
Investigating the Impact of Estrogen Levels on Voiding Characteristics, Bladder Structure, and Related Proteins in a Mouse Model of Menopause-Induced Lower Urinary Tract Symptoms

Abstract

1. Introduction
2. Materials and Methods
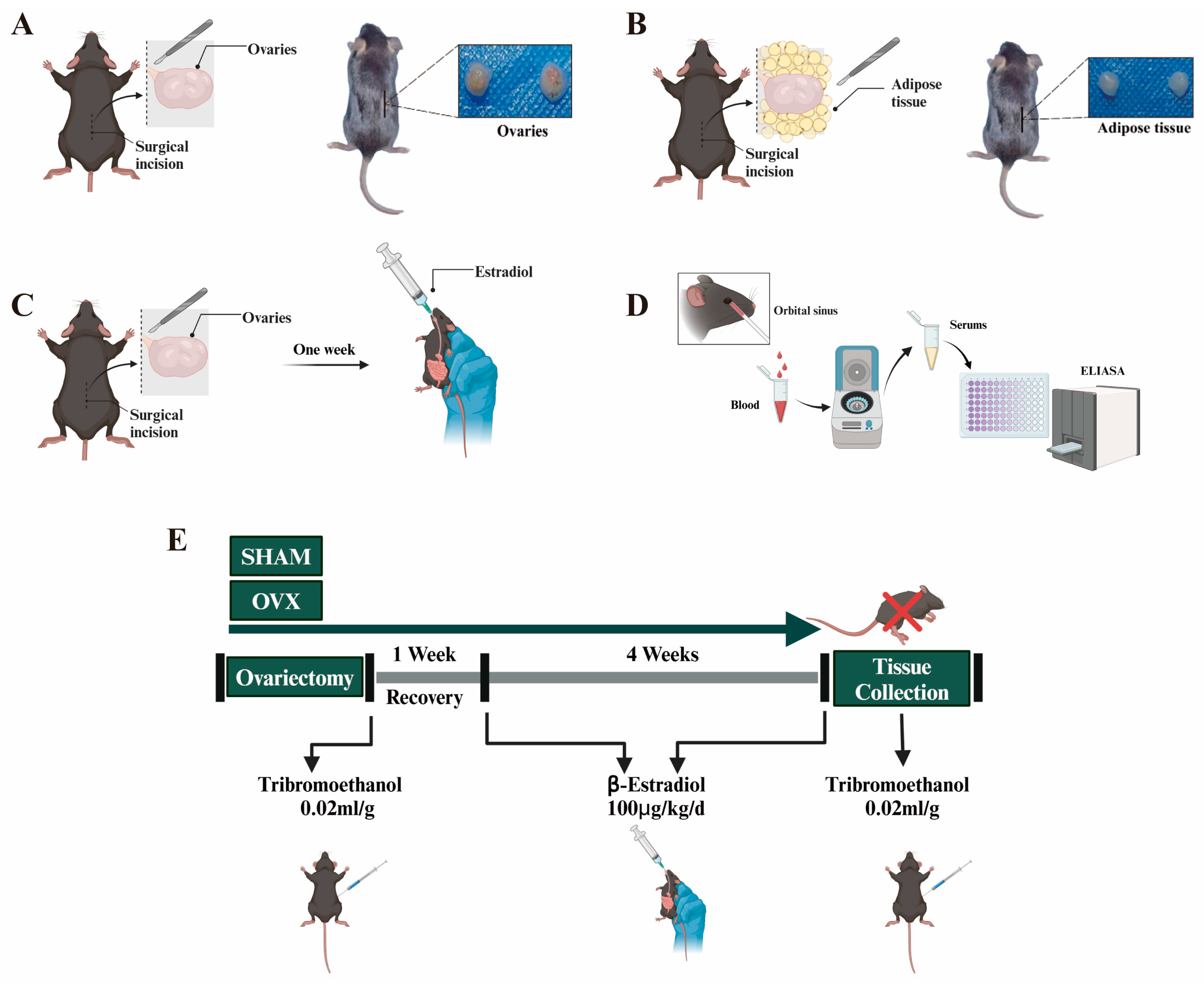
2.1. Experimental Design of Animal Model
2.2. Evaluation of Estradiol Level
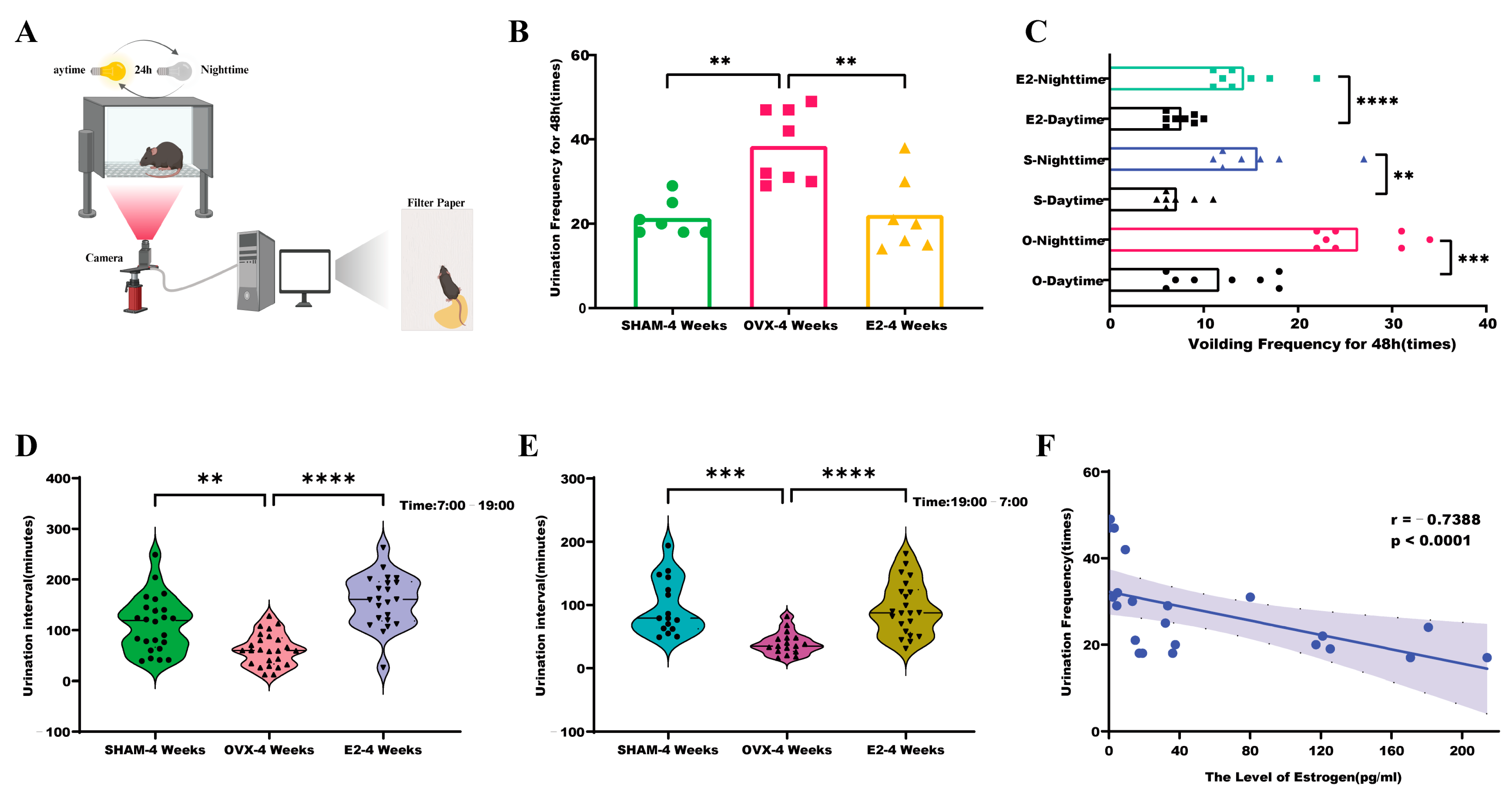
2.3. Statistics on the Frequency of Urination
2.4. Bladder Cystometry (Urodynamics)
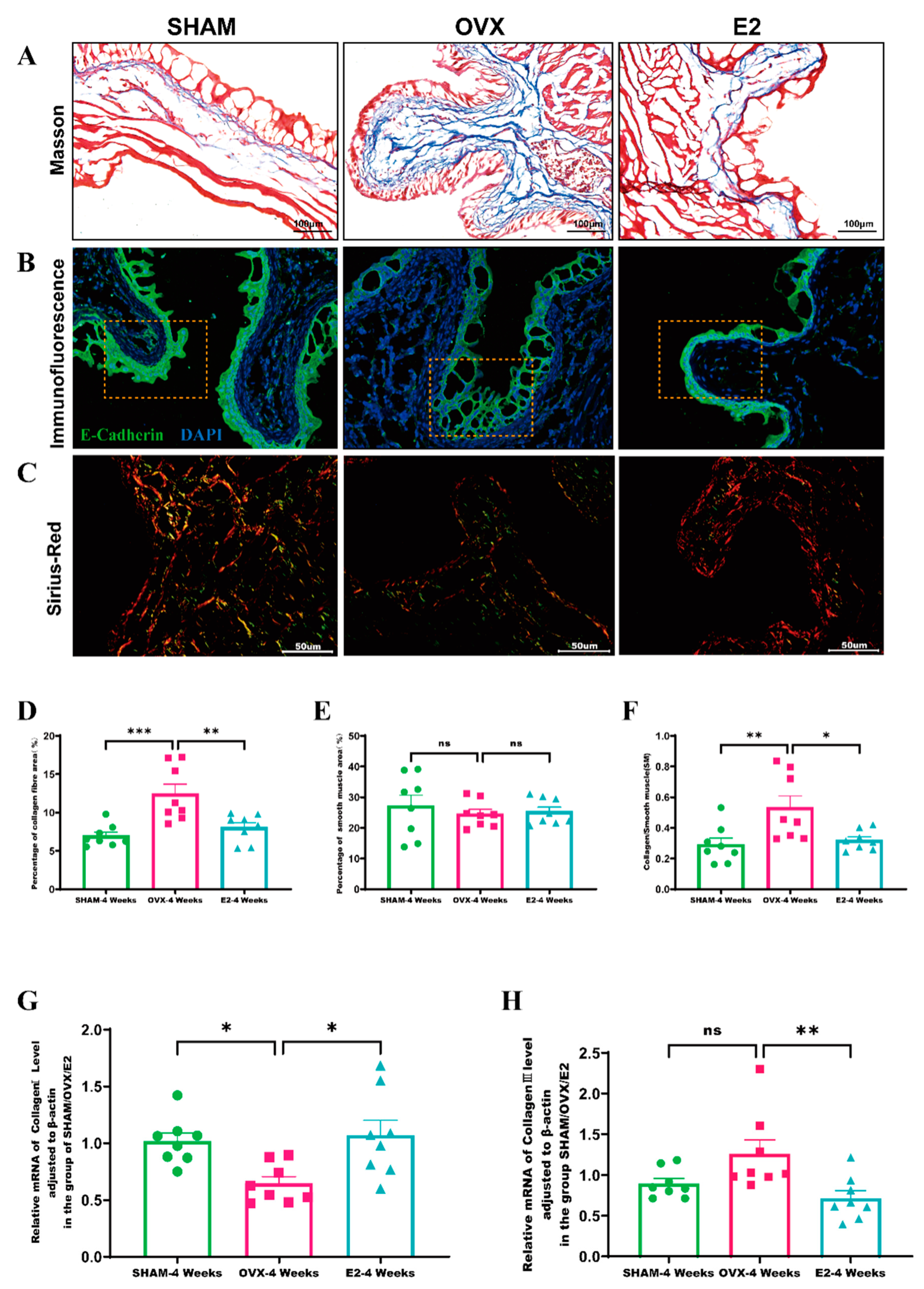
2.5. Masson Staining
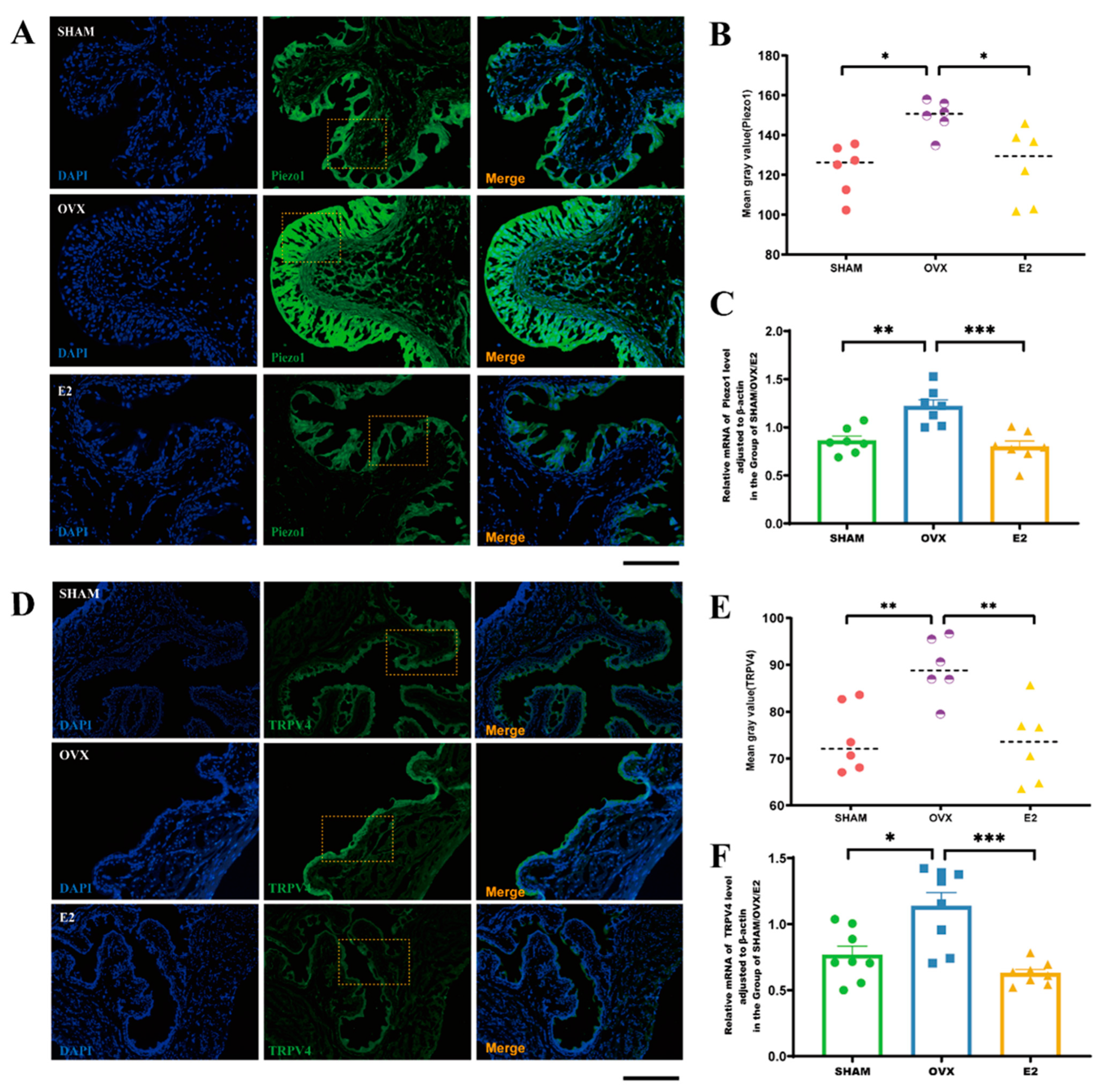
2.6. Immunofluorescence Staining
2.7. Quantitative Real-Time PCR Assay
2.8. Statistical Analysis
3. Results
3.1. Establishment and Validation of Animal Models
3.2. Urination Frequency in Mice Correlates with Estrogen Levels
3.3. Bladder Function Is Correlated with Estrogen Levels in Mice
3.4. Estrogen Intervention Decreases Collagen Deposition in Bladder Tissue
3.5. Estrogen Intervention Decreases Transcript Levels of Piezo1 and TRPV4 in Mice Bladder Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Przydacz, M.; Golabek, T.; Dudek, P.; Lipinski, M.; Chlosta, P. Prevalence and bother of lower urinary tract symptoms and overactive bladder in Poland, an Eastern European Study. Sci. Rep. 2020, 10, 12. [Google Scholar] [CrossRef]
- Liu, S.P.; Chuang, Y.C.; Sumarsono, B.; Chang, H.C. The prevalence and bother of lower urinary tract symptoms in men and women aged 40 years or over in Taiwan. J. Formos. Med. Assoc. 2019, 118, 170–178. [Google Scholar] [CrossRef]
- Stenberg, A.; Heimer, G.; Ulmsten, U. The prevalence of urogenital symptoms in postmenopausal women. Maturitas 1995, 22, S17–S20. [Google Scholar] [CrossRef]
- Christmas, M.M.; Iyer, S.; Daisy, C.; Maristany, S.; Letko, J.; Hickey, M. Menopause hormone therapy and urinary symptoms: A systematic review. Menopause-J. N. Am. Menopause Soc. 2023, 30, 672–685. [Google Scholar] [CrossRef]
- Russo, E.; Misasi, G.; Montt-Guevara, M.M.; Giannini, A.; Simoncini, T. Effects of ospemifene on overactive bladder in postmenopausal women with vulvovaginal atrophy. Climacteric 2023, 26, 284–288. [Google Scholar] [CrossRef]
- Pauwaert, K.; Goessaert, A.-S.; Ghijselings, L.; Bower, W.; Mariman, A.; Vogelaers, D.; Depypere, H.; Everaert, K. Hormone therapy as a possible solution for postmenopausal women with nocturia: Results of a pilot trial. Menopause-J. N. Am. Menopause Soc. 2021, 28, 502–510. [Google Scholar] [CrossRef]
- Robinson, D.; Cardozo, T.H.L. The effect of hormones on the lower urinary tract. Menopause Int. 2013, 19, 155–162. [Google Scholar] [CrossRef]
- Robinson, D.; Cardozo, L. Estrogens and the Lower Urinary Tract. Neurourol. Urodyn. 2011, 30, 754–757. [Google Scholar] [CrossRef]
- Palmieri, K.; Mannikarottu, A.S.; Chichester, P.; Kogan, B.; Leggett, R.E.; Whitbeck, C.; Levin, R.M. The effects of cyclical oestrogen on bladder and urethral structure and function. BJU Int. 2007, 99, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Dalghi, M.G.; Clayton, D.R.; Ruiz, W.G.; Al-Bataineh, M.M.; Satlin, L.M.; Kleyman, T.R.; Ricke, W.A.; Carattino, M.D.; Apodaca, G. Expression and distribution of PIEZO1 in the mouse urinary tract. Am. J. Physiol.-Renal Physiol. 2019, 317, F303–F321. [Google Scholar] [CrossRef]
- Dalghi, M.G.; Ruiz, W.G.; Clayton, D.R.; Montalbetti, N.; Daugherty, S.L.; Beckel, J.M.; Carattino, M.D.; Apodaca, G. Functional roles for PIEZO1 and PIEZO2 in urothelial mechanotransduction and lower urinary tract interoception. JCI Insight. 2021, 6, 19. [Google Scholar] [CrossRef]
- Miyamoto, T.; Mochizuki, T.; Nakagomi, H.; Kira, S.; Watanabe, M.; Takayama, Y.; Suzuki, Y.; Koizumi, S.; Takeda, M.; Tominaga, M. Functional Role for Piezo1 in Stretch-evoked Ca2+ Influx and ATP Release in Urothelial Cell Cultures. J. Biol. Chem. 2014, 289, 16565–16575. [Google Scholar] [CrossRef]
- Beča, K.I.K.; Girard, B.M.; Heppner, T.J.; Hennig, G.W.; Herrera, G.M.; Nelson, M.T.; Vizzard, M.A. The Role of PIEZO1 in Urinary Bladder Function and Dysfunction in a Rodent Model of Cyclophosphamide-Induced Cystitis. Front. Pain Res. 2021, 2, 748385. [Google Scholar] [CrossRef] [PubMed]
- Lai, A.; Cox, C.D.; Sekar, N.C.; Thurgood, P.; Jaworowski, A.; Peter, K.; Baratchi, S. Mechanosensing by Piezo1 and its implications for physiology and various pathologies. Biol. Rev. 2022, 97, 604–614. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Y.; Qi, J.; Wu, C.H.; Rong, W.F. Emerging roles of the TRPV4 channel in bladder physiology and dysfunction. J. Physiol.-Lond. 2021, 599, 39–47. [Google Scholar] [CrossRef]
- Janssen, D.A.W.; Hoenderop, J.G.; Jansen, K.; Kemp, A.W.; Heesakkers, J.; Schalken, J.A. The Mechanoreceptor TRPV4 is Localized in Adherence Junctions of the Human Bladder Urothelium: A Morphological Study. J. Urol. 2011, 186, 1121–1127. [Google Scholar] [CrossRef]
- Gevaert, T.; Vriens, J.; Segal, A.; Everaerts, W.; Roskams, T.; Talavera, K.; Owsianik, G.; Liedtke, W.; Daelemans, D.; Dewachter, I.; et al. Deletion of the transient receptor potential cation channel TRPV4 impairs murine bladder voiding. J. Clin. Investig. 2007, 117, 3453–3462. [Google Scholar] [CrossRef] [PubMed]
- Deruyver, Y.; Weyne, E.; Dewulf, K.; Rietjens, R.; Pinto, S.; Van Ranst, N.; Franken, J.; Vanneste, M.; Albersen, M.; Gevaert, T.; et al. Intravesical Activation of the Cation Channel TRPV4 Improves Bladder Function in a Rat Model for Detrusor Underactivity. Eur. Urol. 2018, 74, 336–345. [Google Scholar] [CrossRef]
- Merrill, L.; Vizzard, M.A. Intravesical TRPV4 blockade reduces repeated variate stress-induced bladder dysfunction by increasing bladder capacity and decreasing voiding frequency in male rats. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2014, 307, R471–R480. [Google Scholar] [CrossRef]
- Zhang, K.; Yang, Q.; Yang, L.; Li, Y.-J.; Wang, X.-S.; Dang, R.-L.; Guan, S.-Y.; Guo, Y.-Y.; Sun, T.; Wu, Y.-M.; et al. CB1 agonism prolongs therapeutic window for hormone replacement in ovariectomized mice. J. Clin. Invest. 2019, 129, 2333–2350. [Google Scholar] [CrossRef]
- Li, C.; Wang, B.; Lu, X.; Huang, Y.; Wang, H.; Xu, D.; Zhang, J. Maternal exposure to cadmium from puberty through lactation induces abnormal reproductive development in female offspring. Ecotox. Environ. Safe 2022, 242, 10. [Google Scholar] [CrossRef]
- Hong, Y.; Xu, W.-Q.; Feng, J.; Lou, H.; Liu, H.; Wang, L.; Cui, H.; Jiang, L.-T.; Xu, R.-C.; Xu, H.-H.; et al. Nitidine chloride induces cardiac hypertrophy in mice by targeting autophagy-related 4B cysteine peptidase. Acta Pharmacol. Sin. 2023, 44, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Gan, D.; Zhang, W.; Huang, C.; Chen, J.; He, W.; Wang, A.; Li, B.; Zhu, X. Ursolic acid ameliorates CCl4-induced liver fibrosis through the NOXs/ROS pathway. J. Cell Physiol. 2018, 233, 6799–6813. [Google Scholar] [CrossRef] [PubMed]
- Munoz, A.; Somogyi, G.T.; Boone, T.B.; Ford, A.P.; Smith, C.P. Modulation of bladder afferent signals in normal and spinal cord-injured rats by purinergic P2X3 and P2X2/3 receptors. BJU Int. 2012, 110, E409–E414. [Google Scholar] [CrossRef]
- Deer, R.R.; Stallone, J.N. Effects of estrogen on cerebrovascular function: Age-dependent shifts from beneficial to detrimental in small cerebral arteries of the rat. Am. J. Physiol.-Heart Circul. Physiol. 2016, 310, H1285–H1294. [Google Scholar] [CrossRef]
- Deer, R.R.; Stallone, J.N.; Pabbidi, M.R.; Kuppusamy, M.; Didion, S.P.; Sanapureddy, P.; Reed, J.T.; Sontakke, S.P.; Toth, P.; Tarantini, S.; et al. Hormonal gain control of a medial preoptic area social reward circuit. Nat. Neurosci. 2017, 20, 449–458. [Google Scholar]
- Piotrowska, K.; Tarnowski, M.; Zgutka, K.; Pawlik, A. Gender Differences in Response to Prolonged Every-Other-Day Feeding on the Proliferation and Apoptosis of Hepatocytes in Mice. Nutrients 2016, 8, 13. [Google Scholar] [CrossRef]
- Yoshida, J.; Aikawa, K.; Yoshimura, Y.; Shishido, K.; Yanagida, T.; Yamaguchi, O. The effects of ovariectomy and estrogen replacement on acetylcholine release from nerve fibres and passive stretch-induced acetylcholine release in female rat bladder. Neurourol. Urodyn. 2007, 26, 1050–1055. [Google Scholar] [CrossRef]
- Kullmann, F.A.; Limberg, B.J.; Artim, D.E.; Shah, M.; Downs, T.R.; Contract, D.; Wos, J.; Rosenbaum, J.S.; de Groat, W.C. Effects of β3-Adrenergic Receptor Activation on Rat Urinary Bladder Hyperactivity Induced by Ovariectomy. J. Pharmacol. Exp. Ther. 2009, 330, 704–717. [Google Scholar] [CrossRef]
- Liang, W.; Afshar, K.; Stothers, L.; Laher, I. The influence of ovariectomy and estrogen replacement on voiding patterns and detrusor muscarinic receptor affinity in the rat. Life Sci. 2002, 71, 351–362. [Google Scholar] [CrossRef]
- Diep, N.; Constantinou, C.E. Age dependent response to exogenous estrogen on micturition, contractility and cholinergic receptors of the rat bladder. Life Sci. 1999, 64, PL279–PL289. [Google Scholar] [CrossRef]
- Hextall, A. The Effect of Oestrogen and the Menopause on the Female Lower Urinary Tract. PhD Thesis, University of London, London, UK, 2002. [Google Scholar]
- Long, C.Y.; Liu, C.M.; Hsu, S.C.; Wu, C.H.; Wang, C.L.; Tsai, E.M. A randomized comparative study of the effects of oral and topical estrogen therapy on the vaginal vascularization and sexual function in hysterectomized postmenopausal women. Menopause-J. N. Am. Menopause Soc. 2006, 13, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann, N.; Christ, G.; Sclafani, T.; Melman, A. The effect of ovariectomy and long-term estrogen replacement on bladder structure and function in the rat. J. Urol. 2002, 168, 1265–1268. [Google Scholar] [CrossRef] [PubMed]
- Rodrìguez, L.V.; Wang, B.Y.; Shortliffe, L.M.D. Structural changes in the bladder walls of pregnant and hormone-treated rats: Correlation with bladder dynamics. BJU Int. 2004, 94, 1366–1372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Liao, L.M. Long-term follow-up of neurogenic bladder patients after bladder augmentation with small intestinal submucosa. World J. Urol. 2020, 38, 2279–2288. [Google Scholar] [CrossRef] [PubMed]
- Collado, A.; Batista, E.; Gelabert-Más, A.; Corominas, J.M.; Arañó, P.; Villavicencio, H. Detrusor quantitative morphometry in obstructed males and controls. J. Urol. 2006, 176, 2722–2728. [Google Scholar] [CrossRef]
- Anumanthan, G.; Tanaka, S.T.; Adams, C.M.; Thomas, J.C.; Wills, M.L.; Adams, M.C.; Hayward, S.W.; Matusik, R.J.; Bhowmick, N.A.; Brock, J.W.; et al. Bladder Stromal Loss of Transforming Growth Factor Receptor II Decreases Fibrosis After Bladder Obstruction. J. Urol. 2009, 182, 1775–1780. [Google Scholar] [CrossRef]
- Zhu, Q.; Ritchie, J.; Marouf, N.; Dion, S.B.; Resnick, N.M.; Elbadawi, A.; Elbadawi, A.; Kuchel, G.A. Role of ovarian hormones in the pathogenesis of impaired detrusor contractility: Evidence in ovariectomized rodents. J. Urol. 2001, 166, 1136–1141. [Google Scholar] [CrossRef]
- Dobberfuhl, A.D.; Schuler, C.; Leggett, R.E.; De, E.J.B.; Levin, R.M. Estrogen replacement is protective to the effect of in vitro hypoxia on female rabbit bladder and pelvic floor contractile response. Investig. Clin. Urol. 2020, 61, 432–440. [Google Scholar] [CrossRef]
- Lin, A.D.-Y.; Mannikarottu, A.; A Kogan, B.; Whitbeck, C.; Chichester, P.; E Leggett, R.; Levin, R.M. Estrogen induces angiogenesis of the female rabbit bladder. J. Endocrinol. 2006, 190, 241–246. [Google Scholar] [CrossRef]
- Parekh, M.H.; Chichester, P.; Lobel, R.W.; Aikawa, K.; Levin, R.M. Effects of castration on female rabbit bladder physiology and morphology. Urology 2004, 64, 1048–1051. [Google Scholar] [CrossRef] [PubMed]
- Hass, M.A.; Nichol, P.; Lee, L.; Levin, R.M. Estrogen modulates permeability and prostaglandin levels in the rabbit urinary bladder. Prostaglandins Leukot Essent Fat. Acids. 2009, 80, 125–129. [Google Scholar] [CrossRef]
- Lin, W.Y.; Radu, F.; Schuler, C.; Leggett, R.E.; Mannikarottu, A.; Levin, R.M. The effect of ovariectomy and oestrogen therapy on the free fatty acid content, endogenous lipase activity, and phospholipid content of the rabbit urinary bladder. BJU Int. 2008, 102, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Landau, E.H.; Jayanthi, V.R.; Churchill, B.M.; Shapiro, E.; Gilmour, R.F.; Khoury, A.E.; Macarak, E.J.; Mclorie, G.A.; Steckler, R.E.; Kogan, B.A. Loss of elasticity in dysfunctional bladders—Urodynamic and histochemical correlation. J. Urol. 1994, 152, 702–705. [Google Scholar] [CrossRef]
- Inaba, M.; Fushiki, S.; Yaoi, T.; Iwata, T.; Kamoi, K.; Okihara, K.; Ukimura, O.; Kawauchi, A.; Miyashita, H.; Kojima, M.; et al. Changes in extracellular matrix components of bladder detrusor in relation to bladder hypertrophy and compliance in patients with benign prostatic hyperplasia. Acta Histochem. Cytochem. 2000, 33, 131–139. [Google Scholar] [CrossRef]
- Zhang, W.; Li, Y.; Xi, X.; Zhu, G.; Wang, S.; Liu, Y.; Song, M. MicroRNA-15a-5p induces pulmonary artery smooth muscle cell apoptosis in a pulmonary arterial hypertension model via the VEGF/p38/MMP-2 signaling pathway. Int. J. Mol. Med. 2020, 45, 461–474. [Google Scholar] [CrossRef]
- Trabucco, E.; Soderberg, M.; Cobellis, L.; Torella, M.; Bystrom, B.; Ekman-Ordeberg, G.; Petraglia, F.; Colacurci, N. Role of proteoglycans in the organization of periurethral connective tissue in women with stress urinary incontinence. Maturitas 2007, 58, 395–405. [Google Scholar] [CrossRef]
- Kobata, S.; Girão, M.; Baracat, E.; Kajikawa, M.; Di Bella, V.; Sartori, M.; Bella, Z.J.-D. Estrogen therapy influence on periurethral vessels in postmenopausal incontinent women using Dopplervelocimetry analysis. Maturitas 2008, 61, 243–247. [Google Scholar] [CrossRef]
- Aikawa, K.; Sugino, T.; Matsumoto, S.; Chichester, P.; Whitbeck, C.; Levin, R.M. The effect of ovariectomy and estradiol on rabbit bladder smooth muscle contraction and morphology. J. Urol. 2003, 170, 634–637. [Google Scholar] [CrossRef]
- Sánchez-Ortiz, R.F.; Wang, Z.; Menon, C.; DiSanto, M.E.; Wein, A.J.; Chacko, S. Estrogen modulates the expression of myosin heavy chain in detrusor smooth muscle. Am. J. Physiol.-Cell Physiol. 2001, 280, C433–C440. [Google Scholar] [CrossRef]
- Blacher, J.; Dabire, H.; Pomies, J.P.; Safar, M.E.; Stimpel, M. Long-term cardiovascular effects of high “osteoprotective” dose levels of 17β-estradiol in spontaneously hypertensive rats. Cardiovasc. Drugs Ther. 2000, 14, 303–307. [Google Scholar] [CrossRef]
- Naomi, R.; Ridzuan, P.M.; Bahari, H. Current Insights into Collagen Type I. Polymers 2021, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.; Ge, M.; Shen, S.; Yang, L.; Jin, T.; Cao, D.; Xu, H.; Zheng, X.; Qiu, S.; Wang, K.; et al. Activation of NFKB-JMJD3 signaling promotes bladder fibrosis via boosting bladder smooth muscle cell proliferation and collagen accumulation. Biochim. Biophys. Acta-Mol. Basis. Dis. 2019, 1865, 2403–2410. [Google Scholar] [CrossRef]
- He, Q.; Lin, Y.; Liao, B.; Zhou, L.; Ai, J.; Jin, X.; Li, H.; Wang, K. The role of interleukin-6/interleukin-6 receptor signaling in the mechanical stress-induced extracellular matrix remodeling of bladder smooth muscle. Arch. Biochem. Biophys. 2021, 702, 8. [Google Scholar] [CrossRef] [PubMed]
- Fakhrieh, M.; Darvish, M.; Ardeshirylajimi, A.; Taheri, M.; Omrani, M.D. Improved bladder smooth muscle cell differentiation of the mesenchymal stem cells when grown on electrospun polyacrylonitrile/polyethylene oxide nanofibrous scaffold. J. Cell Biochem. 2019, 120, 15814–15822. [Google Scholar] [CrossRef]
- Everaerts, W.; Vriens, J.; Owsianik, G.; Appendino, G.; Voets, T.; De Ridder, D.; Nilius, B. Functional characterization of transient receptor potential channels in mouse urothelial cells. Am. J. Physiol.-Renal. Physiol. 2010, 298, F692–F701. [Google Scholar] [CrossRef]
- Zhao, M.; Chen, Z.; Liu, L.; Ding, N.; Wen, J.; Liu, J.; Wang, W.; Ge, N.; Zu, S.; Song, W.; et al. Functional Expression of Transient Receptor Potential and Piezo1 Channels in Cultured Interstitial Cells of Human-Bladder Lamina Propria. Front. Physiol. 2022, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Ihara, T.; Mitsui, T.; Nakamura, Y.; Kanda, M.; Tsuchiya, S.; Kira, S.; Nakagomi, H.; Sawada, N.; Kamiyama, M.; Shigetomi, E.; et al. The time-dependent variation of ATP release in mouse primary-cultured urothelial cells is regulated by the clock gene. Neurourol. Urodyn. 2018, 37, 2535–2543. [Google Scholar] [CrossRef]
- Roberts, M.W.; Sui, G.; Wu, R.; Rong, W.; Wildman, S.; Montgomery, B.; Ali, A.; Langley, S.; Ruggieri Sr, M.R.; Wu, C. TRPV4 receptor as a functional sensory molecule in bladder urothelium: Stretch-independent, tissue-specific actions and pathological implications. Faseb. J. 2020, 34, 263–286. [Google Scholar] [CrossRef]
- Ihara, T.; Mitsui, T.; Nakamura, Y.; Kanda, M.; Tsuchiya, S.; Kira, S.; Nakagomi, H.; Sawada, N.; Kamiyama, M.; Hirayama, Y.; et al. The oscillation of intracellular Ca2+ influx associated with the circadian expression of Piezo1 and TRPV4 in the bladder urothelium. Sci. Rep. 2018, 8, 9. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′→3′) | Forward Primer (5′→3′) |
|---|---|---|
| Piezo1 | ATCGCCATCATCTGGTTCCC | AGCTCCAAGGTGTGCTTCTC |
| TRPV4 | TGAGCAGGCCGAGAAGTACA | AGTCCATCTAGGTCCGCAGT |
| Collagen I | ACGTAAGCACTGGTGGACAG | CAGGAGGGCCATAGCTGAAC |
| Collagen III | GAGGAATGGGTGGCTATCCG | TTGCGTCCATCAAAGCCTCT |
| β-actin | GCAGGAGTACGATGAGTCCG | AGTCCATCTAGGTCCGCAGT |
| Variable | SHAM | OVX | E2 |
|---|---|---|---|
| Serum estradiol level (pg/mL) | 35.20 ± 17.21 | 1.80 ± 1.32 **** | 38.18 ± 22.00 |
| Water intake (mL/48 h) | 9.50 ± 1.38 | 10.69 ± 3.42 | 8.00 ± 1.26 |
| Body weight (g) | 17.59 ± 0.58 | 20.22 ± 1.17 *** | 19.13 ± 1.99 |
| Bladder weight (mg) | 17.00 ± 1.60 | 18.00 ± 2.60 | 25.00 ± 2.20 ** |
| The ratio of bladder weight (mg)/body weight (g) | 0.89 ± 0.13 | 0.90 ± 0.15 | 1.1 ± 0.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Chen, Y.; Yin, L.; Deng, G.; Xia, X.; Tang, X.; Zhang, Y.; Yan, J. Investigating the Impact of Estrogen Levels on Voiding Characteristics, Bladder Structure, and Related Proteins in a Mouse Model of Menopause-Induced Lower Urinary Tract Symptoms. Biomolecules 2024, 14, 1044. https://doi.org/10.3390/biom14091044
Zhang C, Chen Y, Yin L, Deng G, Xia X, Tang X, Zhang Y, Yan J. Investigating the Impact of Estrogen Levels on Voiding Characteristics, Bladder Structure, and Related Proteins in a Mouse Model of Menopause-Induced Lower Urinary Tract Symptoms. Biomolecules. 2024; 14(9):1044. https://doi.org/10.3390/biom14091044
Chicago/Turabian StyleZhang, Chenglong, Yuangui Chen, Lingxuan Yin, Guoxian Deng, Xiaowen Xia, Xiaoshuang Tang, Yifeng Zhang, and Junan Yan. 2024. "Investigating the Impact of Estrogen Levels on Voiding Characteristics, Bladder Structure, and Related Proteins in a Mouse Model of Menopause-Induced Lower Urinary Tract Symptoms" Biomolecules 14, no. 9: 1044. https://doi.org/10.3390/biom14091044
APA StyleZhang, C., Chen, Y., Yin, L., Deng, G., Xia, X., Tang, X., Zhang, Y., & Yan, J. (2024). Investigating the Impact of Estrogen Levels on Voiding Characteristics, Bladder Structure, and Related Proteins in a Mouse Model of Menopause-Induced Lower Urinary Tract Symptoms. Biomolecules, 14(9), 1044. https://doi.org/10.3390/biom14091044