
TrmL and TusA Are Necessary for rpoS and MiaA Is Required for hfq Expression in Escherichia coli

Abstract
:1. Introduction
2. Results
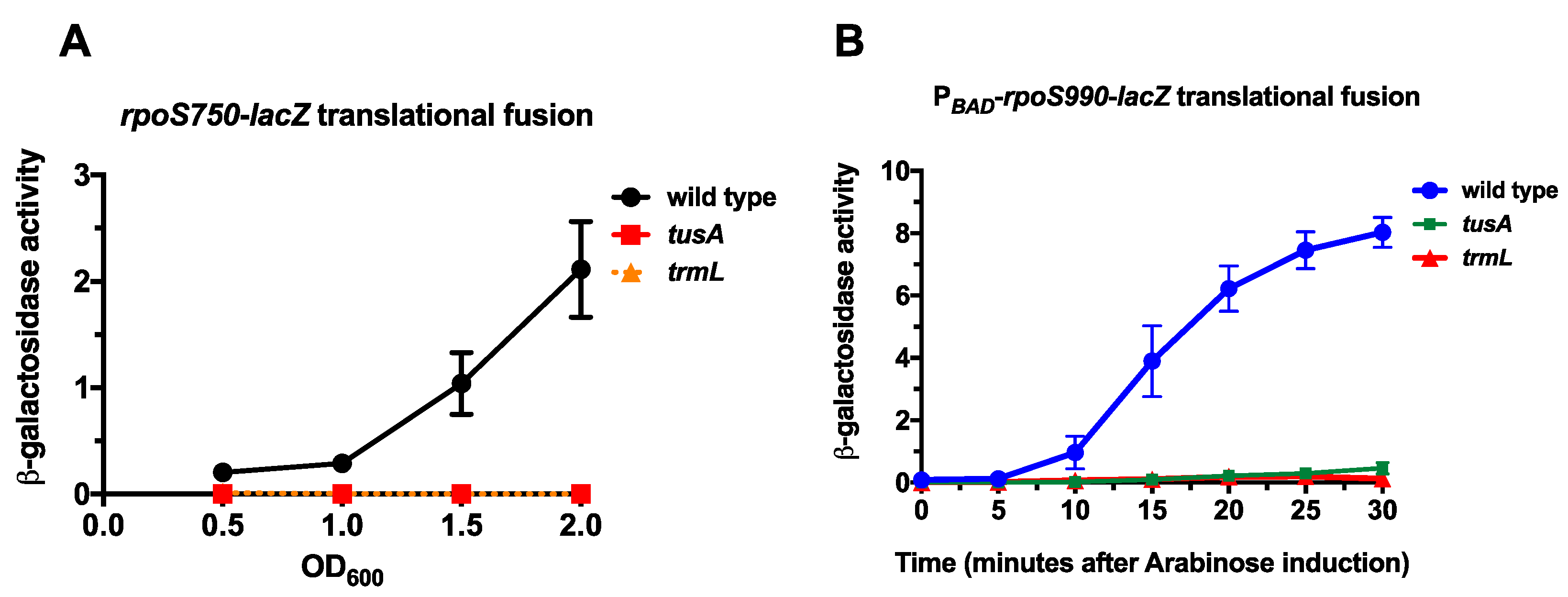
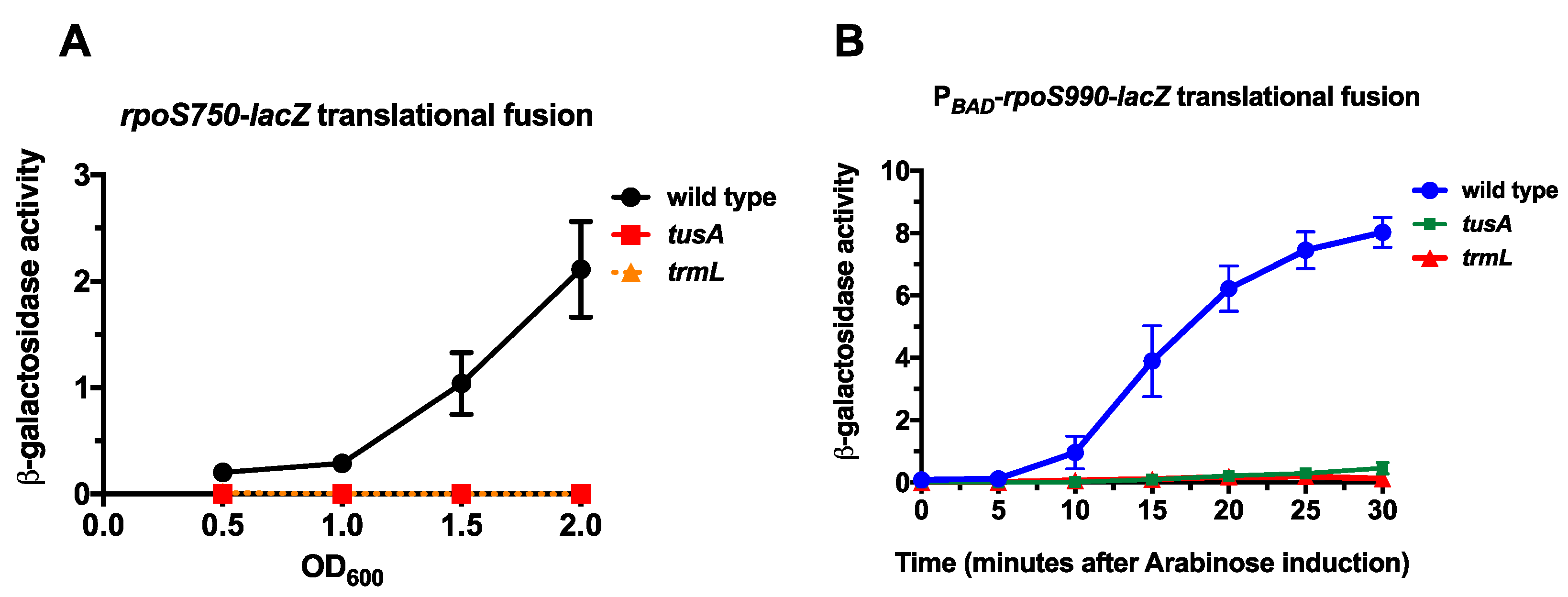
2.1. Wobble Base tRNA Modifications Enzymes TrmL and TusA Are Necessary for RpoS Expression
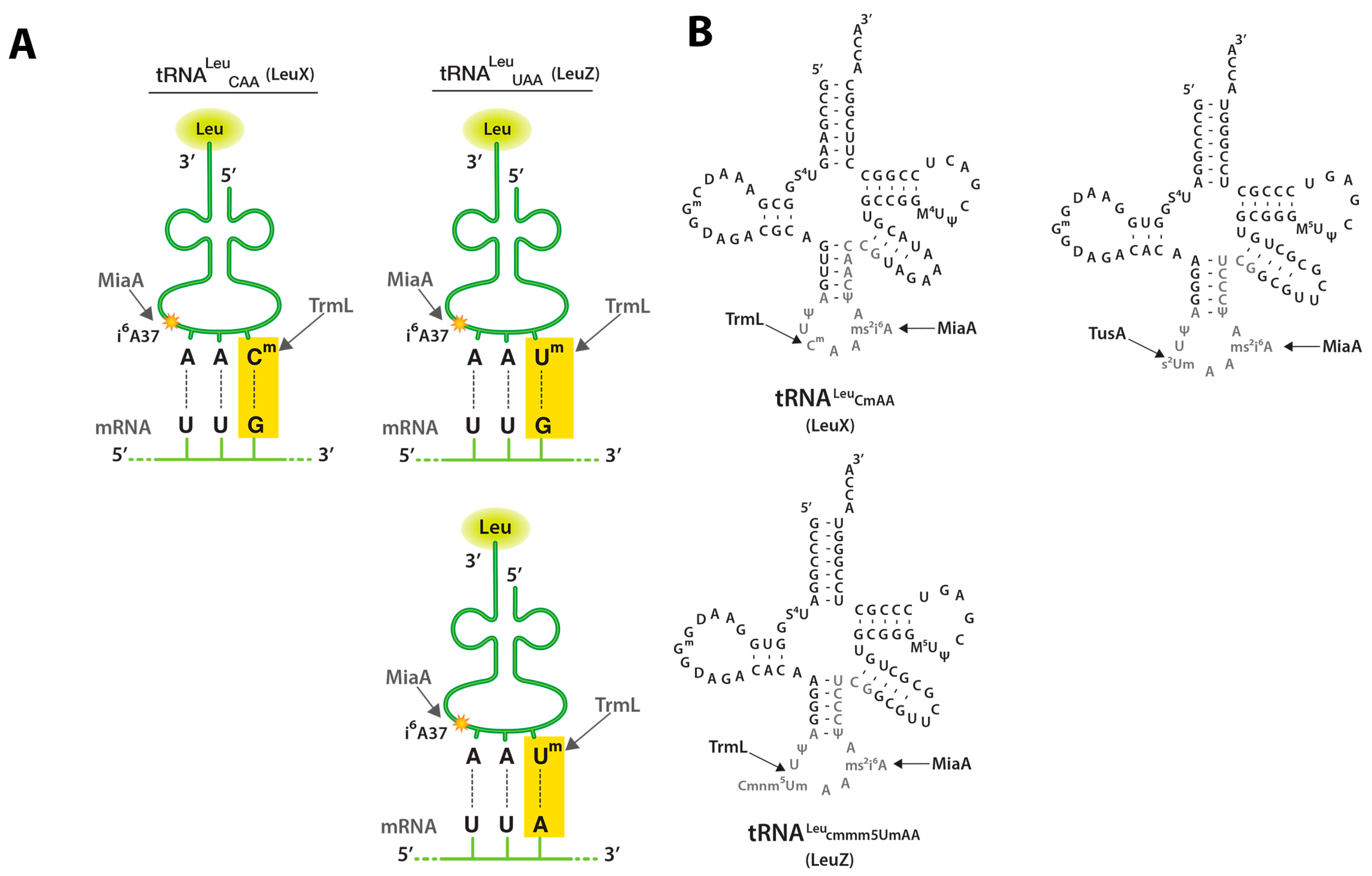
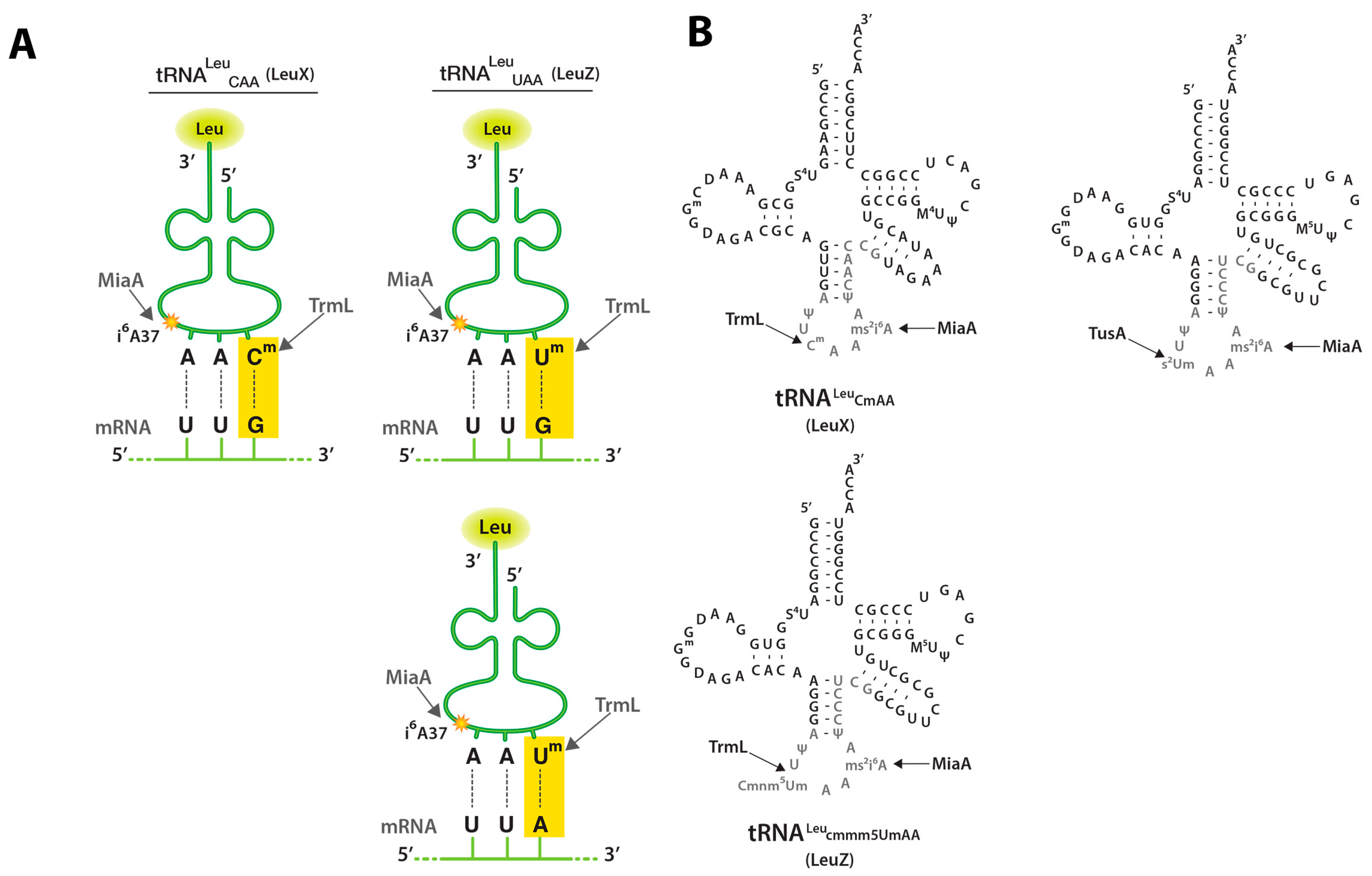
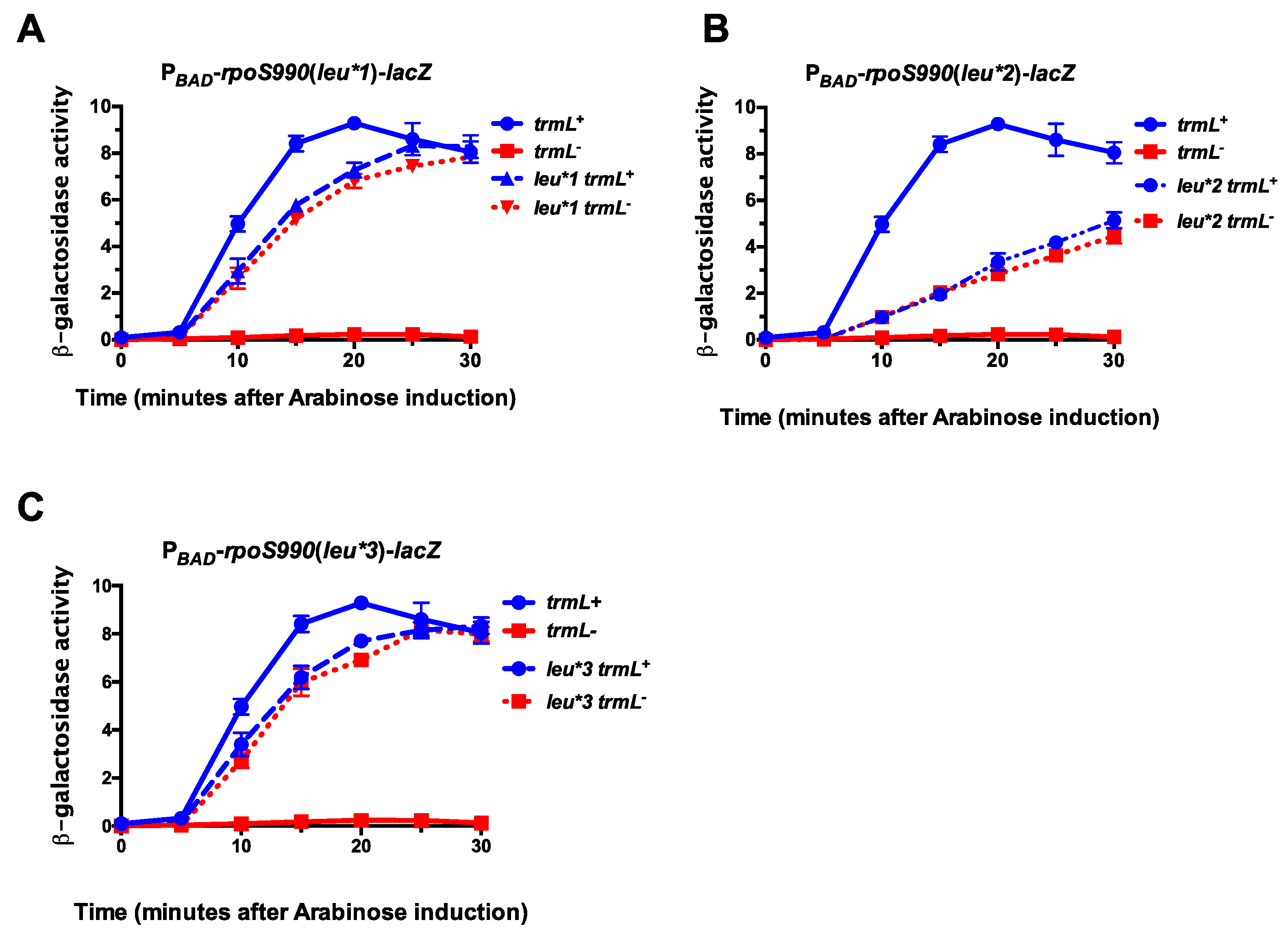
2.2. TrmL tRNA Modification is Necessary for Decoding of UUX-Leucine Decoding in RpoS
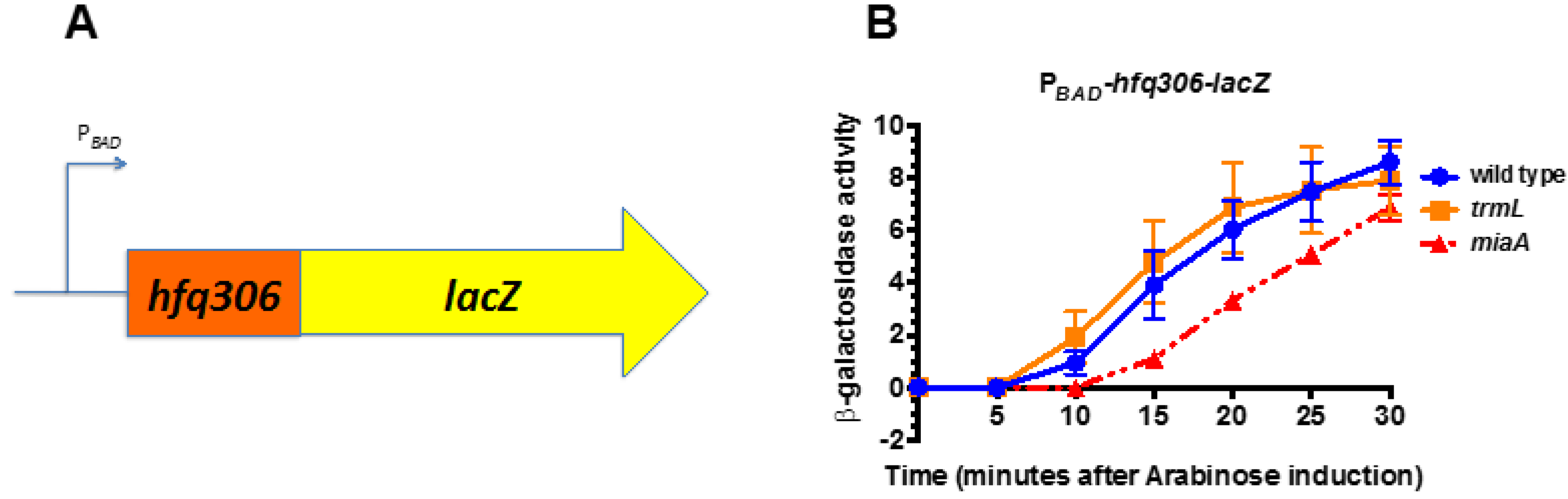
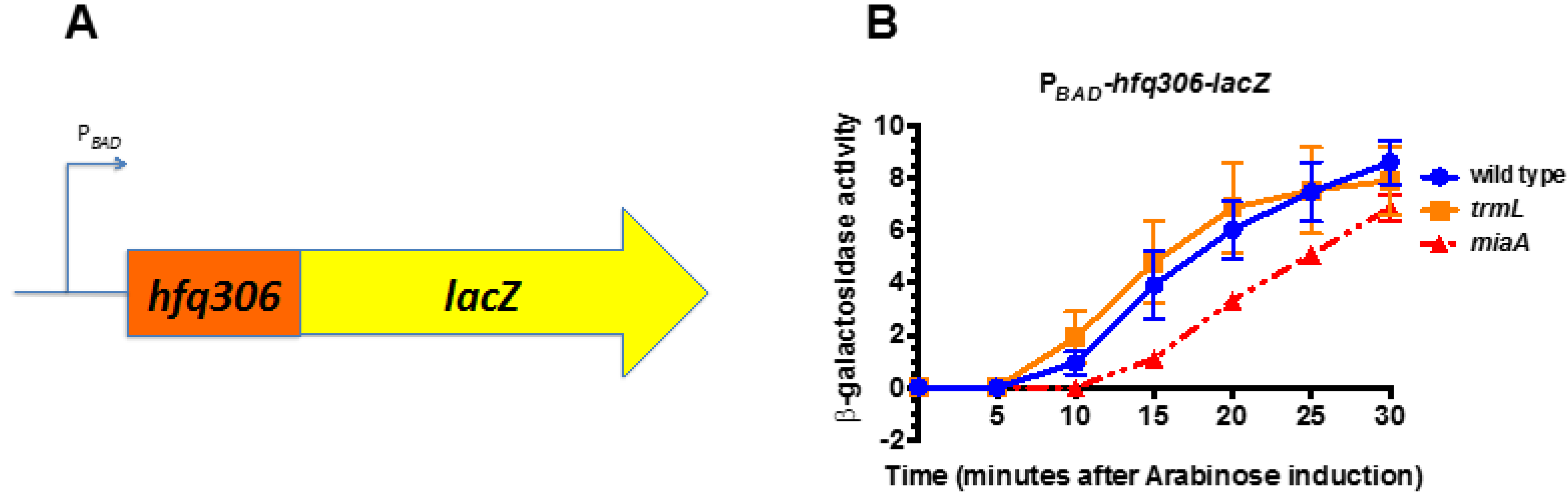
2.3. MiaA Is Necessary, While TrmL Is Dispensable, for Hfq Expression
3. Discussion
3.1. Expanded Network of tRNA Modifications Affecting rpoS Expression and Physiological Implications
3.2. TusA Catalyzed s2U34 and rpoS Translation
3.3. The i6A37 Requirement for Hfq and Implications for HULC Protein Predictive Model and Small RNA Biology
3.4. Implications for the Prokaryotic and Eukaryotic Organisms
4. Materials and Methods
4.1. Strains and Oligonucleotide Primers
4.2. Growth Conditions and Media
4.3. Genetic Constructions
4.4. β-Galactosidase Assays
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tanaka, K.; Takayanagi, Y.; Fujita, N.; Ishihama, A.; Takahashi, H. Heterogeneity of the principal σ factor in Escherichia coli: The rpoS gene product, σ38, is a second principal σ factor of RNA polymerase in stationary-phase Escherichia coli. Proc. Natl. Acad. Sci. USA 1993, 90, 8303. [Google Scholar] [CrossRef] [PubMed]
- Hengge-Aronis, R. Survival of hunger and stress: The role of rpoS in early stationary phase gene regulation in E. coli. Cell 1993, 72, 165–168. [Google Scholar] [CrossRef]
- Sledjeski, D.D.; Gupta, A.; Gottesman, S. The small RNA, DsrA, is essential for the low temperature expression of rpoS during exponential growth in Escherichia coli. EMBO J. 1996, 15, 3993–4000. [Google Scholar] [PubMed]
- Zhang, A.; Altuvia, S.; Tiwari, A.; Argaman, L.; Hengge-Aronis, R.; Storz, G. The OxyS regulatory RNA represses rpoS translation and binds the Hfq (HF-I) protein. EMBO J. 1998, 17, 6061–6068. [Google Scholar] [CrossRef] [PubMed]
- Majdalani, N.; Cunning, C.; Sledjeski, D.D.; Elliott, T.; Gottesman, S. Dsra RNA regulates translation of rpoS message by an anti-antisense mechanism, independent of its action as an antisilencer of transcription. Proc. Natl. Acad. Sci. USA 1998, 95, 12462–12467. [Google Scholar] [CrossRef] [PubMed]
- Majdalani, N.; Hernandez, D.; Gottesman, S. Regulation and mode of action of the second small RNA activator of rpoS translation, RprA. Mol. Microbiol. 2002, 46, 813–826. [Google Scholar] [CrossRef] [PubMed]
- Majdalani, N.; Chen, S.; Murrow, J.; St. John, K.; Gottesman, S. Regulation of rpoS by a novel small RNA: The characterization of RprA. Mol. Microbiol. 2001, 39, 1382–1394. [Google Scholar] [CrossRef] [PubMed]
- Mandin, P.; Gottesman, S. Integrating anaerobic/aerobic sensing and the general stress response through the ArcZ small RNA. EMBO J. 2010, 29, 3094–3107. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Gottesman, S. Regulation of proteolysis of the stationary-phase σ factor rpoS. J. Bacteriol. 1998, 180, 1154–1158. [Google Scholar] [PubMed]
- Bougdour, A.; Wickner, S.; Gottesman, S. Modulating RssB activity: IraP, a novel regulator of σs stability in Escherichia coli. Genes Dev. 2006, 20, 884–897. [Google Scholar] [CrossRef] [PubMed]
- Battesti, A.; Tsegaye, Y.M.; Packer, D.G.; Majdalani, N.; Gottesman, S. H-NS regulation of IraD and IraM antiadaptors for control of rpos degradation. J. Bacteriol. 2012, 194, 2470–2478. [Google Scholar] [CrossRef] [PubMed]
- Bougdour, A.; Cunning, C.; Baptiste, P.J.; Elliott, T.; Gottesman, S. Multiple pathways for regulation of sigmas (rpoS) stability in Escherichia coli via the action of multiple anti-adaptors. Mol. Microbiol. 2008, 68, 298–313. [Google Scholar] [CrossRef] [PubMed]
- Battesti, A.; Majdalani, N.; Gottesman, S. Stress σ factor rposS degradation and translation are sensitive to the state of central metabolism. Proc. Natl. Acad. Sci. USA 2015, 112, 5159–5164. [Google Scholar] [CrossRef] [PubMed]
- Gottesman, S.; Roche, E.; Zhou, Y.; Sauer, R.T. The ClpXP and ClpaAP proteases degrade proteins with carboxy-terminal peptide tails added by the SsrA-tagging system. Genes Dev. 1998, 12, 1338–1347. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.M.; Gottesman, S. The MiaA tRNA modification enzyme is necessary for robust rpoS expression in Escherichia coli. J. Bacteriol. 2014, 196, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Caillet, J.; Droogmans, L. Molecular cloning of the Escherichia coli miaA gene involved in the formation of delta 2-isopentyl adenosine in tRNA. J. Bacteriol. 1988, 170, 4147–4152. [Google Scholar] [CrossRef] [PubMed]
- Connolly, D.M.; Winkler, M.E. Structure of Escherichia coli K-12 miaa and characterization of the mutator phenotype caused by miaa insertion mutations. J. Bacteriol. 1991, 173, 1711–1721. [Google Scholar] [CrossRef] [PubMed]
- Connolly, D.M.; Winkler, M.E. Genetic and physiological relationships among the miaA gene, 2-methylthio-N6-(delta 2-isopentenyl)-adenosine tRNA modification, and spontaneous mutagenesis in Escherichia coli K-12. J. Bacteriol. 1989, 171, 3233–3246. [Google Scholar] [CrossRef] [PubMed]
- Aubee, J.I.; Olu, M.; Thompson, K.M. The i6a37 tRNA modification is essential for proper decoding of UUX-Leucine codons during rpoS and iraP translation. RNA 2016, 22, 729–742. [Google Scholar] [CrossRef] [PubMed]
- Wobble. Available online: http://www.mun.ca/biology/scarr/iGen3_06-08.html (accessed on 24 April 2017).
- Benitez-Paez, A.; Villarroya, M.; Douthwaite, S.; Gabaldon, T.; Armengod, M.E. Yibk is the 2-o-methyltransferase trml that modifies the wobble nucleotide in Escherichia coli tRNA(leu) isoacceptors. RNA 2010, 16, 2131–2143. [Google Scholar] [CrossRef] [PubMed]
- Klassen, R.; Bruch, A.; Schaffrath, R. Independent suppression of ribosomal +1 frameshifts by different tRNA anticodon loop modifications. RNA Biol. 2016, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tukenmez, H.; Xu, H.; Esberg, A.; Bystrom, A.S. The role of wobble uridine modifications in +1 translational frameshifting in eukaryotes. Nucleic Acids Res. 2015, 43, 9489–9499. [Google Scholar] [CrossRef] [PubMed]
- Urbonavicius, J.; Qian, Q.; Durand, J.M.; Hagervall, T.G.; Bjork, G.R. Improvement of reading frame maintenance is a common function for several tRNA modifications. EMBO J. 2001, 20, 4863–4873. [Google Scholar] [CrossRef] [PubMed]
- Patil, A.; Dyavaiah, M.; Joseph, F.; Rooney, J.P.; Chan, C.T.; Dedon, P.C.; Begley, T.J. Increased tRNA modification and gene-specific codon usage regulate cell cycle progression during the DNA damage response. Cell Cycle 2012, 1, 3656–3665. [Google Scholar] [CrossRef] [PubMed]
- Endres, L.; Dedon, P.C.; Begley, T.J. Codon-biased translation can be regulated by wobble-base tRNA modification systems during cellular stress responses. RNA Biol. 2015, 12, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.J.; Zhou, M.; Fang, Z.P.; Wang, M.; Zhou, X.L.; Wang, E.D. The tRNA recognition mechanism of the minimalist SPOUT methyltransferase, TrmL. Nucleic Acids Res. 2013, 41, 7828–7842. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Long, T.; Fang, Z.P.; Zhou, X.L.; Liu, R.J.; Wang, E.D. Identification of determinants for tRNA substrate recognition by Escherichia coli C/U34 2′-O-methyltransferase. RNA Biol. 2015, 12, 900–911. [Google Scholar] [CrossRef] [PubMed]
- Rogers, K.C.; Crescenzo, A.T.; Soll, D. Aminoacylation of transfer RNAs with 2-thiouridine derivatives in the wobble position of the anticodon. Biochimie 1995, 77, 66–74. [Google Scholar] [CrossRef]
- Rodriguez-Hernandez, A.; Spears, J.L.; Gaston, K.W.; Limbach, P.A.; Gamper, H.; Hou, Y.; Kaiser, R.; Agris, P.F.; Perona, J.J. Structural and mechanistic basis for enhanced translational efficiency by 2-thiouridine at the tRNA anticodon wobble position. J. Mol. Biol. 2013, 425, 3888–3906. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, Y.; Shigi, N.; Kato, J.; Nishimura, A.; Suzuki, T. Mechanistic insights into sulfur relay by multiple sulfur mediators involved in thiouridine biosynthesis at tRNA wobble positions. Mol. Cell 2006, 21, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Yamashino, T.; Isomura, M.; Ueguchi, C.; Mizuno, T. The yhhp gene encoding a small ubiquitous protein is fundamental for normal cell growth of Escherichia coli. J. Bacteriol. 1998, 180, 2257–2261. [Google Scholar] [PubMed]
- Tsui, H.C.; Leung, H.C.; Winkler, M.E. Characterization of broadly pleiotropic phenotypes caused by an hfq insertion mutation in Escherichia coli K-12. Mol. Microbiol. 1994, 13, 35–49. [Google Scholar] [CrossRef] [PubMed]
- De Lay, N.; Schu, D.J.; Gottesman, S. Bacterial small RNA-based negative regulation: Hfq and its accomplices. J. Biol. Chem. 2013, 288, 7996–8003. [Google Scholar] [CrossRef] [PubMed]
- Kajitani, M.; Ishihama, A. Identification and sequence determination of the host factor gene for bacteriophage Q beta. Nucleic Acids Res. 1991, 19, 1063–1066. [Google Scholar] [CrossRef] [PubMed]
- Tsui, H.C.; Feng, G.; Winkler, M.E. Transcription of the mutL repair, miaA tRNA modification, hfq pleiotropic regulator, and hflA region protease genes of Escherichia coli K-12 from clustered Eσ32-specific promoters during heat shock. J. Bacteriol. 1996, 178, 5719–5731. [Google Scholar] [CrossRef] [PubMed]
- Armengod, M.E.; Meseguer, S.; Villarroya, M.; Prado, S.; Moukadiri, I.; Ruiz-Partida, R.; Garzon, M.J.; Navarro-Gonzalez, C.; Martinez-Zamora, A. Modification of the wobble uridine in bacterial and mitochondrial tRNAs reading NNA/NNG triplets of 2-codon boxes. RNA Biol. 2014, 11, 1495–1507. [Google Scholar] [CrossRef] [PubMed]
- Armengod, M.E.; Moukadiri, I.; Prado, S.; Ruiz-Partida, R.; Benitez-Paez, A.; Villarroya, M.; Lomas, R.; Garzon, M.J.; Martinez-Zamora, A.; Meseguer, S.; et al. Enzymology of tRNA modification in the bacterial mnmeg pathway. Biochimie 2012, 94, 1510–1520. [Google Scholar] [CrossRef] [PubMed]
- Blum, P.H. Reduced leu operon expression in a miaA mutant of Salmonella typhimurium. J. Bacteriol. 1988, 170, 5125–5133. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Vogel, J. Multiple target regulation by small noncoding RNAs rewires gene expression at the post-transcriptional level. Res Microbiol. 2009, 160, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Tsui, H.C.; Winkler, M.E. Transcriptional patterns of the mutl-miaA superoperon of Escherichia coli K-12 suggest a model for posttranscriptional regulation. Biochimie 1994, 76, 1168–1177. [Google Scholar] [CrossRef]
- Moon, K.; Gottesman, S. Competition among hfq-binding small RNAs in Escherichia coli. Mol. Microbiol. 2011, 82, 1545–1562. [Google Scholar] [CrossRef] [PubMed]
- Patil, A.; Chan, C.T.; Dyavaiah, M.; Rooney, J.P.; Dedon, P.C.; Begley, T.J. Translational infidelity-induced protein stress results from a deficiency in Trm9-catalyzed tRNA modifications. RNA Biol. 2012, 9, 990–1001. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.T.; Pang, Y.L.; Deng, W.; Babu, I.R.; Dyavaiah, M.; Begley, T.J.; Dedon, P.C. Reprogramming of tRNA modifications controls the oxidative stress response by codon-biased translation of proteins. Nat. Commun. 2012, 3, 937. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.T.; Dyavaiah, M.; DeMott, M.S.; Taghizadeh, K.; Dedon, P.C.; Begley, T.J. A quantitative system approach reveals dynamic control of tRNA modifications during cellular stress. PLoS Genet. 2010, 6, e1001247. [Google Scholar] [CrossRef] [PubMed]
- Begley, T.J.; Dyavaiah, M.; Patil, A.; Rooney, J.P.; Direnzo, D.; Young, C.M.; Conklin, D.S.; Zitomer, R.S.; Begley, T.J. Trm-9 catalized tRNA modifications link translation to the DNA damage response. Mol. Cell 2007, 28, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, T.N.; Blewett, N.H.; Crawford, A.K.; Cherkasova, V.A.; Iben, J.R.; Begley, T.J.; Farabaugh, P.J.; Maraia, R.J. Lack of tRNA modification isopentenyl-a37 alters mRNA decoding and causes metabolic deficiences in fission yeast. Mol. Cell. Biol. 2013, 33, 2918–2929. [Google Scholar] [CrossRef] [PubMed]
- Chionh, Y.H.; McBee, M.; Babu, I.R.; Hia, F.; Lin, W.; Zhao, W.; Cao, J.; Dziergowska, A.; Malkiewicz, A.; Begley, T.J.; et al. tRNA-mediated codon-biased translation in mycobacterial hypoxic persistence. Nat. Commun. 2016, 11, 13302. [Google Scholar] [CrossRef] [PubMed]
- Meseguer, S.; Martinez-Zamora, A.; Garcia-Arumi, E.; Andreu, A.L.; Armengod, M.E. The ROS-sensitive microRNA-9/9* controls the expression of mitochondrial tRNA modifyng enzymes and is involved in the molecular mechanism of melas syndrome. Hum. Mol. Genet. 2015, 24, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Court, D.L.; Swaminathan, S.; Yu, D.; Wilson, H.; Baker, T.; Bubunenko, M.; Sawitzke, J.; Sharan, S.K. Mini-λ: A tractable system for chromosome and bac engineering. Gene 2003, 315, 63–69. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (Min after Ara Induction) | Strain (Mean β-Gal Activity) | ||
|---|---|---|---|
| hfq+ | hfq- | hfq+/hfq- Fold Change | |
| 10 | 0.94 | 0.00 | ∞ |
| 15 | 3.91 | 1.11 | 3.52 |
| 20 | 6.04 | 3.35 | 1.80 |
| 25 | 7.50 | 5.10 | 1.47 |
| 30 | 8.62 | 6.89 | 1.25 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aubee, J.I.; Olu, M.; Thompson, K.M. TrmL and TusA Are Necessary for rpoS and MiaA Is Required for hfq Expression in Escherichia coli. Biomolecules 2017, 7, 39. https://doi.org/10.3390/biom7020039
Aubee JI, Olu M, Thompson KM. TrmL and TusA Are Necessary for rpoS and MiaA Is Required for hfq Expression in Escherichia coli. Biomolecules. 2017; 7(2):39. https://doi.org/10.3390/biom7020039
Chicago/Turabian StyleAubee, Joseph I., Morenike Olu, and Karl M. Thompson. 2017. "TrmL and TusA Are Necessary for rpoS and MiaA Is Required for hfq Expression in Escherichia coli" Biomolecules 7, no. 2: 39. https://doi.org/10.3390/biom7020039
APA StyleAubee, J. I., Olu, M., & Thompson, K. M. (2017). TrmL and TusA Are Necessary for rpoS and MiaA Is Required for hfq Expression in Escherichia coli. Biomolecules, 7(2), 39. https://doi.org/10.3390/biom7020039