

Inhibitive Mechanism of Loquat Flower Isolate on Tyrosinase Activity and Melanin Synthesis in Mouse Melanoma B16 Cells
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Preparation of LFP Sample
2.3. Identification of Constituents in LFP
2.4. Tyrosinase Inhibitory Activity and Kinetic Type Assays
2.5. Determination of Copper Ion Chelating Ability of LFP
2.6. Analysis of the Secondary Structure of Tyrosinase
2.7. Analysis of the Conformation Change in Tyrosinase
2.8. Cell Viability Assay
2.9. Determination of Tyrosinase Activity and Melanin Content in B16 Cells
2.10. Western Blotting Analysis
2.11. Statistical Analysis
3. Results and Discussion
3.1. Constituents of LFP
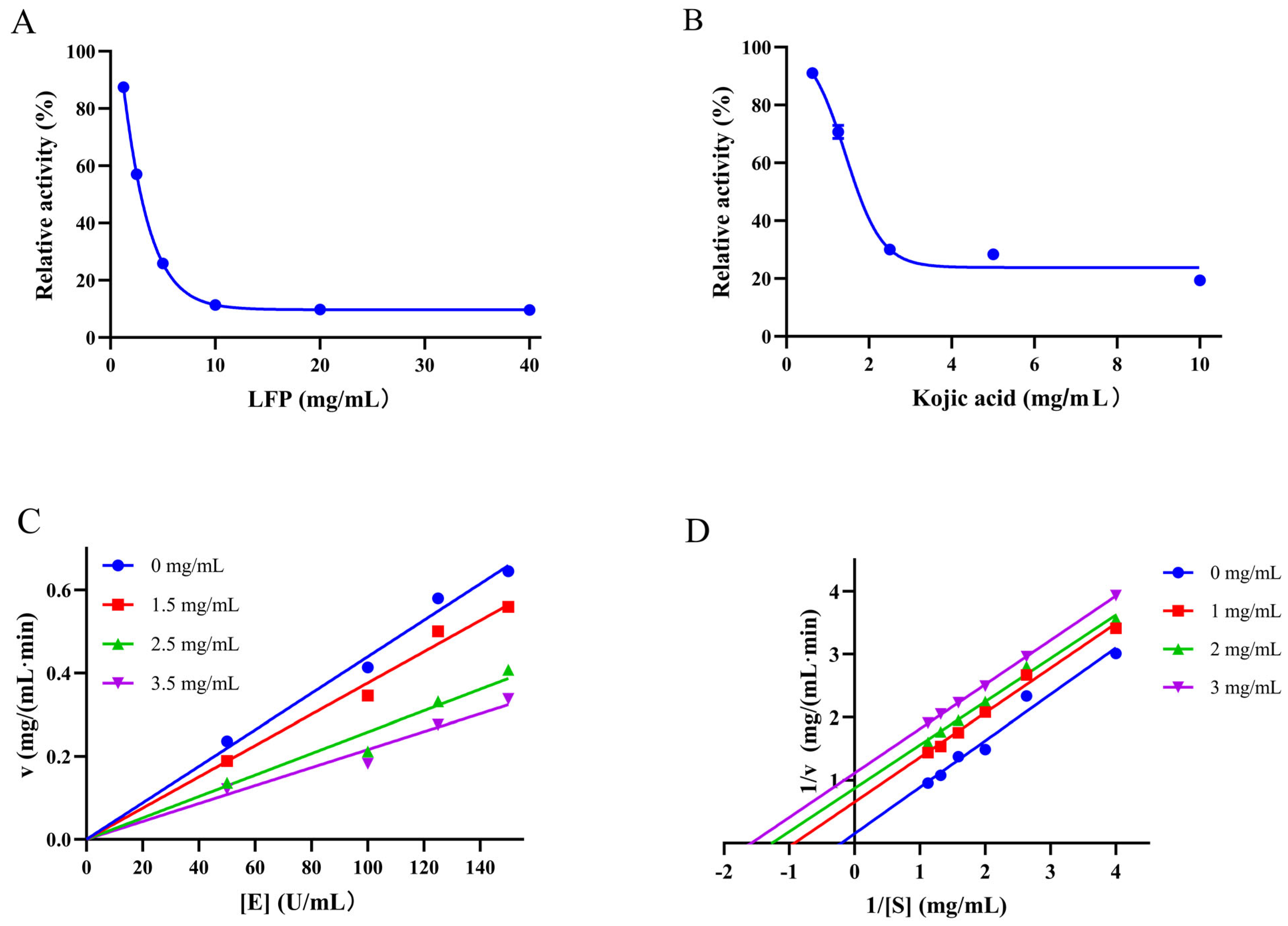
3.2. Inhibition Type of LFP on Tyrosinase Activity
3.3. Copper-Ion Chelating Ability of LFP
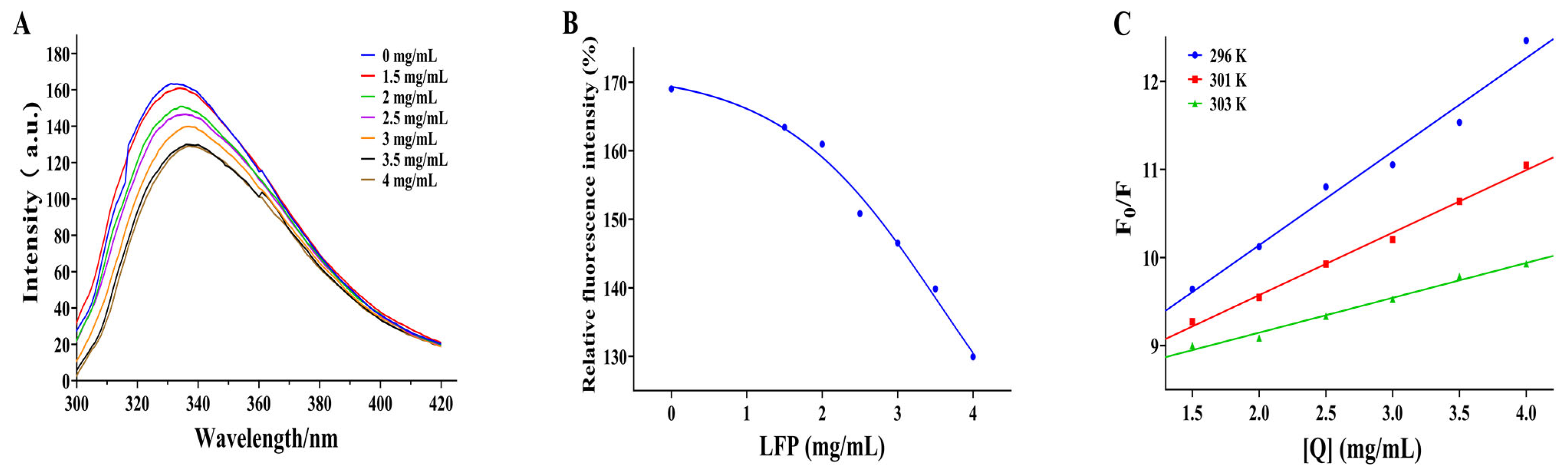
3.4. Effect of LFP on Secondary Structure of Tyrosinase
3.5. Effect of LFP on Conformation of Tyrosinase
3.6. Effect of LFP on the Viability of Mouse Melanoma B16 Cells
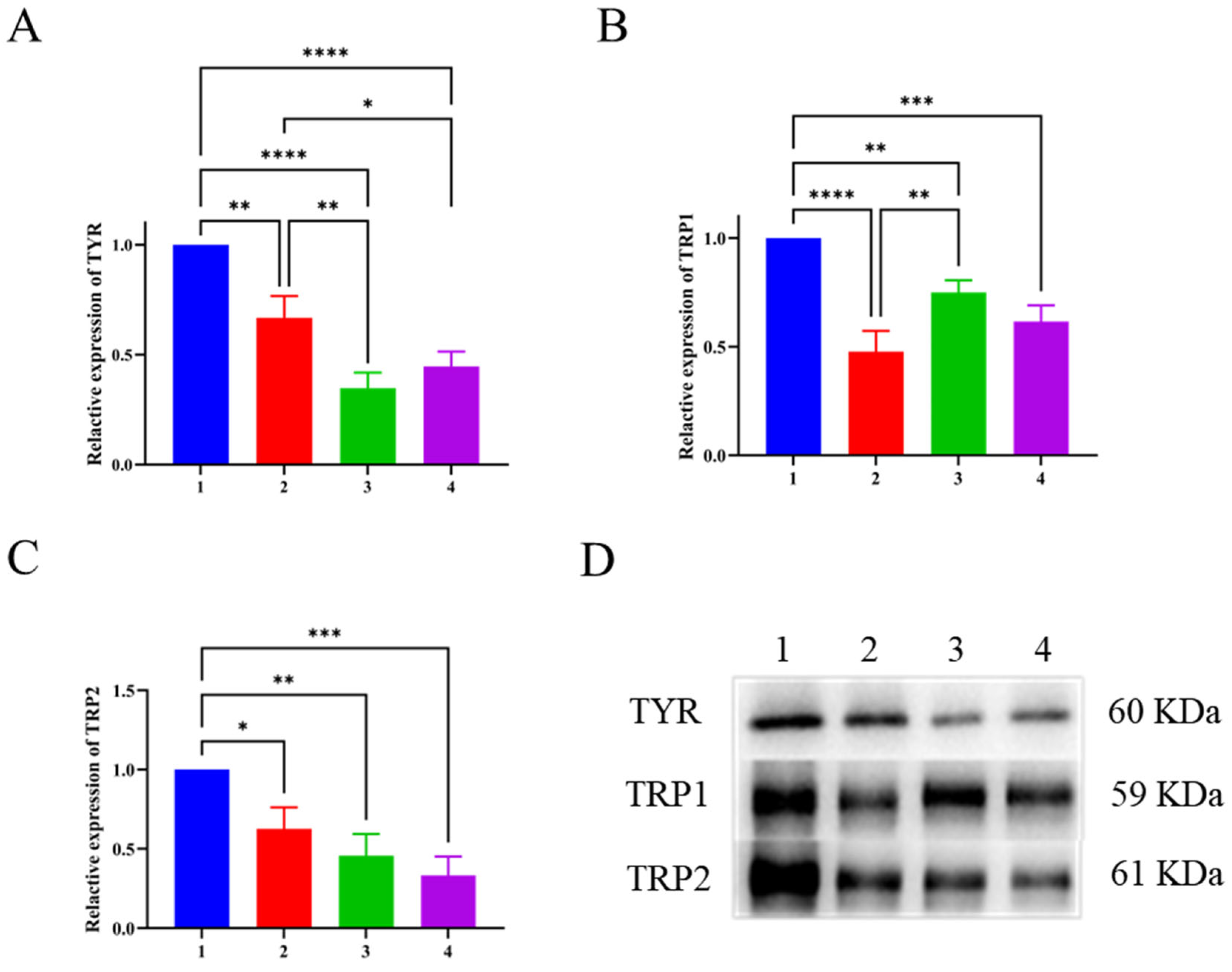
3.7. Effect of LFP on Tyrosinase Activity and Melanin Synthesis in B16 Cells
3.8. Western Blotting Results
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Wu, S.; Zhang, N.; Shen, X.; Mei, W.; He, Y.; Ge, W. Preparation of total flavonoids from loquat flower and its protective effect on acute alcohol-induced liver injury in mice. J. Food Drug Anal. 2015, 23, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Chen, J.; Zhu, J.; Yu, H. Changes of bioactive composition and concentration in loquat flower extracted with water/Chinese Baijiu. Heliyon 2023, 9, 14701. [Google Scholar] [CrossRef]
- Khouya, T.; Ramchoun, M.; Elbouny, H.; Hmidani, A.; Bouhlali, E.D.T.; Alem, C. Loquat (Eriobotrya japonica (Thunb) Lindl.): Evaluation of nutritional value, polyphenol composition, antidiabetic effect, and toxicity of leaf aqueous extract. J. Ethnopharmacol. 2022, 296, 115473. [Google Scholar] [CrossRef]
- Mokhtari, I.; Moumou, M.; Harnafi, M.; Milenkovic, D.; Amrani, S.; Harnafi, H. Loquat fruit peel extract regulates lipid metabolism and liver oxidative stress in mice: In vivo and in silico approaches. J. Ethnopharmacol. 2023, 310, 116376. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Xia, Z.; Deng, N.; Chen, L.; Zhang, H.; Lu, Y.; Liu, Y.; Gao, H. Eumelanin-inspired nanomaterials in electrochemical energy storage devices: A review. Chem. Eng. J. 2023, 452, 138607. [Google Scholar] [CrossRef]
- Flori, E.; Mastrofrancesco, A.; Mosca, S.; Ottaviani, M.; Briganti, S.; Cardinali, G.; Filoni, A.; Cameli, N.; Zaccarini, M.; Zouboulis, C.C.; et al. Sebocytes contribute to melasma onset. iScience 2022, 25, 10387. [Google Scholar] [CrossRef]
- Chen, Q.; Shang, C.; Han, M.; Chen, C.; Tang, W.; Liu, W. Inhibitory mechanism of scutellarein on tyrosinase by kinetics, spectroscopy and molecular simulation. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2023, 296, 122644. [Google Scholar] [CrossRef] [PubMed]
- Wakamatsu, K.; Zippin, J.H.; Ito, S. Chemical and biochemical control of skin pigmentation with special emphasis on mixed melanogenesis. Pigment. Cell Melanoma Res. 2021, 34, 730–747. [Google Scholar] [CrossRef]
- Lee, J.; Jeong, Y.; Jin Jung, H.; Ullah, S.; Ko, J.; Young Kim, G.; Yoon, D.; Hong, S.; Kang, D.; Park, Y.; et al. Anti-tyrosinase flavone derivatives and their anti-melanogenic activities: Importance of the beta-phenyl-alpha, beta-unsaturated carbonyl scaffold. Bioorg. Chem. 2023, 135, 106504. [Google Scholar] [CrossRef]
- Yu, Q.; Fan, L.; Ding, Z. The inhibition mechanisms between asparagus polyphenols after hydrothermal treatment and tyrosinase: A circular dichroism spectrum, fluorescence, and molecular docking study. Food Biosci. 2022, 48, 101790. [Google Scholar] [CrossRef]
- Ramadhan, R.; Maharani, R.; Devi, A.P.; Warnida, H.; Fatriasari, W. Biomass as Whitening Agents Derived from Plants. In Biomass-Based Cosmetics; Arung, E.T., Kusuma, W.F.I.W., Shimizu, H.K.K., Azelee, N.I.W., Edis, Z., Eds.; Springer: Singapore, 2024. [Google Scholar] [CrossRef]
- Feng, D.; Fang, Z.; Zhang, P. The melanin inhibitory effect of plants and phytochemicals: A systematic review. Phytomedicine 2022, 107, 154449. [Google Scholar] [CrossRef] [PubMed]
- Ha, A.C.; Le, T.M. Improvement of anti-tyrosinase activity in potential skin whitening products by combining herbal ex-tracts and reducing their tannin content by collagen fibre adsorption. S. Afr. J. Bot. 2023, 155, 118–126. [Google Scholar] [CrossRef]
- Liu, L.; Li, J.; Zhang, L.; Wei, S.; Qin, Z.; Liang, D.; Ding, B.; Chen, H.; Song, W. Conformational changes of tyrosinase caused by pentagalloylglucose binding: Implications for inhibitory effect and underlying mechanism. Food Res. Int. 2022, 157, 111312. [Google Scholar] [CrossRef] [PubMed]
- Ju, X.; Cheng, S.; Li, H.; Xu, X.; Wang, Z.; Du, M. Tyrosinase inhibitory effects of the peptides from fish scale with the metal copper ions chelating ability. Food Chem. 2022, 390, 133146. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Zeng, H.; Xia, F.; Ji, Q.; Xue, J.; Ren, R.; Que, F.; Zhou, B. The dietary flavonoid isoliquiritigenin induced apopto-sis and suppressed metastasis in melanoma cells: An in vitro and in vivo study. Life Sci. 2021, 264, 118598. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Noh, S.G.; Park, Y.; Kang, D.; Chun, P.; Chung, H.Y.; Jung, H.J.; Moon, H.R. A potent tyrosinase inhibitor, (E)-3-(2, 4-Dihydroxyphenyl)-1-(thiophen-2-yl) prop-2-en-1-one, with anti-melanogenesis properties in α-MSH and IBMX-induced B16F10 melanoma cells. Molecules 2018, 23, 2725. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Fan, L.; Duan, Z. Five individual polyphenols as tyrosinase inhibitors: Inhibitory activity, synergistic effect, action mechanism, and molecular docking. Food Chem. 2019, 297, 124910. [Google Scholar] [CrossRef]
- Gong, G.; Guan, Y.Y.; Zhang, Z.L.; Rahman, K.; Wang, S.J.; Zhou, S.; Luan, X.; Zhang, H. Isorhamnetin: A review of pharmacological effects. Biomed. Pharmacother. 2020, 128, 110301. [Google Scholar] [CrossRef]
- Imen, M.B.; Chaabane, F.; Nadia, M.; Soumaya, K.J.; Kamel, G.; Leila, C.G. Anti-melanogenesis and antigenotoxic activities of eriodictyol in murine melanoma (B16-F10) and primary human keratinocyte cells. Life Sci. 2015, 135, 173–178. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, M.; Zhu, X.; Liu, R.; Lu, Q. Metabolomics reveals that phenolamides are the main chemical components contributing to the anti-tyrosinase activity of bee pollen. Food Chem. 2022, 389, 133071. [Google Scholar] [CrossRef]
- Shi, Y.; Chen, Q.X.; Wang, Q.; Song, K.K.; Qiu, L. Inhibitory effects of cinnamic acid and its derivatives on the diphenolase activity of mushroom (Agaricus bisporus) tyrosinase. Food Chem. 2005, 92, 707–712. [Google Scholar] [CrossRef]
- Varela, M.T.; Ferrarini, M.; Mercaldi, V.G.; Sufi, B.d.S.; Padovani, G.; Nazato, L.I.S.; Fernandes, J.P.S. Coumaric acid derivatives as tyrosinase inhibitors: Efficacy studies through in silico, in vitro and ex vivo approaches. Bioorg. Chem. 2020, 103, 104108. [Google Scholar] [CrossRef] [PubMed]
- Zheleva-Dimitrova, D.; Zengin, G.; Sinan, K.I.; Yıldıztugay, E.; Mahomoodally, M.F.; Ak, G.; Picot-Allain, M.C.N.; Gevrenova, R. Identification of bioactive compounds from Rhaponticoides iconiensis extracts and their bioactivities: An endemic plant to Turkey flora. J. Pharmaceut. Biomed. 2020, 190, 113537. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Fan, L. Understanding the combined effect and inhibition mechanism of 4-hydroxycinnamic acid and ferulic acid as tyrosinase inhibitors. Food Chem. 2021, 352, 129369. [Google Scholar] [CrossRef] [PubMed]
- Ghalla, H.; Issaoui, N.; Bardak, F.; Atac, A. Intermolecular interactions and molecular docking investigations on 4-methoxybenzaldehyde. Comp. Mater. Sci. 2018, 149, 291–300. [Google Scholar] [CrossRef]
- Satish, L.; Santra, S.; Tsurkan, M.V.; Werner, C.; Jana, M.; Sahoo, H. Conformational changes of GDNF-derived peptide induced by heparin, heparan sulfate, and sulfated hyaluronic acid—Analysis by circular dichroism spectroscopy and molecular dynamics simulation. Int. J. Biol. Macromol. 2021, 182, 2144–2150. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Qi, Z.; Liu, C. Inhibition mechanisms of humic acid and protein on the degradation of sulfamethazine by horse-radish peroxidase. Colloids Surf. A 2021, 629, 127473. [Google Scholar] [CrossRef]
- Chen, W.; Shi, X.; Xu, W.; McClements, D.J.; Liu, X.; Liu, F. Effects of different polyphenols on the structure and properties of sodium caseinate films mediated by tyrosinase. J. Agr. Food Res. 2022, 10, 100395. [Google Scholar] [CrossRef]
- Buitrago, E.; Faure, C.; Carotti, M.; Bergantino, E.; Hardre, R.; Maresca, M.; Philouze, C.; Vanthuyne, N.; Boumendjel, A.; Bubacco, L.; et al. Exploiting HOPNO-dicopper center interaction to development of inhibitors for human tyrosinase. Eur. J. Med. Chem. 2023, 248, 115090. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, Y.; Wang, Y.; Xu, M.; Hu, X. UV–Vis spectroscopy combined with chemometric study on the interactions of three dietary flavonoids with copper ions. Food Chem. 2018, 263, 208–215. [Google Scholar] [CrossRef]
- Baltacioglu, H.; Bayindirli, A.; Severcan, M.; Severcan, F. Effect of thermal treatment on secondary structure and conformational change of mushroom polyphenol oxidase (PPO) as food quality related enzyme: A FTIR study. Food Chem. 2015, 187, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.; Hild, M.B.; Kussicke, A.; Kurre, A.; Lindinger, A. Fluorescence contrast improvement by polarization shaped laser pulses for autofluorescent biomolecules. Optik 2020, 207, 163777. [Google Scholar] [CrossRef]
- Sun, Q.; Guo, Y.; Li, X.; Luo, X.; Qiu, Y.; Liu, G. A tyrosinase fluorescent probe with large Stokes shift and high fluorescence enhancement for effective identification of liver cancer cells. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2023, 285, 121831. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Zhang, G.; Deng, S.; Huang, Z.; Peng, J.; Zhang, G.; Su, L.; He, W.; Wu, Y.; Ding, N.; et al. Synergistic dual-mechanism fluorescence quenching immunochromatographic assay based on Fe-polydopamine submicrobeads for sensitive detection of enrofloxacin. Chem. Eng. J. 2023, 454, 140444. [Google Scholar] [CrossRef]
- Jia, Y.; Yan, X.; Li, X.; Zhang, S.; Huang, Y.; Zhang, D.; Li, Y.; Qi, B. Soy protein-phlorizin conjugate prepared by tyrosinase catalysis: Identification of covalent binding sites and alterations in protein structure and functionality. Food Chem. 2023, 404 Pt A, 134610. [Google Scholar] [CrossRef]
- Deng, B.; Zhao, J.; He, M.; Tian, S. Curcumin treatment enhances bioactive metabolite accumulation and reduces enzymatic browning in soybean sprouts during storage. Food Chem. X 2023, 17, 100607. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, B.R.; Vitousek, M.N.; Safran, R.J. Signaling stress? An analysis of phaeomelanin-based plumage color and indi-vidual corticosterone levels at two temporal scales in North American barn swallows, Hirundo rustica erythrogaster. Horm. Behav. 2013, 64, 665–672. [Google Scholar] [CrossRef]
- Suzuki, T.; Hoshino, M.; Nishimura, M.; Ide, F.; Kusama, K.; Sakashita, H.; Kikuchi, K. A rare case of melanin-pigmented dentinogenic ghost cell tumor. J. Oral Maxil. Surg. Med. Pathol. 2023, 35, 335–340. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Compound Name (Literature) | Molecular Formula | Molecular Weight | CAS Number | Retention Time (min) |
|---|---|---|---|---|---|
| Flavonoids | |||||
| 1 | Quercetin [18] | C15H10O7 | 302. 042 0 | 117-39-5 | 6.643 |
| 2 | Isorhamnetin [19] | C16H12O7 | 316. 057 7 | 480-19-3 | 7.108 |
| 3 | Eriodictyol [20] | C15H12O6 | 288. 063 0 | 209-016-4 | 6.894 |
| Cinnamic acid and its derivatives | |||||
| 4 | Tricoumaroyl spermidine [21] | C34H37N3O6 | 583. 267 4 | NA | 9.236 |
| 5 | Dicoumaramide spermidine [21] | C25H31N3O4 | 437. 230 7 | NA | 9.225 |
| 6 | Ethyl caffeate [22] | C11H12O4 | 208. 073 2 | 102-37-4 | 9.294 |
| 7 | 2-Hydroxycinnamic acid [23] | C9H8O3 | 164. 047 2 | 614-60-8 | 5.88 |
| 8 | Coumaric acid [23] | C9H8O3 | 164. 047 2 | 614-60-8 | 10.271 |
| 9 | trans-4-Methoxycinnamic acid [24] | C10H10O3 | 178. 062 9 | 943-89-5 | 7.181 |
| 10 | Ferulic acid [25] | C10H10O4 | 194. 057 7 | 1 135-24-6 | 6.894 |
| Benzene and its derivatives | |||||
| 11 | 4-Methoxybenzaldehyde [26] | C8H8O2 | 136. 052 4 | 123-11-5 | 5.952 |
| 12 | p-Anisic acid (4-Methoxybenzoic acid) [26] | C8H8O3 | 152. 047 2 | 100-09-4 | 10.034 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Q.; Tao, W.; Wang, J.; Li, J.; Zheng, M.; Liu, Y.; Lu, S.; Fang, Z. Inhibitive Mechanism of Loquat Flower Isolate on Tyrosinase Activity and Melanin Synthesis in Mouse Melanoma B16 Cells. Biomolecules 2024, 14, 895. https://doi.org/10.3390/biom14080895
Chen Q, Tao W, Wang J, Li J, Zheng M, Liu Y, Lu S, Fang Z. Inhibitive Mechanism of Loquat Flower Isolate on Tyrosinase Activity and Melanin Synthesis in Mouse Melanoma B16 Cells. Biomolecules. 2024; 14(8):895. https://doi.org/10.3390/biom14080895
Chicago/Turabian StyleChen, Qianqian, Wenyang Tao, Jianfeng Wang, Jingrui Li, Meiyu Zheng, Yinying Liu, Shengmin Lu, and Zhongxiang Fang. 2024. "Inhibitive Mechanism of Loquat Flower Isolate on Tyrosinase Activity and Melanin Synthesis in Mouse Melanoma B16 Cells" Biomolecules 14, no. 8: 895. https://doi.org/10.3390/biom14080895
APA StyleChen, Q., Tao, W., Wang, J., Li, J., Zheng, M., Liu, Y., Lu, S., & Fang, Z. (2024). Inhibitive Mechanism of Loquat Flower Isolate on Tyrosinase Activity and Melanin Synthesis in Mouse Melanoma B16 Cells. Biomolecules, 14(8), 895. https://doi.org/10.3390/biom14080895