://doi.org/10.3390/biom14081020
APA StyleMilosevic, A., Milosevic, K., Zivkovic, A., Lavrnja, I., Savic, D., Bjelobaba, I., & Janjic, M. M. (2024). Alterations in the Hypothalamic–Pituitary–Adrenal Axis as a Response to Experimental Autoimmune Encephalomyelitis in Dark Agouti Rats of Both Sexes. Biomolecules, 14(8), 1020. https://doi.org/10.3390/biom14081020