
Differential Expression of miR-223-3p and miR-26-5p According to Different Stages of Mastitis in Dairy Cows

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Milk Samples
2.3. Circulating microRNA Extraction and Reverse Transcription
2.4. Gene-Expression Analysis
2.5. Isolation of Bovine Lymphocytes, Monocytes, and Neutrophils
2.6. Inflammatory Stimulation of Bovine Immune-Cell Cultures
2.7. Statistical Analysis
3. Results
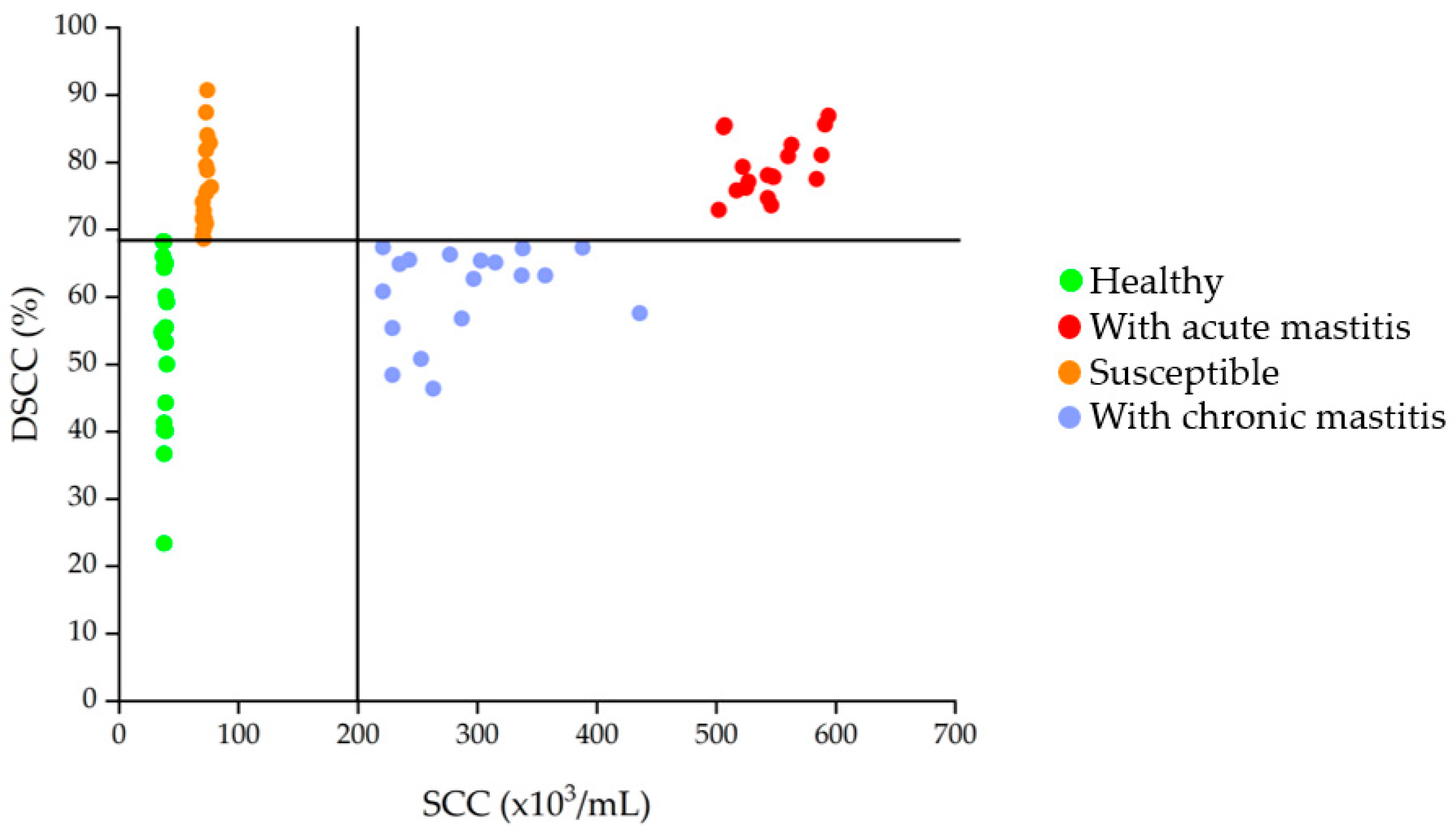
3.1. SCC and DSCC in Milk Samples
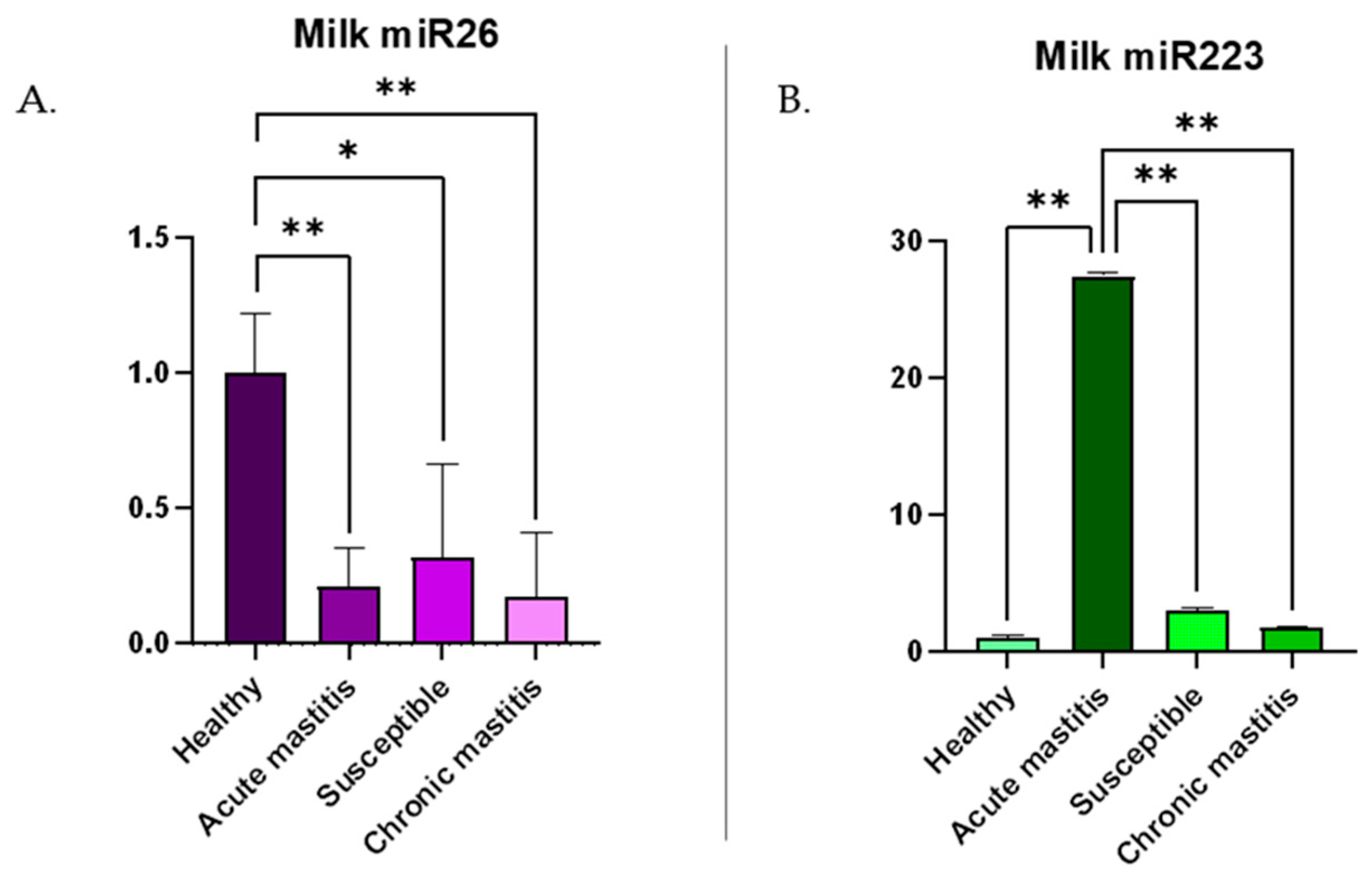
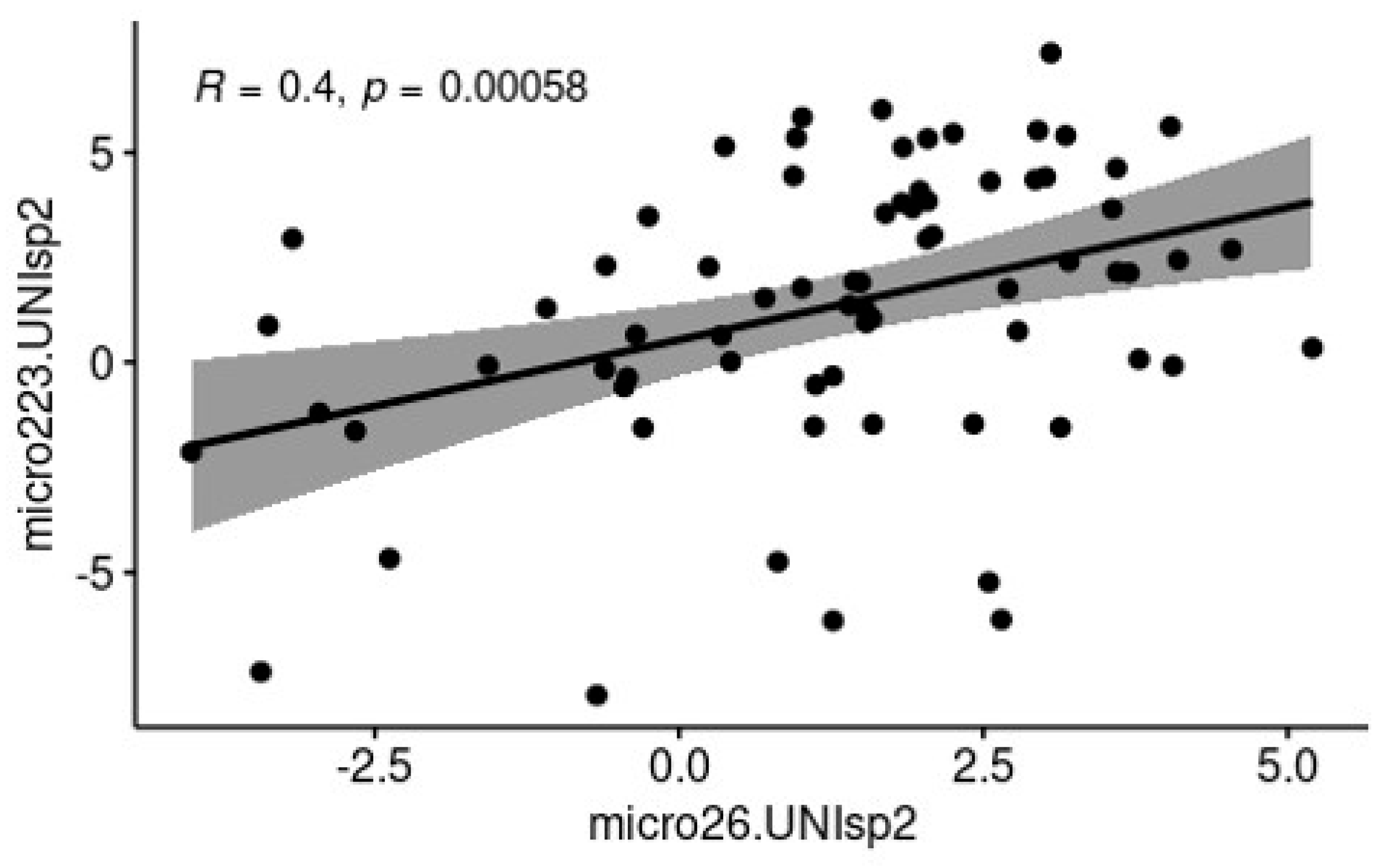
3.2. c-miRNA Expression in Milk
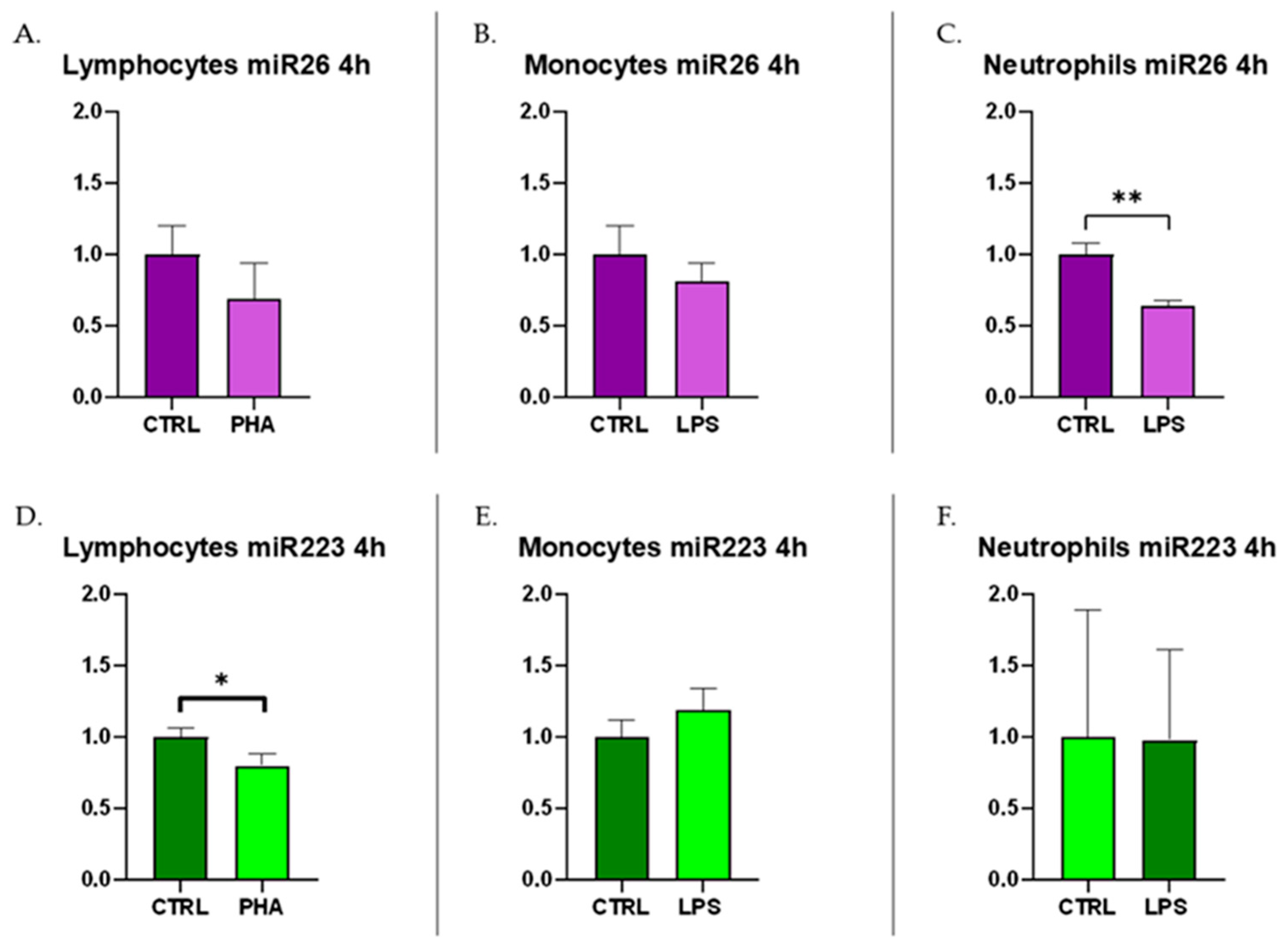
3.3. c-miRNA Expression in Immune Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huszenicza, G.; Jánosi, S.; Kulcsár, M.; Kóródi, P.; Reiczigel, J.; Kátai, L.; Peters, A.R.; De Rensis, F. Effects of clinical mastitis on ovarian function in post-partum dairy cows. Reprod. Domest. Anim. 2005, 40, 199–204. [Google Scholar] [CrossRef]
- Sharun, K.; Dhama, K.; Tiwari, R.; Gugjoo, M.B.; Iqbal Yatoo, M.; Patel, S.K.; Pathak, M.; Karthik, K.; Khurana, S.K.; Singh, R.; et al. Advances in therapeutic and managemental approaches of bovine mastitis: A comprehensive review. Vet. Q. 2021, 41, 107–136. [Google Scholar] [CrossRef]
- Zecconi, A.; Zaghen, F.; Meroni, G.; Sora, V.; Martino, P.A.; Laterza, G.; Zanini, L. Early Milk Total and Differential Cell Counts as a Diagnostic Tool to Improve Antimicrobial Therapy Protocols. Animals 2023, 13, 1143. [Google Scholar] [CrossRef]
- Piccinini, R.; Binda, E.; Belotti, M.; Casirani, G.; Zecconi, A. Comparison of blood and milk non-specific immune parameters in heifers after calving in relation to udder health. Vet. Res. 2005, 36, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Damm, M.; Holm, C.; Blaabjerg, M.; Bro, M.N.; Schwarz, D. Differential somatic cell count-A novel method for routine mastitis screening in the frame of Dairy Herd Improvement testing programs. J. Dairy Sci. 2017, 100, 4926–4940. [Google Scholar] [CrossRef]
- Zecconi, A.; Vairani, D.; Cipolla, M.; Rizzi, N.; Zanini, L. Assessment of subclinical mastitis diagnostic accuracy by differential cell count in individual cow milk. Ital. J. Anim. Sci. 2019, 18, 460–465. [Google Scholar] [CrossRef]
- Stocco, G.; Summer, A.; Cipolat-Gotet, C.; Zanini, L.; Vairani, D.; Dadousis, C.; Zecconi, A. Differential Somatic Cell Count as a Novel Indicator of Milk Quality in Dairy Cows. Animals 2020, 10, 753. [Google Scholar] [CrossRef]
- Stocco, G.; Cipolat-Gotet, C.; Stefanon, B.; Zecconi, A.; Francescutti, M.; Mountricha, M.; Summer, A. Herd and animal factors affect the variability of total and differential somatic cell count in bovine milk. J. Anim. Sci. 2023, 101, 406. [Google Scholar] [CrossRef]
- Arora, S.; Dev, K.; Agarwal, B.; Das, P.; Syed, M.A. Macrophages: Their role, activation and polarization in pulmonary diseases. Immunobiology 2018, 223, 383–396. [Google Scholar] [CrossRef]
- Halasa, T.; Kirkeby, C. Differential Somatic Cell Count: Value for Udder Health Management. Front. Vet. Sci. 2020, 7, 609055. [Google Scholar] [CrossRef]
- Ceciliani, F.; Ávila Morales, G.; De Matteis, G.; Grandoni, F.; Furioso Ferreira, R.; Roccabianca, P.; Lecchi, C. Methods in isolation and characterization of bovine monocytes and macrophages. Methods 2021, 186, 22–41. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef]
- Sarikaya, H.; Schlamberger, G.; Meyer, H.H.; Bruckmaier, R.M. Leukocyte populations and mRNA expression of inflammatory factors in quarter milk fractions at different somatic cell score levels in dairy cows. J. Dairy Sci. 2006, 89, 2479–2486. [Google Scholar] [CrossRef]
- Miretti, S.; Lecchi, C.; Ceciliani, F.; Baratta, M. MicroRNAs as Biomarkers for Animal Health and Welfare in Livestock. Front. Vet. Sci. 2020, 7, 578193. [Google Scholar] [CrossRef]
- Lai, Y.C.; Fujikawa, T.; Maemura, T.; Ando, T.; Kitahara, G.; Endo, Y.; Yamato, O.; Koiwa, M.; Kubota, C.; Miura, N. Inflammation-related microRNA expression level in the bovine milk is affected by mastitis. PLoS ONE 2017, 12, e0177182. [Google Scholar] [CrossRef] [PubMed]
- Luoreng, Z.M.; Wang, X.P.; Mei, C.G.; Zan, L.S. Comparison of microRNA Profiles between Bovine Mammary Glands Infected with Staphylococcus aureus and Escherichia coli. Int. J. Biol. Sci. 2018, 14, 87–99. [Google Scholar] [CrossRef]
- Srikok, S.; Patchanee, P.; Boonyayatra, S.; Chuammitri, P. Potential role of MicroRNA as a diagnostic tool in the detection of bovine mastitis. Prev. Vet. Med. 2020, 182, 105101. [Google Scholar] [CrossRef]
- Luoreng, Z.M.; Yang, J.; Wang, X.P.; Wei, D.W.; Zan, L.S. Expression Profiling of microRNA From Peripheral Blood of Dairy Cows in Response to Staphylococcus aureus-Infected Mastitis. Front. Vet. Sci. 2021, 8, 691196. [Google Scholar] [CrossRef]
- Cuccato, M.; Divari, S.; Giannuzzi, D.; Grange, C.; Moretti, R.; Rinaldi, A.; Leroux, C.; Sacchi, P.; Cannizzo, F.T. Extracellular vesicle miRNome during subclinical mastitis in dairy cows. Vet Res 2024, 55, 112. [Google Scholar] [CrossRef]
- Tzelos, T.; Ho, W.; Charmana, V.I.; Lee, S.; Donadeu, F.X. MiRNAs in milk can be used towards early prediction of mammary gland inflammation in cattle. Sci. Rep. 2022, 12, 5131. [Google Scholar] [CrossRef]
- ICAR (International Committee for Animal Recording). Section 2–Guidelines for Dairy Cows Milk Recording; ICAR: Utrecht, The Netherlands, 2023; Available online: www.icar.org (accessed on 3 February 2025).
- Ioannidis, J.; Risse, J.; Donadeu, F.X. Profiling of MicroRNAs in the Biofluids of Livestock Species. In MicroRNA Protocols. Methods in Molecular Biology; Ying, S.Y., Ed.; Humana Press: New York, NY, USA, 2018; Volume 1733. [Google Scholar]
- Schanzenbach, C.I.; Kirchner, B.; Ulbrich, S.E.; Pfaffl, M.W. Can milk cell or skim milk miRNAs be used as biomarkers for early pregnancy detection in cattle? PLoS ONE 2017, 12, e0172220. [Google Scholar] [CrossRef]
- Del Pozo-Acebo, L.; Hazas, M.L.L.; Tomé-Carneiro, J.; Gil-Cabrerizo, P.; San-Cristobal, R.; Busto, R.; García-Ruiz, A.; Dávalos, A. Bovine Milk-Derived Exosomes as a Drug Delivery Vehicle for miRNA-Based Therapy. Int. J. Mol. Sci. 2021, 22, 1105. [Google Scholar] [CrossRef]
- Schaefer, A.; Jung, M.; Miller, K.; Lein, M.; Kristiansen, G.; Erbersdobler, A.; Jung, K. Suitable reference genes for relative quantification of miRNA expression in prostate cancer. Exp. Mol. Med. 2010, 42, 749–758. [Google Scholar] [CrossRef]
- Ferrari, L.; Martelli, P.; Saleri, R.; De Angelis, E.; Cavalli, V.; Bresaola, M.; Benetti, M.; Borghetti, P. Lymphocyte activation as cytokine gene expression and secretion is related to the porcine reproductive and respiratory syndrome virus (PRRSV) isolate after in vitro homologous and heterologous recall of peripheral blood mononuclear cells (PBMC) from pigs vaccinated and exposed to natural infection. Vet. Immunol. Immunopathol. 2013, 151, 193–206. [Google Scholar] [PubMed]
- Kouoh, F.; Gressier, B.; Luyckx, M.; Brunet, C.; Dine, T.; Ballester, L.; Cazin, M.; Cazin, J.C. A simple method for isolating human and rabbit polymorphonuclear neutrophils (PMNs). Biol. Pharm. Bull. 2000, 23, 1382–1383. [Google Scholar] [CrossRef]
- Dagur, P.K.; McCoy, J.P. Collection, storage, and preparation of human blood cells. Curr. Protoc. Cytom. 2015, 73, 5-1. [Google Scholar] [CrossRef]
- Leroux, C.; Cuccato, M.; Pawłowski, K.; Cannizzo, F.T.; Sacchi, P.; Pires, J.A.A.; Faulconnier, Y. Milk fat miRNome changes in response to LPS challenge in Holstein cows. Vet Res 2023, 54, 111. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.; Gai, Y.; Liao, Q.; Zhang, L.; Li, Z.; Li, B.; Bai, M.; Li, N.; Deng, L. An Integrated Analysis of Lactation-Related miRNA and mRNA Expression Profiles in Donkey Mammary Glands. Genes 2022, 13, 1637. [Google Scholar] [CrossRef]
- He, X.; Cheng, X.; Ding, J.; Xiong, M.; Chen, B.; Cao, G. Hyperglycemia induces miR-26-5p down-regulation to overexpress PFKFB3 and accelerate epithelial-mesenchymal transition in gastric cancer. Bioengineered 2022, 13, 2902–2917. [Google Scholar] [CrossRef]
- Uttamani, J.R.; Naqvi, A.R.; Estepa, A.M.V.; Kulkarni, V.; Brambila, M.F.; Martínez, G.; Chapa, G.; Wu, C.D.; Li, W.; Rivas-Tumanyan, S.; et al. Downregulation of miRNA-26 in chronic periodontitis interferes with innate immune responses and cell migration by targeting phospholipase C beta 1. J. Clin. Periodontol. 2023, 50, 102–113. [Google Scholar] [CrossRef]
- Tzelos, T.; Lee, S.; Pegg, A.; Donadeu, F.X. Association between blood miR-26a levels following artificial insemination, and pregnancy outcome in dairy cattle. PLoS ONE 2023, 18, e0289342. [Google Scholar] [CrossRef]
- Di Mauro, S.; Filipe, J.; Facchin, A.; Roveri, L.; Addis, M.F.; Monistero, V.; Piccinini, R.; Sala, G.; Pravettoni, D.; Zamboni, C.; et al. The secretome of Staphylococcus aureus strains with opposite within-herd epidemiological behavior affects bovine mononuclear cell response. Vet. Res. 2023, 54, 120. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Gu, B.; Lu, W.; Liu, J.; Lu, J. miR-142-5p regulates lipopolysaccharide-induced bovine epithelial cell proliferation and apoptosis via targeting BAG5. Exp. Therap. Med. 2021, 22, 1425. [Google Scholar] [CrossRef] [PubMed]
- Hue, D.T.; Petrovski, K.; Chen, T.; Williams, J.L.; Bottema, C.D.K. Analysis of immune-related microRNAs in cows and newborn calves. J. Dairy Sci. 2023, 106, 2866–2878. [Google Scholar] [CrossRef]
- Sun, J.; Aswath, K.; Schroeder, S.G.; Lippolis, J.D.; Reinhardt, T.A.; Sonstegard, T.S. MicroRNA expression profiles of bovine milk exosomes in response to Staphylococcus aureus infection. BMC Genom. 2015, 16, 806. [Google Scholar] [CrossRef]
- Wang, X.P.; Luoreng, Z.M.; Zan, L.S.; Li, F.; Li, N. Bovine miR-146a regulates inflammatory cytokines of bovine mammary epithelial cells via targeting the TRAF6 gene. J. Dairy Sci. 2017, 100, 7648–7658. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.P.; Luoreng, Z.M.; Zan, L.S.; Raza, S.H.; Li, F.; Li, N.; Liu, S. Expression patterns of miR-146a and miR-146b in mastitis infected dairy cattle. MCP 2016, 30, 342–344. [Google Scholar] [CrossRef] [PubMed]
- Johnnidis, J.B.; Harris, M.H.; Wheeler, R.T.; Stehling-Sun, S.; Lam, M.H.; Kirak, O.; Brummelkamp, T.R.; Fleming, M.D.; Camargo, F.D. Regulation of progenitor cell proliferation and granulocyte function by microRNA-223. Nature 2008, 451, 1125–1129. [Google Scholar] [CrossRef]
- Li, L.; Huang, J.; Zhang, X.; Ju, Z.; Qi, C.; Zhang, Y.; Li, Q.; Wang, C.; Miao, W.; Zhong, J.; et al. One SNP in the 3’-UTR of HMGB1 gene affects the binding of target bta-miR-223 and is involved in mastitis in dairy cattle. Immunogenetics 2012, 64, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Jiao, P.; Wang, X.P.; Luoreng, Z.M.; Yang, J.; Jia, L.; Ma, Y.; Wei, D.W. miR-223: An Effective Regulator of Immune Cell Differentiation and Inflammation. Int. J. Biol. Sci. 2021, 17, 2308–2322. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Lu, K.; Ye, T.; Zhang, Z. MicroRNA 223 attenuates LPS induced inflammation in an acute lung injury model via the NLRP3 inflammasome and TLR4/NF κB signaling pathway via RHOB. Int. J. Mol. Med. 2019, 43, 1467–1477. [Google Scholar] [CrossRef]
- Valmiki, S.; Ahuja, V.; Puri, N.; Paul, J. miR-125b and miR-223 Contribute to Inflammation by Targeting the Key Molecules of NFκB Pathway. Front. Med. 2020, 6, 313. [Google Scholar] [CrossRef]
- Jiao, P.; Wang, J.; Yang, J.; Wang, X.; Luoreng, Z. Bta-miR-223 Targeting the RHOB Gene in Dairy Cows Attenuates LPS-Induced Inflammatory Responses in Mammary Epithelial Cells. Cells 2022, 11, 3144. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Barkema, H.W.; Gao, J.; Yang, J.; Wang, Y.; Kastelic, J.P.; Khan, S.; Liu, G.; Han, B. MicroRNA miR-223 modulates NLRP3 and Keap1, mitigating lipopolysaccharide-induced inflammation and oxidative stress in bovine mammary epithelial cells and murine mammary glands. Vet. Res. 2023, 54, 78. [Google Scholar] [CrossRef]
- Ifergan, I.; Chen, S.; Zhang, B.; Miller, S.D. Cutting Edge: MicroRNA-223 Regulates Myeloid Dendritic Cell-Driven Th17 Responses in Experimental Autoimmune Encephalomyelitis. J. Immunol. 2016, 196, 1455–1459. [Google Scholar] [CrossRef] [PubMed]
- Mohajeri, M.; Banach, M.; Atkin, S.L.; Butler, A.E.; Ruscica, M.; Watts, G.F.; Sahebkar, A. MicroRNAs: Novel Molecular Targets and Response Modulators of Statin Therapy. TIPS 2018, 39, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, M.W.; Zhao, Y.H.; Lu, Y.X.; Wang, Y.A.; Tong, C.W. Baicalin attenuates lipopolysaccharide-induced renal tubular epithelial cell injury by inhibiting the TXNIP/NLRP3 signalling pathway via increasing miR-223-3p expression. JBRHA 2020, 34, 69–82. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | SCC Value (Cells/mL) | DSCC Value (%) | Description |
|---|---|---|---|
| CTRL | <2 × 105 | <68.5 | Indicative values of absence of inflammation |
| SU | <2 × 105 | >68.5 | Indicative values of initial inflammation resulting in increased susceptibility to mastitis |
| AM | >2 × 105 | >68.5 | Indicative values of acute inflammatory state |
| CM | >2 × 105 | <68.5 | Indicative values of a chronic decease |
| Group | Parity | Days in Milk (d) | Milk Yield (kg/Sampling) |
|---|---|---|---|
| CTRL | 2.83 ± 1.34 | 184 ± 108 | 13.75 ± 4.81 |
| SU | 2.56 ± 1.34 | 205 ± 118 | 12.44 ± 3.79 |
| AM | 3.00 ± 2.14 | 181 ± 117 | 11.36 ± 4.38 |
| CM | 2.72 ± 1.93 | 217 ± 107 | 10.53 ± 3.84 |
| Group | Fat (%) | Protein (%) | Casein (%) | Lactose (%) | SCC (Cells/mL) | DSCC (%) |
|---|---|---|---|---|---|---|
| CTRL | 4.06 | 3.70 | 2.95 | 4.85 | 0.38 × 105 ± 0.01 | 52.52 |
| SU | 4.08 | 3.72 | 2.96 | 4.84 | 0.73 × 105 ± 0.02 | 76.73 |
| AM | 4.08 | 3.78 | 2.98 | 4.65 | 5.45 × 105 ± 0.30 | 79.37 |
| CM | 4.39 | 3.98 | 3.15 | 4.62 | 2.91 × 105 ± 0.62 | 60.80 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dall’Olio, E.; De Rensis, F.; Martignani, E.; Miretti, S.; Ala, U.; Cavalli, V.; Cipolat-Gotet, C.; Andrani, M.; Baratta, M.; Saleri, R. Differential Expression of miR-223-3p and miR-26-5p According to Different Stages of Mastitis in Dairy Cows. Biomolecules 2025, 15, 235. https://doi.org/10.3390/biom15020235
Dall’Olio E, De Rensis F, Martignani E, Miretti S, Ala U, Cavalli V, Cipolat-Gotet C, Andrani M, Baratta M, Saleri R. Differential Expression of miR-223-3p and miR-26-5p According to Different Stages of Mastitis in Dairy Cows. Biomolecules. 2025; 15(2):235. https://doi.org/10.3390/biom15020235
Chicago/Turabian StyleDall’Olio, Eleonora, Fabio De Rensis, Eugenio Martignani, Silvia Miretti, Ugo Ala, Valeria Cavalli, Claudio Cipolat-Gotet, Melania Andrani, Mario Baratta, and Roberta Saleri. 2025. "Differential Expression of miR-223-3p and miR-26-5p According to Different Stages of Mastitis in Dairy Cows" Biomolecules 15, no. 2: 235. https://doi.org/10.3390/biom15020235
APA StyleDall’Olio, E., De Rensis, F., Martignani, E., Miretti, S., Ala, U., Cavalli, V., Cipolat-Gotet, C., Andrani, M., Baratta, M., & Saleri, R. (2025). Differential Expression of miR-223-3p and miR-26-5p According to Different Stages of Mastitis in Dairy Cows. Biomolecules, 15(2), 235. https://doi.org/10.3390/biom15020235