Inhibition of Rho GTPases in Invertebrate Growth Cones Induces a Switch in Responsiveness to Retinoic Acid

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Cell Culture Procedures
2.3. Chemicals
2.4. Growth Cone Turning Assays
2.5. Growth Cone Measurements
2.6. Data and Statistical Analysis
3. Results
3.1. The Inhibition of Cdc42 Induces a Switch in Growth Cone Responsiveness to AtRA, but not to 9-cis RA
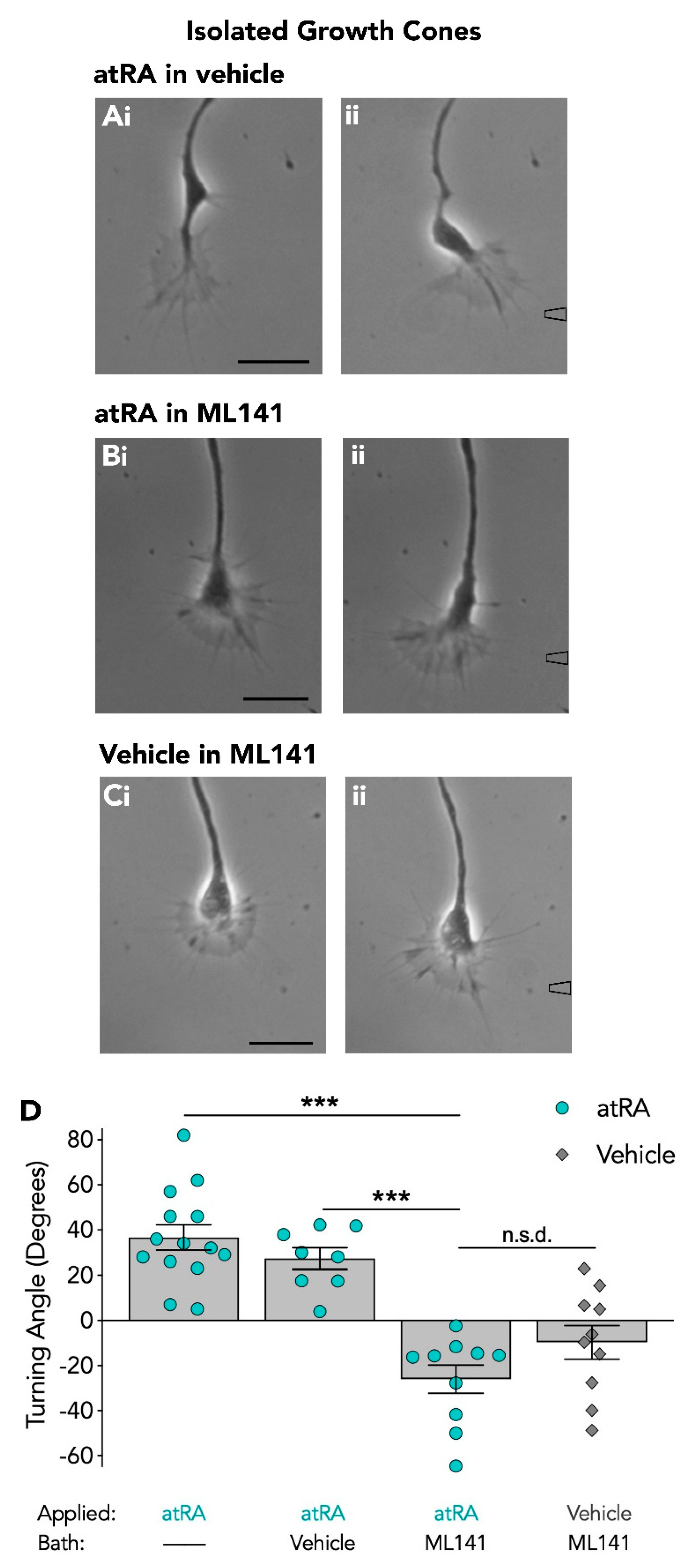
3.2. The Cdc42 Inhibitor Blocks Retinoic Acid-Induced Growth Cone Turning of Isolated Growth Cones
3.3. The Rac Inhibitor Induced a Switch in Growth Cone Responsiveness to AtRA, but not to 9-cis RA
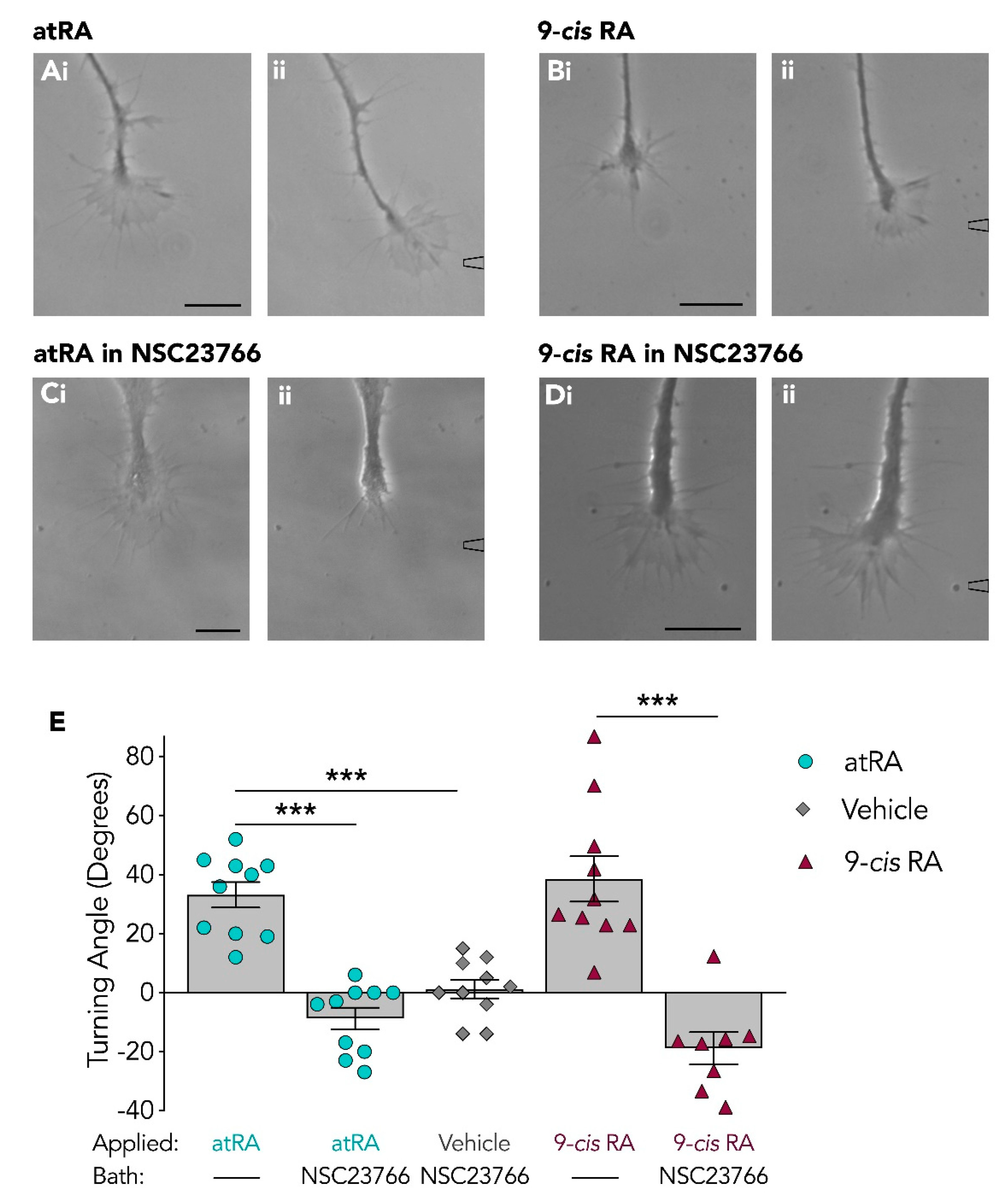
3.4. Isolated Growth Cones Continue to Turn Toward Retinoids Following Rac Inhibition
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Chen, Y.; Huang, L.; Solursh, M. A concentration gradient of retinoids in the early Xenopus laevis embryo. Dev. Biol. 1994, 161, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Zile, M.H. Vitamin A and embryonic development: An overview. J. Nutr. 1998, 128, 455S–458S. [Google Scholar] [CrossRef] [PubMed]
- Tsonis, P.A.; Tsavaris, M.; Call, M.K.; Chandraratna, R.A.S.; Del Rio-Tsonis, K. Expression and role of retinoic acid receptor alpha in lens regeneration. Dev. Growth Differ. 2002, 44, 391–394. [Google Scholar] [CrossRef] [PubMed]
- Maden, M.; Hind, M. Retinoic acid, a regeneration-inducing molecule. Dev. Dyn. 2003, 226, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Maden, M.; Gale, E.; Kostetskii, I.; Zile, M. Vitamin A-deficient quail embryos have half a hindbrain and other neural defects. Curr. Biol. 1996, 6, 417–426. [Google Scholar] [CrossRef] [Green Version]
- Corcoran, J.; So, P.-L.; Barber, R.D.; Vincent, K.J.; Mazarakis, N.D.; Mitrophanous, K.A.; Kingsman, S.M.; Maden, M. Retinoic acid receptor β2 and neurite outgrowth in the adult mouse spinal cord in vitro. J. Cell Sci. 2002, 115, 3779–3786. [Google Scholar] [CrossRef] [PubMed]
- Prince, D.J.; Carlone, R.L. Retinoic acid involvement in the reciprocal neurotrophic interactions between newt spinal cord and limb blastemas in vitro. Brain Res. Dev. Brain Res. 2003, 140, 67–73. [Google Scholar] [CrossRef]
- Dmetrichuk, J.M.; Spencer, G.E.; Carlone, R.L. Retinoic acid-dependent attraction of adult spinal cord axons towards regenerating newt limb blastemas in vitro. Dev. Biol. 2005, 281, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Dmetrichuk, J.M.; Carlone, R.L.; Spencer, G.E. Retinoic acid induces neurite outgrowth and growth cone turning in invertebrate neurons. Dev. Biol. 2006, 294, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Maghsoodi, B.; Poon, M.M.; Nam, C.I.; Aoto, J.; Ting, P.; Chen, L. Retinoic acid regulates RARα-mediated control of translation in dendritic RNA granules during homeostatic synaptic plasticity. PNAS 2008, 105, 16015–16020. [Google Scholar] [CrossRef]
- Ochoa, W.F.; Torrecillas, A.; Fita, I.; Verdaguer, N.; Corbalán-García, S.; Gomez-Fernandez, J.C. Retinoic acid binds to the C2-domain of protein kinase C(alpha). Biochemistry 2003, 42, 8774–8779. [Google Scholar] [CrossRef] [PubMed]
- Radominska-Pandya, A.; Chen, G.; Czernik, P.J.; Little, J.M.; Samokyszyn, V.M.; Carter, C.A.; Nowak, G. Direct interaction of all-trans-retinoic acid with protein kinase C (PKC). Implications for PKC signaling and cancer therapy. J. Biol. Chem. 2000, 275, 22324–22330. [Google Scholar] [CrossRef] [PubMed]
- Aoto, J.; Nam, C.I.; Poon, M.M.; Ting, P.; Chen, L. Synaptic signaling by all-trans retinoic acid in homeostatic synaptic plasticity. Neuron 2008, 60, 308–320. [Google Scholar] [CrossRef] [PubMed]
- Vesprini, N.D.; Spencer, G.E. Retinoic acid induces changes in electrical properties of adult neurons in a dose- and isomer-dependent manner. J. Neurophysiol. 2014, 111, 1318–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vesprini, N.D.; Dawson, T.F.; Yuan, Y.; Bruce, D.; Spencer, G.E. Retinoic acid affects calcium signaling in adult molluscan neurons. J. Neurophysiol. 2015, 113, 172–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrar, N.R.; Dmetrichuk, J.M.; Carlone, R.L.; Spencer, G.E. A novel, nongenomic mechanism underlies retinoic acid-induced growth cone turning. J. Neurosci. 2009, 29, 14136–14142. [Google Scholar] [CrossRef] [PubMed]
- Maden, M.; Keen, G.; Jones, G.E. Retinoic acid as a chemotactic molecule in neuronal development. Int. J. Dev. Neurosci. 1998, 16, 317–322. [Google Scholar] [CrossRef]
- Li, X.; Saint-Cyr-Proulx, E.; Aktories, K.; Lamarche-Vane, N. Rac1 and Cdc42 but not RhoA or Rho kinase activities are required for neurite outgrowth induced by the netrin-1 receptor DCC (deleted in colorectal cancer) in N1E-115 neuroblastoma cells. J. Biol. Chem. 2002, 277, 15207–15214. [Google Scholar] [CrossRef]
- Yuan, X.; Jin, M.; Xu, X.; Song, Y.; Wu, C.; Poo, M.; Duan, S. Signalling and crosstalk of Rho GTPases in mediating axon guidance. Nat. Cell Biol. 2003, 5, 38–45. [Google Scholar] [CrossRef]
- Wahl, S.; Barth, H.; Ciossek, T.; Aktories, K.; Mueller, B.K. Ephrin-A5 induces collapse of growth cones by activating Rho and Rho kinase. J. Cell Biol. 2000, 149, 263–270. [Google Scholar] [CrossRef]
- Jin, M.; Guan, C.; Jiang, Y.; Chen, G.; Zhao, C.; Cui, K.; Song, Y.; Wu, C.; Poo, M.; Yuan, X. Ca2+-dependent regulation of rho GTPases triggers turning of nerve growth cones. J. Neurosci. 2005, 25, 2338–2347. [Google Scholar] [CrossRef] [PubMed]
- Spencer, G.E.; Syed, N.I.; Lukowiak, K.; Winlow, W. Halothane-induced synaptic depression at both in vivo and in vitro reconstructed synapses between identified Lymnaea neurons. J. Neurophysiol. 1995, 74, 2604–2613. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.J.; Farrar, N.; Carlone, R.L.; Spencer, G.E. Developmental expression of a molluscan RXR and evidence for its novel, nongenomic role in growth cone guidance. Dev. Biol. 2010, 343, 124–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, C.J.; Rand, C.; Mohammad, I.; Lepp, A.; Vesprini, N.; Wiebe, O.; Carlone, R.; Spencer, G.E. Expression of a retinoic acid receptor (RAR)-like protein in the embryonic and adult nervous system of a protostome species. J. Exp. Zool. B Mol. Dev. Evol. 2015, 324, 51–67. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.G.; Hadley, R.D.; Kater, S.B.; Hauser, G.C. Neurite outgrowth in molluscan organ and cell cultures: The role of conditioning factor(s). J. Neurosci. 1981, 1, 1008–1021. [Google Scholar] [CrossRef] [PubMed]
- Hong, L.; Kenney, S.R.; Phillips, G.K.; Simpson, D.; Schroeder, C.E.; Nöth, J.; Romero, E.; Swanson, S.; Waller, A.; Strouse, J.J.; et al. Characterization of a Cdc42 protein inhibitor and its use as a molecular probe. J. Biol. Chem. 2013, 288, 8531–8543. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Zhang, Z.; Yuan, X.; Poo, M. Spatiotemporal dynamics of traction forces show three contraction centers in migratory neurons. J. Cell Biol. 2015, 209, 759–774. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Dickerson, J.B.; Guo, F.; Zheng, J.; Zheng, Y. Rational design and characterization of a Rac GTPase-specific small molecule inhibitor. Proc. Natl. Acad. Sci. USA 2004, 101, 7618–7623. [Google Scholar] [CrossRef] [Green Version]
- Hou, H.; Chávez, A.E.; Wang, C.-C.; Yang, H.; Gu, H.; Siddoway, B.A.; Hall, B.J.; Castillo, P.E.; Xia, H. The Rac1 inhibitor NSC23766 suppresses CREB signaling by targeting NMDA receptor function. J. Neurosci. 2014, 34, 14006–14012. [Google Scholar] [CrossRef]
- Kaufmann, N.; Wills, Z.P.; Van Vactor, D. Drosophila Rac1 controls motor axon guidance. Development 1998, 125, 453–461. [Google Scholar]
- Rajnicek, A.M.; Foubister, L.E.; McCaig, C.D. Temporally and spatially coordinated roles for Rho, Rac, Cdc42 and their effectors in growth cone guidance by a physiological electric field. J. Cell. Sci. 2006, 119, 1723–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dmetrichuk, J.M.; Carlone, R.L.; Jones, T.R.B.; Vesprini, N.D.; Spencer, G.E. Detection of endogenous retinoids in the molluscan CNS and characterization of the trophic and tropic actions of 9-cis retinoic acid on isolated neurons. J. Neurosci. 2008, 28, 13014–13024. [Google Scholar] [CrossRef] [PubMed]
- Dickson, B.J. Rho GTPases in growth cone guidance. Curr. Opin. Neurobiol. 2001, 11, 103–110. [Google Scholar] [CrossRef]
- Hall, A.; Lalli, G. Rho and Ras GTPases in axon growth, guidance, and branching. Cold Spring Harb. Perspect. Biol. 2010, 2. [Google Scholar] [CrossRef] [PubMed]
- Ruchhoeft, M.L.; Ohnuma, S.; McNeill, L.; Holt, C.E.; Harris, W.A. The neuronal architecture of Xenopus retinal ganglion cells is sculpted by rho-family GTPases in vivo. J. Neurosci. 1999, 19, 8454–8463. [Google Scholar] [CrossRef] [PubMed]
- Ng, J.; Nardine, T.; Harms, M.; Tzu, J.; Goldstein, A.; Sun, Y.; Dietzl, G.; Dickson, B.J.; Luo, L. Rac GTPases control axon growth, guidance and branching. Nature 2002, 416, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Lundquist, E.A. Rac proteins and the control of axon development. Curr. Opin, Neurobiol. 2003, 13, 384–390. [Google Scholar] [CrossRef]
- Dalpé, G.; Zhang, L.W.; Zheng, H.; Culotti, J.G. Conversion of cell movement responses to semaphorin-1 and plexin-1 from attraction to repulsion by lowered levels of specific Rac GTPases in C. elegans. Development 2004, 131, 2073–2088. [Google Scholar] [CrossRef] [PubMed]
- Ming, G.; Song, H.; Berninger, B.; Holt, C.E.; Tessier-Lavigne, M.; Poo, M. cAMP-dependent growth cone guidance by netrin-1. Neuron 1997, 19, 1225–1235. [Google Scholar] [CrossRef]
- Nishiyama, M.; Hoshino, A.; Tsai, L.; Henley, J.R.; Goshima, Y.; Tessier-Lavigne, M.; Poo, M.-M.; Hong, K. Cyclic AMP/GMP-dependent modulation of Ca2+ channels sets the polarity of nerve growth-cone turning. Nature 2003, 423, 990–995. [Google Scholar] [CrossRef]
- Song, H.J.; Ming, G.L.; Poo, M.M. cAMP-induced switching in turning direction of nerve growth cones. Nature 1997, 388, 275–279. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, K.L.; Mercurio, A.M. Protein kinase A regulates Rac and is required for the growth factor-stimulated migration of carcinoma cells. J. Biol. Chem. 2001, 276, 47895–47900. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, D.J.; Pujic, Z.; Goodhill, G.J. Calcium signaling in axon guidance. Trends Neurosci. 2014, 37, 424–432. [Google Scholar] [CrossRef] [PubMed]
- De Hoog, E.; Lukewich, M.K.; Spencer, G.E. Retinoic acid inhibits neuronal voltage-gated calcium channels. Cell Calcium 2018, 72, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Guirland, C.; Ming, G.-L.; Zheng, J.Q. A CaMKII/calcineurin switch controls the direction of Ca(2+)-dependent growth cone guidance. Neuron 2004, 43, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Donald, S.; Hill, K.; Lecureuil, C.; Barnouin, R.; Krugmann, S.; John Coadwell, W.; Andrews, S.R.; Walker, S.A.; Hawkins, P.T.; Stephens, L.R.; et al. P-Rex2, a new guanine-nucleotide exchange factor for Rac. FEBS Lett. 2004, 572, 172–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, C.; Diaz-Cuadros, M.; Chalfie, M. GEFs and Rac GTPases control directional specificity of neurite extension along the anterior-posterior axis. Proc. Natl. Acad. Sci. USA 2016, 113, 6973–6978. [Google Scholar] [CrossRef] [PubMed]
- Sayyad, W.A.; Fabris, P.; Torre, V. The role of Rac1 in the growth cone dynamics and force generation of DRG neurons. PLoS ONE 2016, 11, e0146842. [Google Scholar] [CrossRef]
- Sattler, R.; Tymianski, M.; Feyaz, I.; Hafner, M.; Tator, C.H. Voltage-sensitive calcium channels mediate calcium entry into cultured mammalian sympathetic neurons following neurite transection. Brain Res. 1996, 719, 239–246. [Google Scholar] [CrossRef]
- Govek, E.-E.; Newey, S.E.; Aelst, L.V. The role of the Rho GTPases in neuronal development. Genes Dev. 2005, 19, 1–49. [Google Scholar] [CrossRef] [Green Version]
- Bokoch, G.M. Biology of the p21-activated kinases. Annu. Rev. Biochem. 2003, 72, 743–781. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.; de Hoog, E.; Tolentino, M.; Nasser, T.; Spencer, G.E. Pharmacological evidence for the role of RAR in axon guidance and embryonic development of a protostome species. Genesis 2019, e23301. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Kim, S.-W.; Cheon, K.; Tabassam, F.H.; Yoon, J.-H.; Koo, J.S. Nonclassical action of retinoic acid on the activation of the cAMP response element-binding protein in normal human bronchial epithelial cells. Mol. Biol. Cell 2006, 17, 566–575. [Google Scholar] [CrossRef] [PubMed]
- Bouzigues, C.; Morel, M.; Triller, A.; Dahan, M. Asymmetric redistribution of GABA receptors during GABA gradient sensing by nerve growth cones analyzed by single quantum dot imaging. Proc. Natl. Acad. Sci. USA 2007, 104, 11251–11256. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, A.; Nasser, T.I.N.; Spencer, G.E. Inhibition of Rho GTPases in Invertebrate Growth Cones Induces a Switch in Responsiveness to Retinoic Acid. Biomolecules 2019, 9, 460. https://doi.org/10.3390/biom9090460
Johnson A, Nasser TIN, Spencer GE. Inhibition of Rho GTPases in Invertebrate Growth Cones Induces a Switch in Responsiveness to Retinoic Acid. Biomolecules. 2019; 9(9):460. https://doi.org/10.3390/biom9090460
Chicago/Turabian StyleJohnson, Alysha, Tamara I. N. Nasser, and Gaynor E. Spencer. 2019. "Inhibition of Rho GTPases in Invertebrate Growth Cones Induces a Switch in Responsiveness to Retinoic Acid" Biomolecules 9, no. 9: 460. https://doi.org/10.3390/biom9090460
APA StyleJohnson, A., Nasser, T. I. N., & Spencer, G. E. (2019). Inhibition of Rho GTPases in Invertebrate Growth Cones Induces a Switch in Responsiveness to Retinoic Acid. Biomolecules, 9(9), 460. https://doi.org/10.3390/biom9090460