Poly-β-Hydroxybutyrate Production by the Cyanobacterium Scytonema geitleri Bharadwaja under Varying Environmental Conditions

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Organism and Culture Conditions
2.2. Growth Measurement
2.3. Extraction of Polyhydroxybutyrate
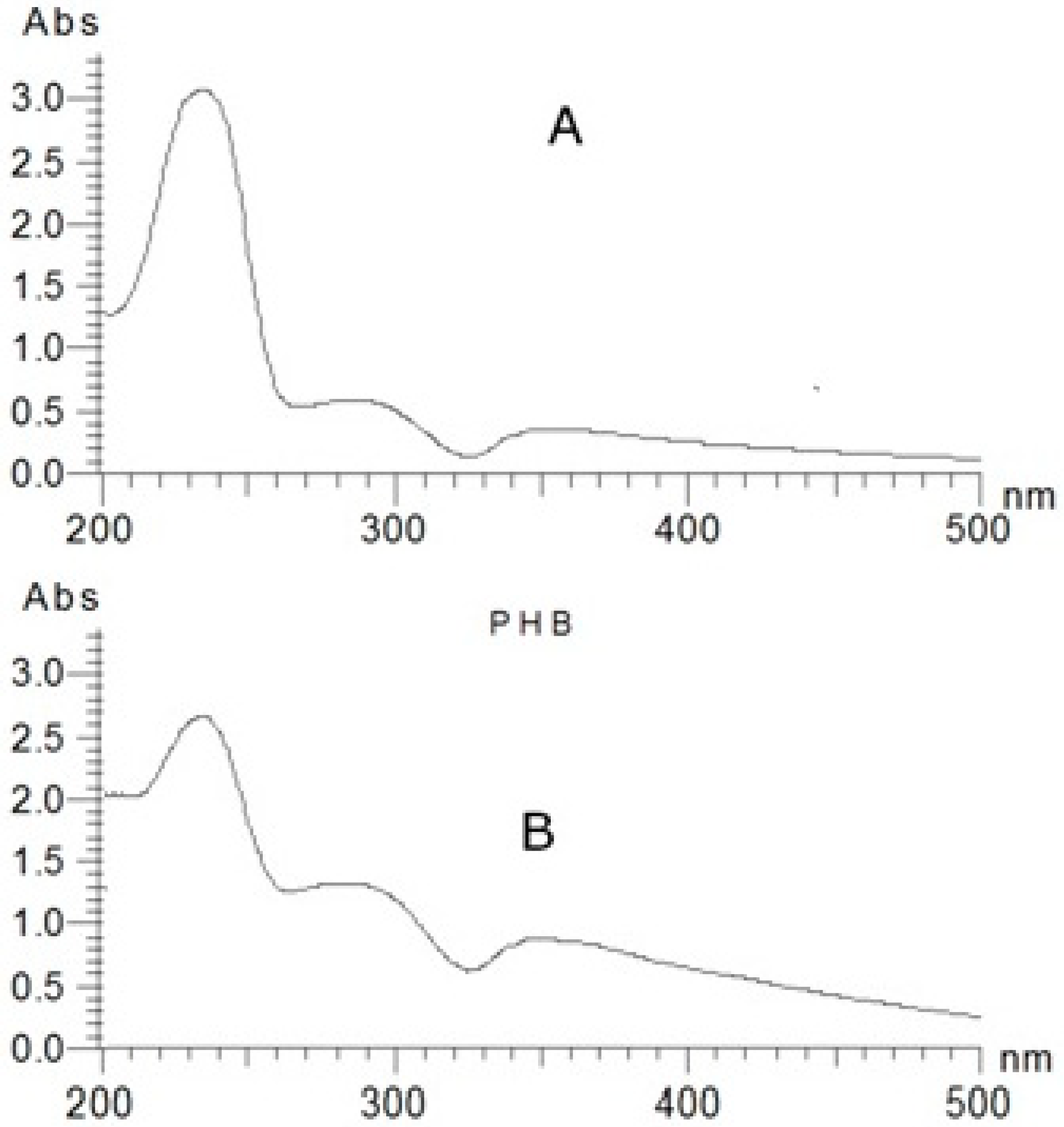
2.4. Assay of Polyhydroxybutyrate
2.5. Quantification of Polyhydroxybutyrate
3. Results and Discussion
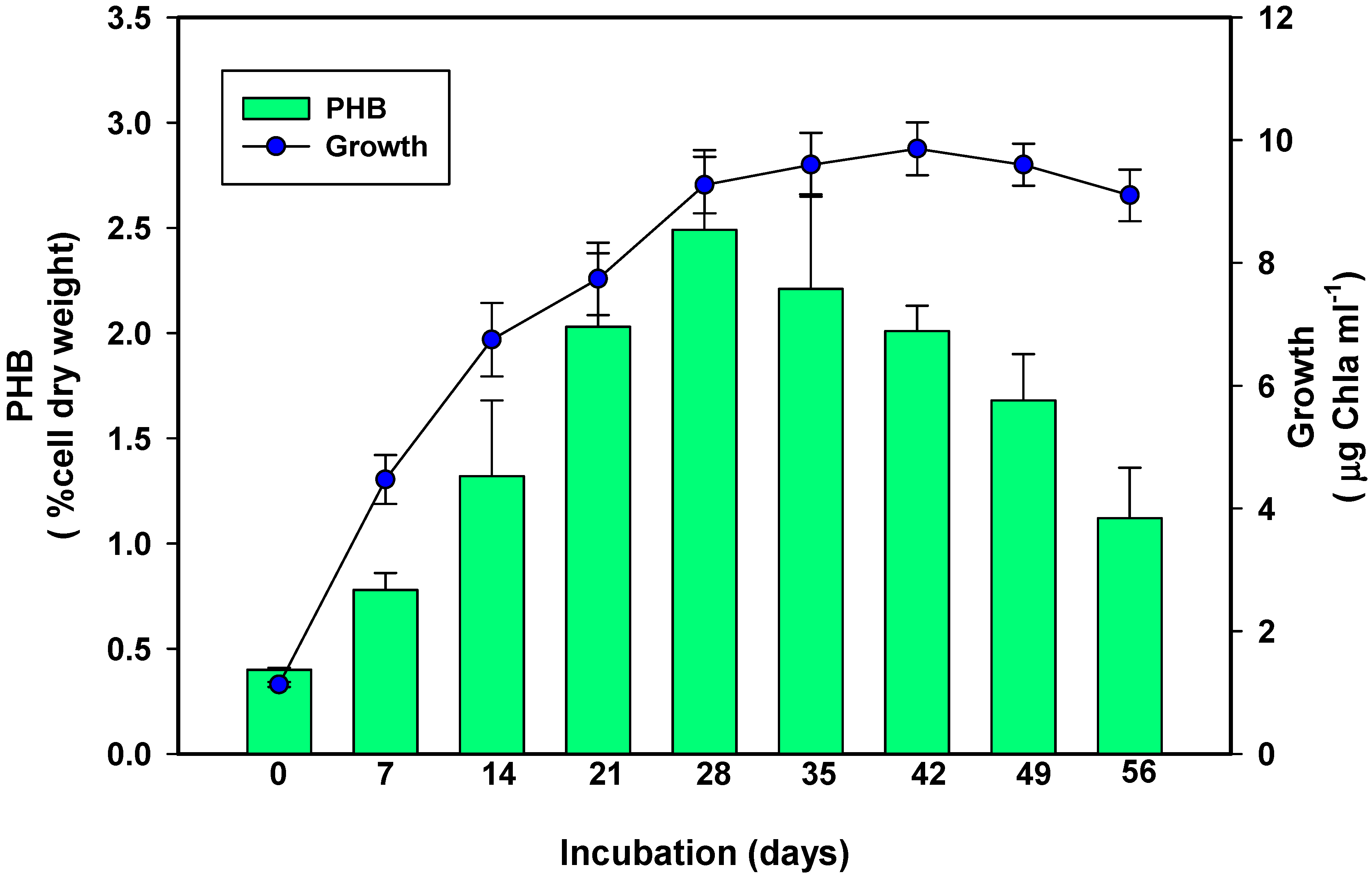
3.1. Relationship between Growth and Polyhydroxybutyrate Production
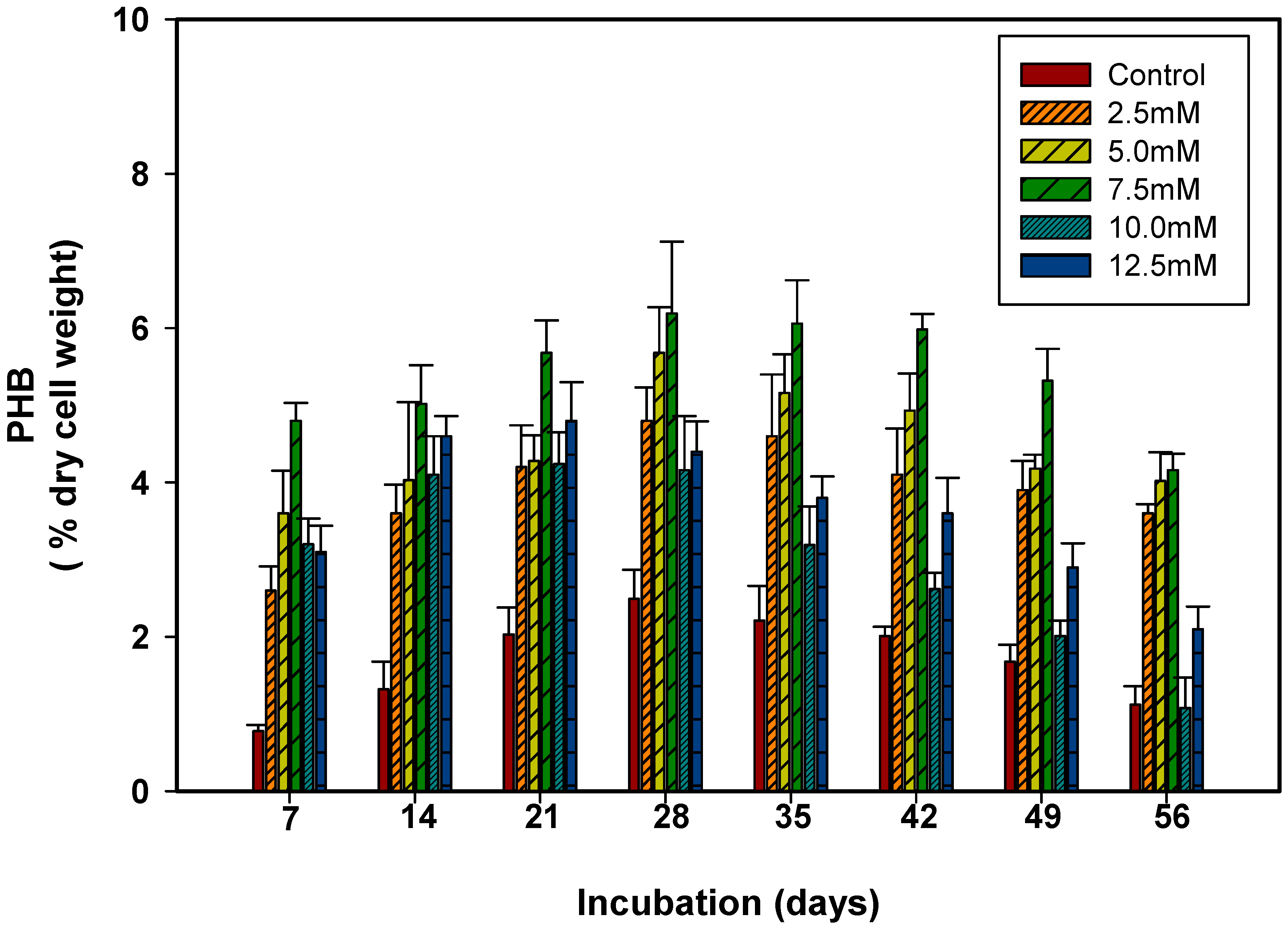
3.2. Relative Contribution of Carbon Sources on Polyhydroxybutyrate Production
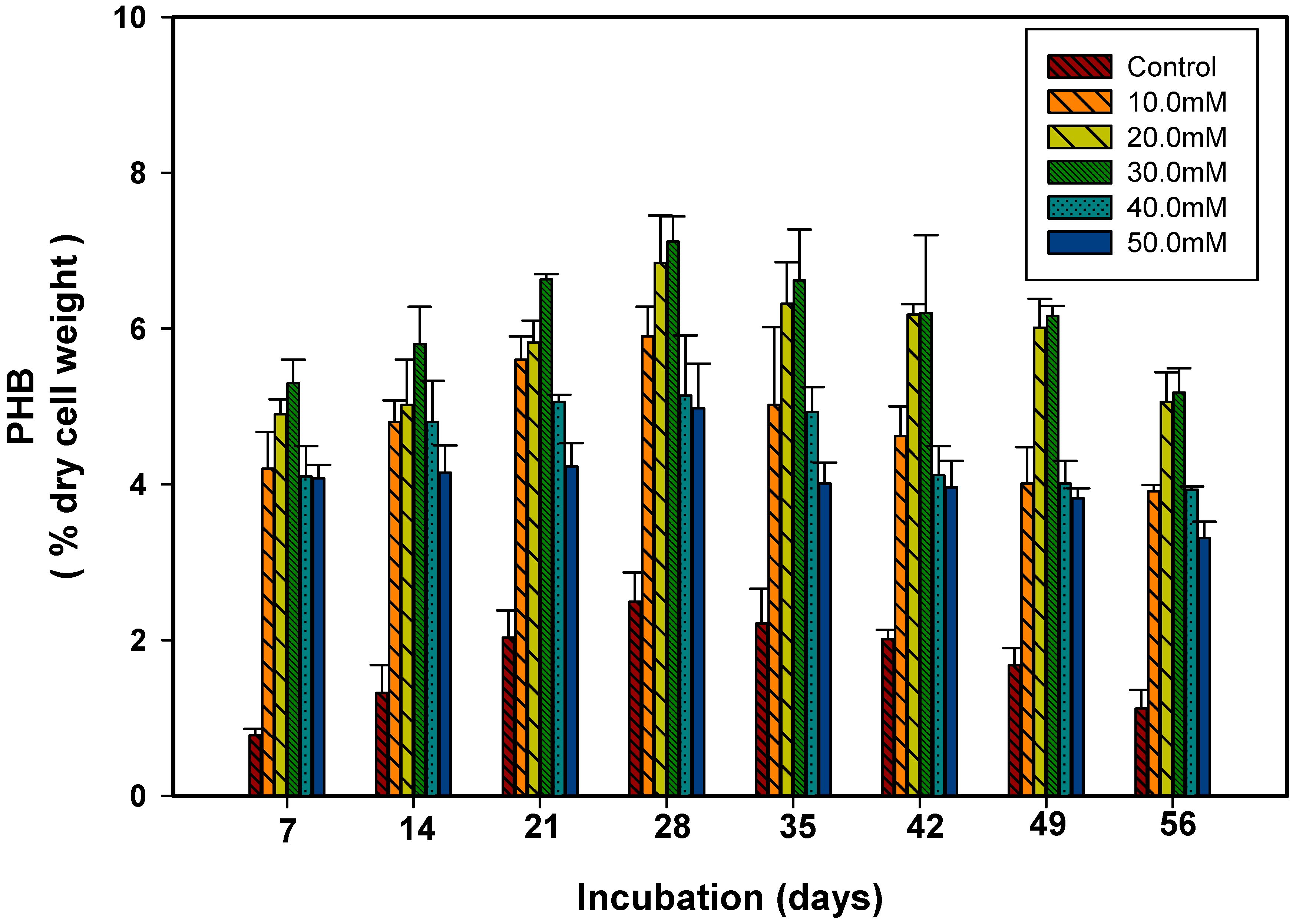
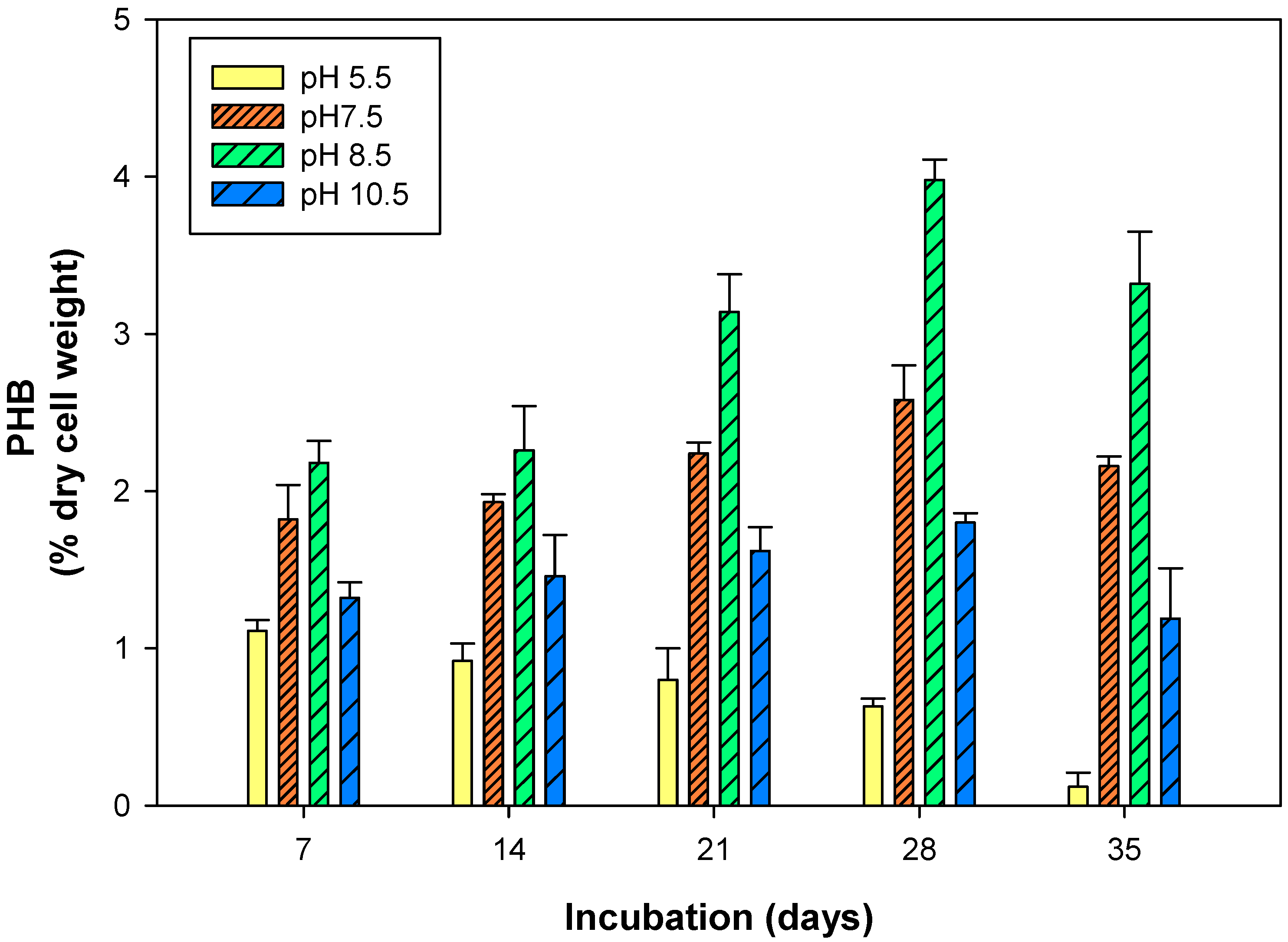
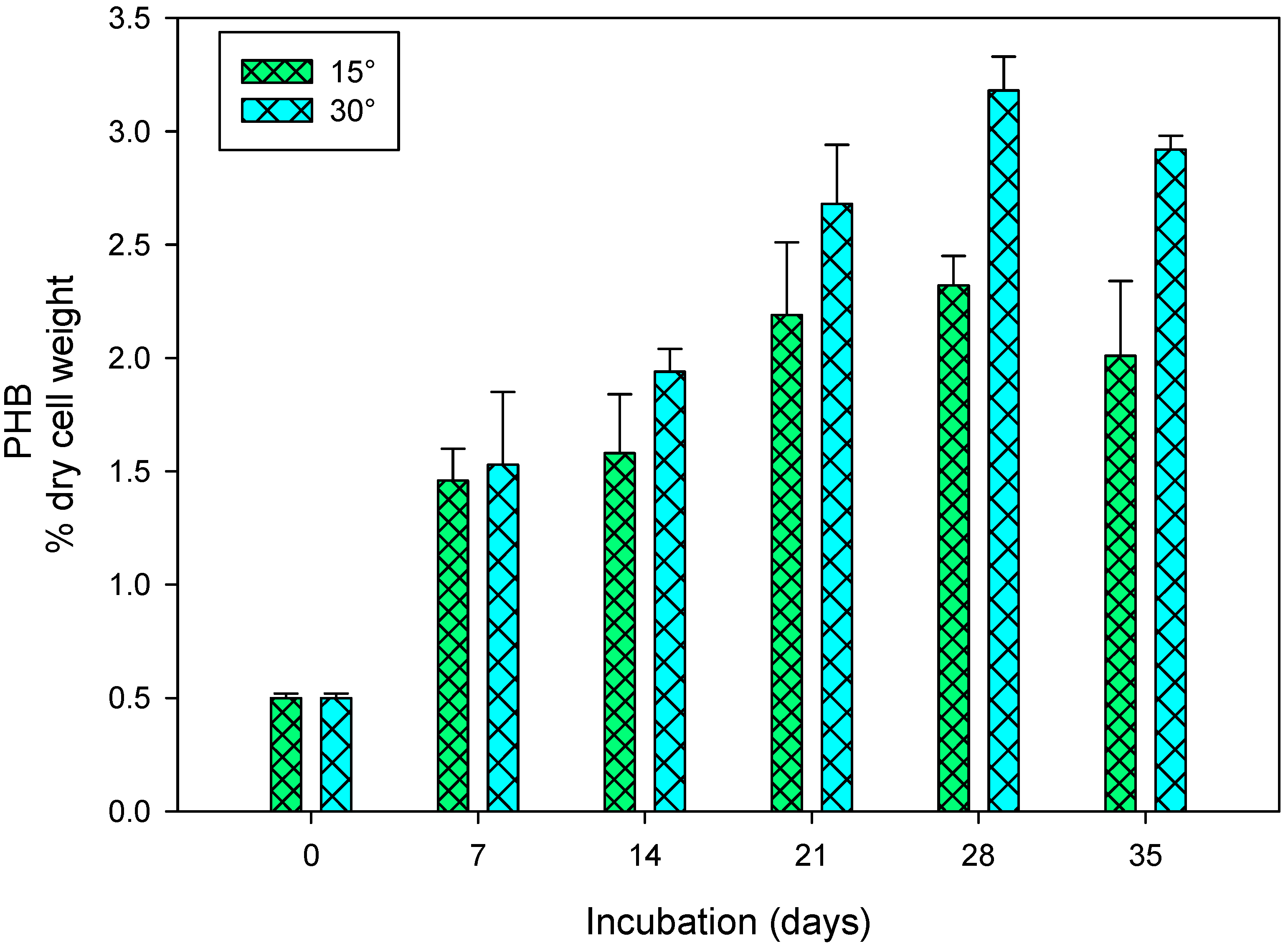
3.3. Polyhydroxybutyrate Production in Response to pH and Temperature
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Singh, M.; Patel, S.K.; Kalia, V.C. Bacillus subtilis as potential producer for polyhydroxyalkanoates. Microb. Cell Factories 2009, 8, 38. [Google Scholar] [CrossRef] [PubMed]
- Liebergesell, M.; Sonomoto, K.; Madkour, M.; Mayer, F.; Steinbüchel, A. Purification and Characterization of the Poly(Hydroxyalkanoic Acid) Synthase from Chromatium vinosum and Localization of the Enzyme at the Surface of Poly(Hydroxyalkanoic Acid) Granules. JBIC J. Boil. Inorg. Chem. 1994, 226, 71–80. [Google Scholar]
- Stubbe, J.; Tian, J.; He, A.; Sinskey, A.J.; Lawrence, A.G.; Liu, P. Nontemplate-Dependent Polymerization Processes: Polyhydroxyalkanoate Synthases as a Paradigm. Annu. Biochem. 2005, 74, 433–480. [Google Scholar] [CrossRef]
- Tian, J.; Stubbe, J. Polyhydroxyalkanoate (PHA) homeostasis: The role of the PHA synthase. Nat. Prod. Rep. 2003, 20, 445. [Google Scholar]
- Fukada, E.; Ando, Y. Piezoelectric properties of poly-β-hydroxybytyrate and copolymers of β-hydroxybutyrate and β-hydroxyvalerate. Int. J. Biol. Macromol. 1986, 8, 361–366. [Google Scholar] [CrossRef]
- Duvernoy, O.; Malm, T.; Ramström, J.; Bowald, S. A Biodegradable Patch used as a Pericardial Substitute after Cardiac Surgery: 6- and 24-Month Evaluation with CT. Thorac. Cardiovasc. 1995, 43, 271–274. [Google Scholar] [CrossRef]
- Samantaray, S.; Mallick, N. Production and characterization of poly-β-hydroxybutyrate (PHB) polymer from Aulosira fertilissima. J. Appl. Phycol. 2012, 24, 803–814. [Google Scholar] [CrossRef]
- Carr, N. The occurrence of poly-β-hydroxybutyrate in the blue-green alga, Chlorogloea fritschii. Biochim. Biophys. Acta 1966, 120, 308–310. [Google Scholar] [CrossRef]
- Campbell, J.; Stevens, S.E.; Bankwill, D.L. Accumulation of poly-β-hydroxybutyrate in Spirulina platensis. J. Bacteriol. 1982, 149, 361–366. [Google Scholar] [PubMed]
- Stal, L.J.; Heyer, H.; Jacobs, G. Occurrence and Role of Poly-Hydroxy-Alkanoate in the Cyanobacterium Oscillatoria Limosa. In Novel Biodegradable Microbial Polymers; Dawes, E.A., Ed.; NATO ASI Series (Series E: Applied Sciences); Springer: Dordrecht, The Netherlands, 1990; Volume 186, pp. 435–438. [Google Scholar]
- Vincenzini, M.; Sili, C.; De Philippis, R.; Ena, A.; Materassi, R. Occurrence of poly-β-hydroxybutyrate in Spirulina species. J. Bacteriol. 1990, 172, 2791–2792. [Google Scholar] [CrossRef]
- Shrivastav, A.; Mishra, S.K.; Mishra, S. Polyhydroxyalkanoates synthesis by Spirulina subsalsa from Gujrat coast of India. Int. J. Biol. Macromol. 2010, 46, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Wu, Q.; Shen, Z. Accumulation of poly-β-hydroxybutyrate in cyanobacterium Synechocystis sp. PCC6803. Bioresour. Technol. 2001, 76, 85–90. [Google Scholar] [CrossRef]
- Panda, B.; Mallick, N. Enhanced poly-β-hydroxybutyrate accumulation in a unicellular cyanobacterium, Synechocystis sp. PCC 6803. Lett. Appl. Microbiol. 2007, 44, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Mallick, N. Accumulation of poly-β-hydroxybutyrate in Nostoc muscorum: Regulation by pH, light-dark cycles, N and P status and carbon sources. Bioresour. Technol. 2005, 96, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Mallick, N. Enhancement of poly-β-hydroyxybutyrate accumulation in Nostoc muscorum under mixotrophy, chemoheterotrophy and limitation of gas exchange. Biotechnol. Lett. 2005, 27, 59–62. [Google Scholar] [CrossRef]
- Kamravamanesh, D.; Kovacs, T.; Pflügl, S.; Druzhinina, I.; Kroll, P.; Lackner, M.; Herwig, C. Increased poly-β-hydroxybutyrate production from carbon dioxide in randomly mutated cells of cyanobacterial strain Synechocystis sp. PCC 6714: Mutant generation and characterization. Bioresour. Technol. 2018, 266, 34–44. [Google Scholar] [CrossRef]
- Troschl, C.; Meixner, K.; Fritz, I.; Leitner, K.; Romero, A.P.; Kovalcik, A.; Sedlacek, P.; Drosg, B. Pilot-scale production of poly-β-hydroxybutyrate with the cyanobacterium Synechocytis sp. CCALA192 in a non-sterile tubular photobioreactor. Algal. Res. 2018, 34, 116–125. [Google Scholar] [CrossRef]
- Hassan, M.A.; Bakhiet, E.K.; Ali, S.G.; Hussien, H.R. Production and characterization of polyhydroxybutyrate (PHB) produced by Bacillus sp. isolated from Egypt. J. Appl. Pharm. Sci. 2016, 6, 046–051. [Google Scholar] [CrossRef]
- Desikachary, T.V. Cyanophyta; Indian Council of Agricultural Research: New Delhi, India, 1959. [Google Scholar]
- Chu, S.P. The influence of the mineral composition of the medium on the growth of planktonic algae. I. Method and culture media. J. Ecol. 1942, 30, 284–325. [Google Scholar] [CrossRef]
- Mackinney, G. Absorption of light by chlorophyll solution. J. Biol. Chem. 1941, 140, 315–322. [Google Scholar]
- Williamson, D.H.; Wilkinson, J.F. The isolation and estimation of the poly-β-hydroxy- butyrate inclusions of Bacillus species. J. Gen. Microbiol. 1958, 19, 198–209. [Google Scholar] [CrossRef]
- Huang, R.; Reusch, R.N. Poly (β-hydroxybutyrate) is associated with specific protein in the cytoplasm and membrane of Escherichia coli. J. Biol. Chem. 1996, 271, 22196–22202. [Google Scholar] [CrossRef]
- Law, J.H.; Slepecky, R.A. Assay of Poly-Β-Hydroxybutyric Acid. J. Bacteriol. 1961, 82, 33–36. [Google Scholar]
- Brandl, H.; Gross, R.A.; Lenz, R.W.; Fuller, R.C. Pseudomonas oleovorans as a Source of Poly(β-Hydroxyalkanoates) for Potential Applications as Biodegradable Polyesters. Appl. Environ. Microbiol. 1988, 54, 1977–1982. [Google Scholar]
- Braunegg, G.; Sonnleimer, B.; Lafferty, R.M. A rapid gas chromatographic method for the determination of Poly-β-hydroxybutyric acid in microbial biomass. Eur. J. Appl. Microbiol. Biotechnol. 1978, 6, 29–37. [Google Scholar] [CrossRef]
- Cui, Y.; Barford, J.P.; Renneberg, R. Gas Chromatographic Determination of Poly(3-hydroxybutyrate) with Alkaline Hydrolysis and Acid Esterification. Anal. Lett. 2007, 40, 2915–2924. [Google Scholar] [CrossRef]
- Ansari, S.; Fatma, T. Cyanobacterial Polyhydroxybutyrate (PHB): Screening, Optimization and Characterization. PLoS ONE 2016, 11, 0158168. [Google Scholar] [CrossRef] [PubMed]
- Stal, L.J. Poly(hydroxyalkanoates) in cyanobacteria: A review. FEMS Microbiol. Rev. 1992, 103, 169–180. [Google Scholar] [CrossRef]
- Mallick, N. Polyhydroxyalkanoates in cyanobacteria: Progress and prospects. In New Frontiers of Environmental Biotechnological Applications; Santra, S.C., Mallik, A., Eds.; ENVIS Centre on Environmental Biotechnology: Kolkata, India, 2011; pp. 56–80. [Google Scholar]
- Samantaray, S.; Nayak, J.K.; Mallick, N. Wastewater Utilization for Poly-β-Hydroxybutyrate Production by the Cyanobacterium Aulosira fertilissima in a Recirculatory Aquaculture System. Appl. Environ. Microbiol. 2011, 77, 8735–8743. [Google Scholar] [CrossRef]
- Dawes, E.A.; Senior, P.J. The Role and Regulation of Energy Reserve Polymers in Micro-organisms. Adv. Microb. Physiol. 1973, 10, 135–266. [Google Scholar]
- Bhati, R.; Mallick, N. Poly(3-hydroxybutyrate-co-3-hydroxyvalerate) copolymer production by the diazotrophic cyanobacterium Nostoc muscorum Agardh: Process optimization and polymer characterization. Algal Res. 2015, 7, 78–85. [Google Scholar] [CrossRef]
- Panda, B.; Jain, P.; Sharma, L.; Mallick, N. Optimization of cultural and nutritional conditions for accumulation of poly-β-hydroxybutyrate in Synechocystis sp. PCC 6803. Bioresour. Technol. 2006, 97, 1296–1301. [Google Scholar] [CrossRef]
- Poirier, Y.; Nawrath, C.; Somerville, C. Production of Polyhydroxyalkanoates, a Family of Biodegradable Plastics and Elastomers, in Bacteria and Plants. Nat. Biotechnol. 1995, 13, 142–150. [Google Scholar] [CrossRef]
- Gibson, J. Movement of acetate across the cytoplasmic membrane of the unicellular cyanobacteria Synechococcus and Aphanocapsa. Arch. Microbiol. 1981, 130, 175–179. [Google Scholar] [CrossRef]
- Khatipov, E. Accumulation of poly-β-hydroxybutyrate by Rhodobacter sphaeroides on various carbon and nitrogen substrates. FEMS Microbiol. Lett. 1998, 162, 39–45. [Google Scholar] [CrossRef]
- Lee, I.Y.; Kim, M.K.; Chang, H.N.; Park, Y.H. Regulation of poly-β-hydroxybutyrate biosynthesis by nicotinamide nucleotides in Alcaligenes eutrophus. FEMS Microbiol. Lett. 1995, 131, 35–39. [Google Scholar]
- Sangkharak, K.; Prasertsan, P. Nutrient optimization for production of polyhydroxybutyrate from halotolerant photosynthetic bacteria cultivated under aerobic-dark condition. Electron. J. Biotechnol. 2008, 11, 83–94. [Google Scholar] [CrossRef]
- Tabandeh, F.; Vasheghani-Farahani, E. Biosynthesis of poly-β-hydroxybutyrate as a biodegradable polymer. Iran Polym. J. 2003, 12, 37–42. [Google Scholar]
- Mallick, N.; Gupta, S.; Panda, B.; Sen, R. Process optimization for poly(3-hydroxybutyrate-co-3-hydroxyvalerate) co-polymer production by Nostoc muscorum. Biochem. Eng. J. 2007, 37, 125–130. [Google Scholar] [CrossRef]
- Nishioka, M.; Nakai, K.; Miyake, M.; Asada, Y.; Taya, M. Production of poly-β-hydroxybutyrate by thermophilic cyanobacterium, Synechococcus sp. MA19, under phosphate-limited conditions. Biotechnol. Lett. 2001, 23, 1095–1099. [Google Scholar] [CrossRef]
- Suzuki, T.; Tsygankov, A.A.; Miyake, J.; Tokiwa, Y.; Asada, Y. Accumulation of polyhydroxybutyrate by a non-sulphur photosynthetic bacterium Rhodobacter spheroids RV at different pH. Biotech. Lett. 1995, 17, 395–400. [Google Scholar] [CrossRef]
- Karbasi, F.; Ardjmand, M.; Yunesi, H.; Kordi, A.S.; Yaghmaei, S. Investigation of optimum fermentation condition for PHA production by four species: Hydrogenophaga pseudoflava DSMZ 1034, Azohydromonas lata DSMZ 1123, Cupriavidus necator DSMZ 545, Azotobacter beijinckii DSMZ 1041. World Appl. Sci. J. 2011, 14, 36–47. [Google Scholar]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, M.K.; Rai, P.K.; Rai, A.; Singh, S.; Singh, J.S. Poly-β-Hydroxybutyrate Production by the Cyanobacterium Scytonema geitleri Bharadwaja under Varying Environmental Conditions. Biomolecules 2019, 9, 198. https://doi.org/10.3390/biom9050198
Singh MK, Rai PK, Rai A, Singh S, Singh JS. Poly-β-Hydroxybutyrate Production by the Cyanobacterium Scytonema geitleri Bharadwaja under Varying Environmental Conditions. Biomolecules. 2019; 9(5):198. https://doi.org/10.3390/biom9050198
Chicago/Turabian StyleSingh, Manoj K., Pradeep K. Rai, Anuradha Rai, Surendra Singh, and Jay Shankar Singh. 2019. "Poly-β-Hydroxybutyrate Production by the Cyanobacterium Scytonema geitleri Bharadwaja under Varying Environmental Conditions" Biomolecules 9, no. 5: 198. https://doi.org/10.3390/biom9050198
APA StyleSingh, M. K., Rai, P. K., Rai, A., Singh, S., & Singh, J. S. (2019). Poly-β-Hydroxybutyrate Production by the Cyanobacterium Scytonema geitleri Bharadwaja under Varying Environmental Conditions. Biomolecules, 9(5), 198. https://doi.org/10.3390/biom9050198