Spatial Analysis of Digital Imagery of Weeds in a Maize Crop

 ,
, 
Abstract
:1. Introduction
Our Investigation
- Greig Smith’s [24] test of departure from randomness,
- standard geostatistical variography of quadrat counts and
- wavelet analysis.
2. Materials and Methods
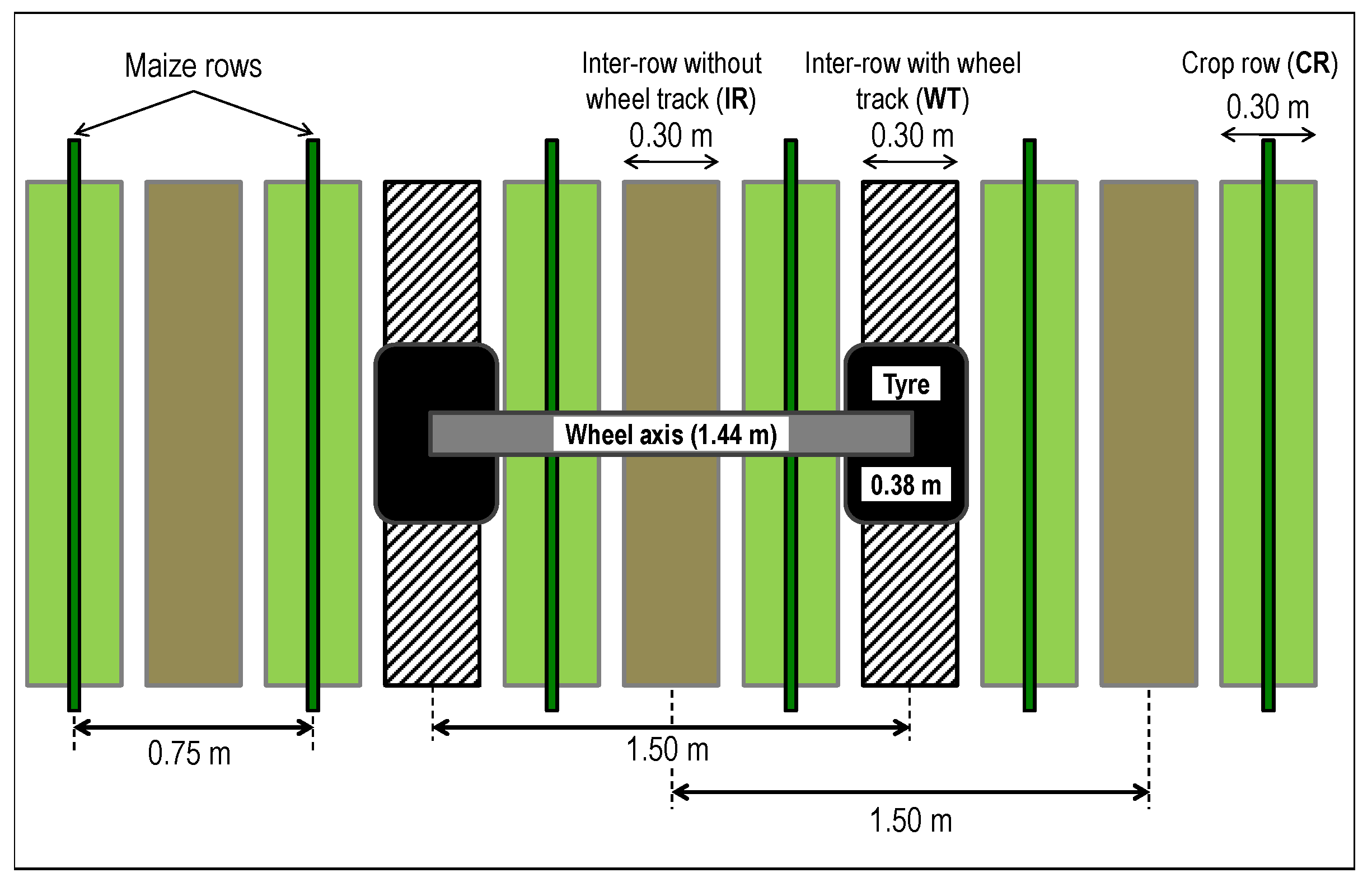
2.1. Field of Study
2.2. Weed Imaging
2.3. Statistical Analysis
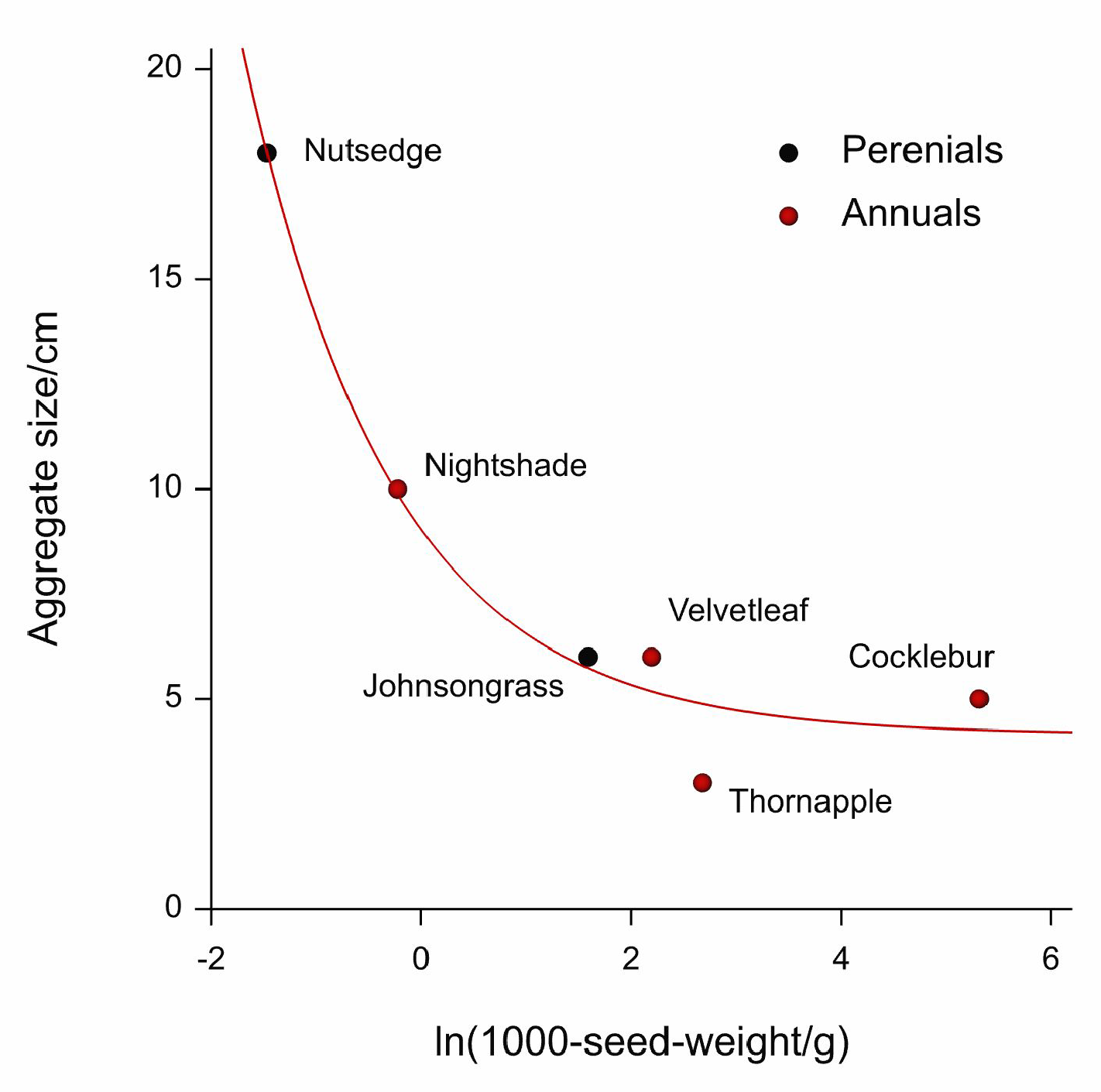
2.3.1. Aggregation Analysis
- (1)
- inter-rows without wheel tracks (IR);
- (2)
- inter-rows with wheel tracks (WT); and
- (3)
- crop rows (CR).
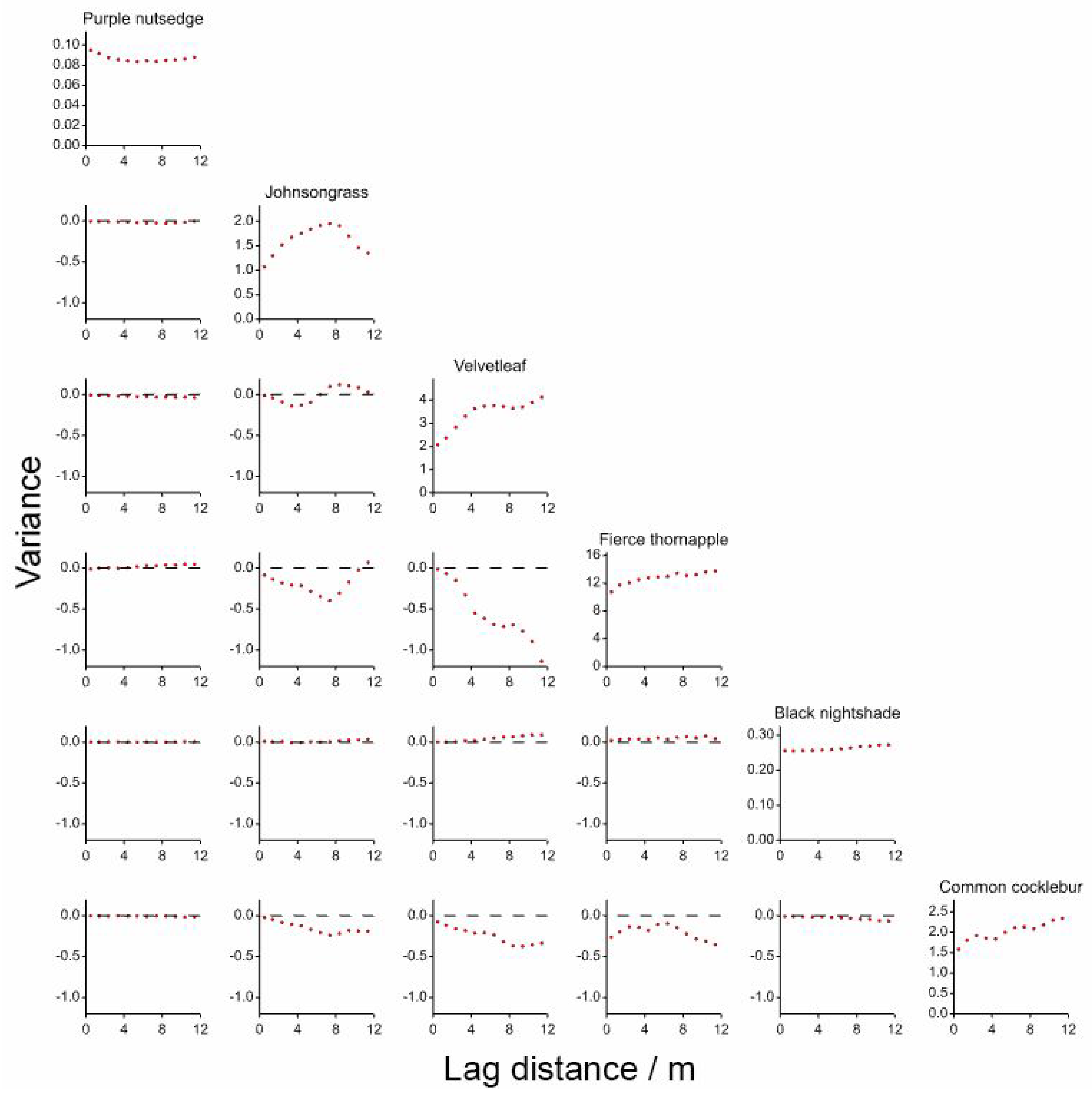
2.3.2. Geostatistical Analysis
2.3.3. Wavelet Analysis
3. Results and Discussion
3.1. Aggregation Analysis
3.2. Geostatistical Analysis
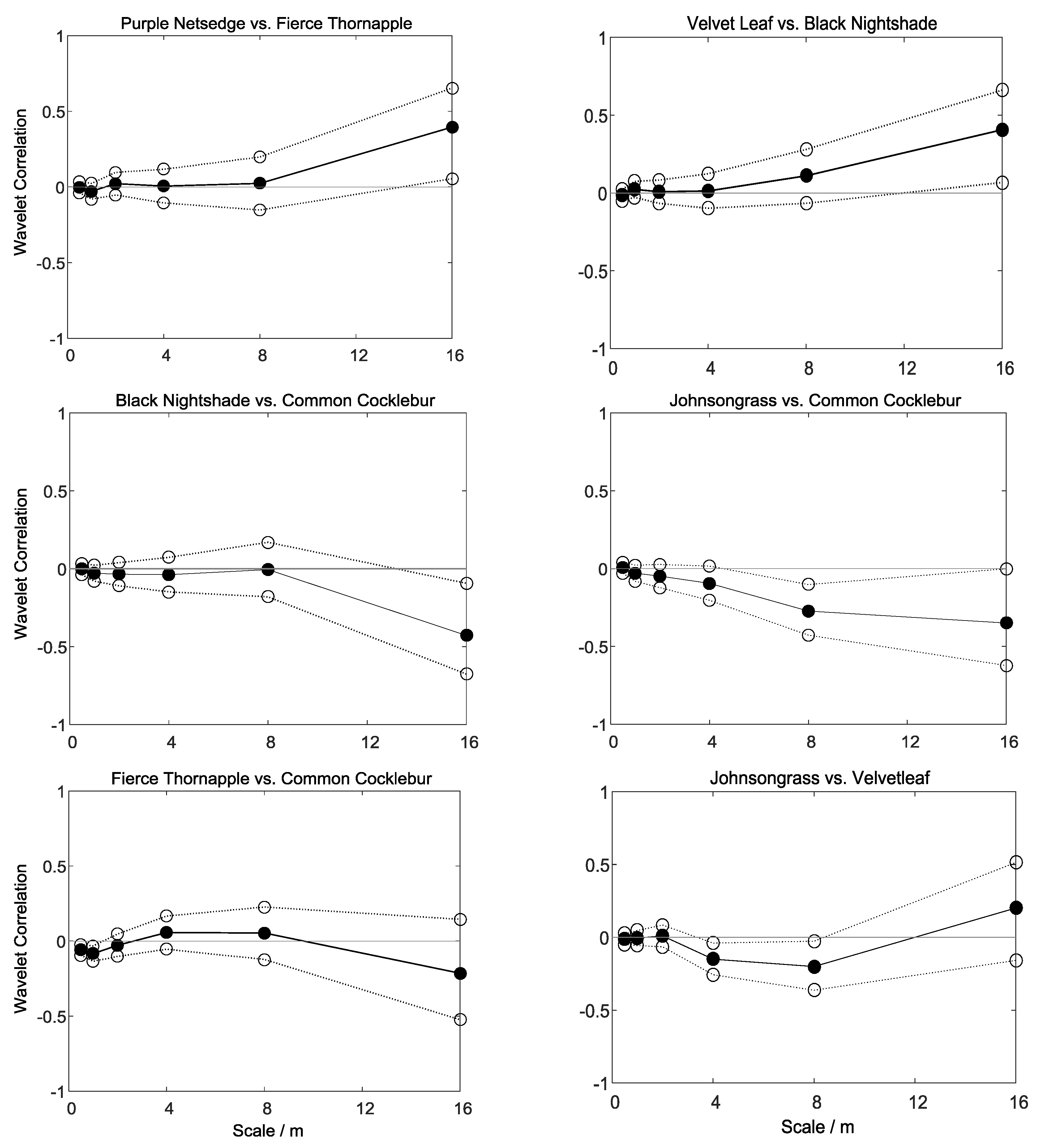
3.3. Wavelet Analysis
- (1)
- There were significant positive correlations between the pair nutsedge and thornapple and the pair velvetleaf and nightshade at the coarsest scale (16–32 m), suggesting that these pairs of weeds grow in similar regions of the plot (Figure 9a,b).
- (2)
- There were significant negative correlations between nightshade and cocklebur and between johnsongrass and cocklebur at the coarser scales (Figure 9c,d); it seems that the weeds in each pair exclude one another spatially, being located in different regions in the crop.
- (3)
- A weak, but significant negative correlation between thornapple and cocklebur appears at the finest scales (0.5–2 m); this suggests that there is local competition between these species (Figure 9e).
- (4)
- For the pairs velvetleaf and johnsongrass, velvetleaf and cocklebur, and velvetleaf and thornapple, we observed weak, but significant negative correlations at the mid-range scales (4–16 m) (for example, Figure 9f).
- (5)
- All other pairs had weak correlations across the scales that were not significantly different from zero.
4. Implications
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Maxwell, B.D.; Luschei, E.C. Justification for site-specific weed management based on ecology and economics. Weed Sci. 2005, 53, 221–227. [Google Scholar] [CrossRef]
- San Martıín, C.; Andújar, D.; Barroso, J.; Fernández-Quintanilla, C.; Dorado, J. Weed decision threshold as a key factor for herbicide reductions in site-specific weed management. Weed Technol. 2016, 30, 888–897. [Google Scholar] [CrossRef]
- Stoate, C.; Boatman, N.D.; Borralho, R.J.; Rio Carvalho, C.; de Snoo, G.R.; Eden, P. Ecological impacts of arable intensification in Europe. J. Environ. Manag. 2001, 63, 337–365. [Google Scholar] [CrossRef]
- Andújar, D.; Ribeiro, Á.; Fernández-Quintanilla, C.; Dorado, J. Accuracy and feasibility of optoelectronic sensors for weed mapping in wide row crops. Sensors 2011, 11, 2304–2318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgos-Artizzu, X.P.; Ribeiro, A.; Guijarro, M.; Pajares, G. Real-time image processing for crop/weed discrimination in maize fields. Comput. Electron. Agric. 2011, 75, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Christensen, S.; Søgaard, H.T.; Kudsk, P.; Nørremark, M.; Lund, I.; Nadimi, E.S.; Jørgensen, R. Site-specific weed control technologies. Weed Res. 2009, 49, 233–241. [Google Scholar] [CrossRef]
- Herrera, P.J.; Dorado, J.; Ribeiro, Á. A novel approach for weed type classification based on shape descriptors and a fuzzy decision-making method. Sensors 2014, 14, 15304–15324. [Google Scholar] [CrossRef] [PubMed]
- Dieleman, J.A.; Mortensen, D.A.; Buhler, D.D.; Cambardella, C.A.; Moorman, T.B. Identifying associations among site properties and weed species abundance. I. Multivariate analysis. Weed Sci. 2000, 48, 567–575. [Google Scholar] [CrossRef]
- Dieleman, J.A.; Mortensen, D.A.; Buhler, D.D.; Ferguson, R.B. Identifying associations among site properties and weed species abundance. II. Hypothesis generation. Weed Sci. 2000, 48, 576–587. [Google Scholar] [CrossRef]
- Rew, L.J.; Cousens, R.D. Spatial distribution of weeds in arable crops: Are current sampling and analytical methods appropriate? Weed Res. 2001, 41, 1–18. [Google Scholar] [CrossRef]
- Metcalfe, H.; Milne, A.E.; Webster, R.; Lark, R.M.; Murdoch, A.J.; Storkey, J. Designing a sampling scheme to reveal correlations between weeds and soil properties at multiple spatial scales. Weed Res. 2016, 56, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benech-Arnold, R.L.; Sanchez, R.A.; Forcella, F.; Kruk, B.C.; Ghersa, C.M. Environmental control of dormancy in weed seed banks in soil. Weed Sci. 2000, 67, 105–122. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Singh, R.G.; Mahajan, G. Ecology and management of weeds under conservation agriculture: A review. Crop Prot. 2012, 38, 57–65. [Google Scholar] [CrossRef]
- Gallagher, R.; Cardina, J. The effect of light environment during tillage on the recruitment of the various summer annuals. Weed Sci. 1998, 46, 214–216. [Google Scholar]
- Tardif-Paradis, C.; Simard, M.-J.; Leroux, G.D.; Panneton, B.; Nurse, R.E.; Vanasse, A. Effect of planter and tractor wheels on row and inter-row weed populations. Crop Prot. 2015, 71, 66–71. [Google Scholar] [CrossRef]
- Longchamps, L.; Panneton, B.; Simard, M.-J.; Leroux, G.D. Could weed sensing in corn interrows result in efficient weed control? Weed Technol. 2012, 26, 649–656. [Google Scholar] [CrossRef]
- Andújar, D.; Ruiz, D.; Ribeiro, Á.; Fernández-Quintanilla, C.; Dorado, J. Spatial distribution patterns of Johnsongrass (Sorghum halepense) in corn fields in Spain. Weed Sci. 2011, 59, 82–89. [Google Scholar] [CrossRef]
- Ruiz, D.; Escribano, C.; Fernández-Quintanilla, C. Assessing the opportunity for site-specific management of Avena sterilis in winter barley fields in Spain. Weed Res. 2006, 46, 379–387. [Google Scholar] [CrossRef]
- Howard, C.L.; Mortimer, A.M.; Gould, P.; Putwain, P.D.; Cousens, R.; Cussans, G.W. The dispersal of weeds: Seed movement in arable agriculture. In Proceedings of Brighton Crop Protection Conference–Weeds; British Crop Protection Council: Brighton, UK, 1991; pp. 821–828. [Google Scholar]
- Paice, M.E.R.; Day, W.; Rew, L.J.; Howard, A. A stochastic simulation model for evaluating the concept of patch spraying. Weed Res. 1998, 38, 373–388. [Google Scholar] [CrossRef]
- Boyd, N.; Van Acker, R. Seed and microsite limitations to emergence of four annual weed species. Weed Sci. 2004, 52, 571–577. [Google Scholar] [CrossRef]
- Royal Botanic Gardens Kew. Seed Information Database (SID). Version 7.1. Available online: http://data.kew.org/sid/ (accessed on 8 February 2018).
- San Martín, C.; Andújar, D.; Fernández-Quintanilla, C.; Dorado, J. Spatio-temporal dynamics of Sorghum halepense in poplar short-rotation coppice under several vegetation management systems. Forest Ecol. Manag. 2016, 379, 37–49. [Google Scholar] [CrossRef]
- Greig-Smith, P. The use of random and contiguous quadrats in the study of the structure of plant communities. Ann. Bot. 1952, 16, 293–316. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO). World Reference Base for Soil Resources. In A Framework for International Classification, Correlation and Communication; World Soil Resources Reports 103; Food and Agriculture Organization (FAO): Rome, Italy, 2006. [Google Scholar]
- Environmental Systems Research Institute (ESRI). ArcGIS Desktop: Release 10.1; Environmental Systems Research Institute: Redlands, CA, USA, 2013. [Google Scholar]
- Matérn, B. Spatial variation: stochastic models and their application to problems in forest surveys and other sampling investigations. Meddelanden från Statens Skogforskinstitut 1960, 49, 1–144. [Google Scholar]
- Webster, R.; Oliver, M.A. Geostatistics for Environmental Scientists, 2nd ed.; John Wiley & Sons: Chichester, UK, 2007. [Google Scholar]
- Payne, R.W. (Ed.) The Guide to GenStat Release 18—Part 2: Statistics; VSN International: Hemel Hempstead, UK, 2016. [Google Scholar]
- Percival, D.B.; Walden, A.T. Wavelet Methods for Time Series Analysis; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Milne, A.E.; Webster, R.; Lark, R.M. Spectral and wavelet analysis of gilgai patterns from air photography. Aus. J. Soil Res. 2010, 48, 309–325. [Google Scholar] [CrossRef]
- Lark, R.M.; Webster, R. Changes in variance and correlation of soil properties with scale and location: Analysis using an adapted maximal overlap discrete wavelet transform. Eur. J. Soil Sci. 2001, 52, 547–562. [Google Scholar] [CrossRef]
- Whitcher, B.; Guttorp, P.; Percival, D.B. Wavelet analysis of covariance with application to atmospheric time series. J. Geophys. Res. Atmos. 2000, 105, 14941–14962. [Google Scholar] [CrossRef]
- Daubechies, I. Orthonormal bases of compactly supported wavelets. Comm. Pure Appl. Math. 1988, 41, 909–996. [Google Scholar] [CrossRef]
- Heijting, S.; Van der Werf, W.; Kruijer, W.; Stein, A. Testing the spatial significance of weed patterns in arable land using Mead’s test. Weed Res. 2007, 47, 396–405. [Google Scholar] [CrossRef]
- Cardina, J.; Johnson, G.A.; Sparrow, D.H. The nature and consequence of weed spatial distribution. Weed Sci. 1997, 45, 364–373. [Google Scholar]
- San Martín, C.; Andújar, D.; Fernández-Quintanilla, C.; Dorado, J. Spatial distribution patterns of weed communities in corn fields of Central Spain. Weed Sci. 2015, 63, 936–945. [Google Scholar] [CrossRef]
- Colbach, N.; Roger-Estrade, J.; Chauvel, B.; Caneill, J. Modeling vertical and lateral seed bank movements during mouldboard ploughing. Eur. J. Agron. 2000, 13, 111–124. [Google Scholar] [CrossRef]
- Brown, A.D.; Dexter, A.R.; Chamen, W.C.T.; Spoor, G. Effect of soil macroporosity and aggregate size on seed-soil contact. Soil Tillage Res. 1996, 38, 203–216. [Google Scholar] [CrossRef]
- Jurik, T.M.; Zhang, S. Tractor wheel traffic effects on weed emergence in central Iowa. Weed Technol. 1999, 13, 741–746. [Google Scholar]
- Theisen, G.; Bastiaans, L. Low disturbance seeding suppresses weeds in no-tillage soyabean. Weed Res. 2015, 55, 598–608. [Google Scholar] [CrossRef]
- Ballaré, C.; Scopel, A.; Sánchez, R. Foraging for light: Photosensory ecology and agricultural implications. Plant Cell Environ. 1997, 20, 820–825. [Google Scholar] [CrossRef]
- Norsworthy, J.K. Soybean canopy formation effects on pitted morningglory (Ipomoea lacunosa), common cocklebur (Xanthium strumarium), and sicklepod (Senna obtusifolia) emergence. Weed Sci. 2004, 52, 954–960. [Google Scholar] [CrossRef]
- Barroso, J.; Andújar, D.; San Martín, C.; Fernández-Quintanilla, C.; Dorado, J. Johnsongrass (Sorghum halepense) seed dispersal in corn crops under Mediterranean conditions. Weed Sci. 2012, 60, 34–41. [Google Scholar] [CrossRef]
- Andújar, D.; Barroso, J.; Fernández-Quintanilla, C.; Dorado, J. Spatial and temporal dynamics of Sorghum halepense patches in maize crops. Weed Res. 2012, 52, 411–1420. [Google Scholar] [CrossRef]
- Benech-Arnold, R.L.; Sanchez. Handbook of Seed Physiology. Application to Agriculture; Food Products Press: Binghamton, NY, USA, 2004; 480p. [Google Scholar]
- Dexter, A. Advances in characterization of soil structure. Soil Tillage Res. 1988, 11, 199–238. [Google Scholar] [CrossRef]
- Głąb, L.; Sowiński, J.; Bough, R.; Dayan, F.E. Allelopathic potential of sorghum (Sorghum bicolor (L.) Moench) in weed control: a comprehensive review. Adv. Agron. 2017, 145, 44–95. [Google Scholar]
- Calviño-Cancela, M. Seed and microsite limitations of recruitment and the impacts of post-dispersal seed predation at the within population level. Plant Ecol. 2007, 192, 35–44. [Google Scholar] [CrossRef]
- Scott, A.J.; Morgan, J.W. Dispersal and microsite limitation in Australian old fields. Oecologia 2012, 170, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Borgy, B.; Perronne, R.; Kohler, C.; Grison, A.-L.; Amiaud, B.; Gaba, S. Changes in functional diversity and intraspecific trait variability of weeds. Weed Res. 2016, 56, 102–113. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Biotype | Fruit Type | Seed Size/mm | Means of Dispersal | 1000 Seed Weight/g |
|---|---|---|---|---|---|
| Nutsedge | Perennial | Achene | 1.2–1.5 × 0.4–0.7 | Vegetative (rhizomes) | 0.23 |
| Johnsongrass | Perennial | Achene | 4.5 × 2 | Vegetative (rhizomes) | 4.9 |
| Velvetleaf | Annual | Capsule | 2–3 × 2 | Gravity and agriculture | 9.0 |
| Thornapple | Annual | Capsule | 3.8–4.6 × 3.0–3.8 | Gravity and agriculture | 14.5 |
| Nightshade | Annual | Berry | 1.5–2.0 × 1.4–1.8 | Zoochory | 0.8 |
| Cocklebur | Annual | Bur | 5–15 × 5–7 | Zoochory and agriculture | 203.4 |
| Species | Aggregate Size/cm | Variance:Mean |
|---|---|---|
| Nutsedge | 18 | 1.25 |
| Johnsongrass | 6 | 1.25 |
| Velvetleaf | 6 | 1.09 |
| Thornapple | 3 | 1.09 |
| Nightshade | 10 | 1.33 |
| Cocklebur | 5 | 1.20 |
| Species | Direction * | Model | A | /m | ** | ||||
|---|---|---|---|---|---|---|---|---|---|
| Johnsongrass | 0 | Periodic | 1.147 | 0.601 | 8.82 | ||||
| 90 | Periodic | 0.993 | 0.347 | 1.29 | |||||
| Velvetleaf | 0 | Periodic | 2.061 | 1.592 | 7.28 | ||||
| 90 | Periodic | 2.101 | 0.616 | 4.32 | |||||
| Thornapple | 0 | Exponential | 10.49 | 3.124 | 3.38 | ||||
| 90 | Periodic | 11.91 | 2.451 | 1.29 | |||||
| Nightshade | 0 | Power | 0.255 | 1.75 | 0.000271 | ||||
| 90 | Periodic | 0.260 | 0.0164 | 1.37 | |||||
| Cocklebur | 0 | Exponential | 1.551 | 0.938 | 0.070 | 7.84 | 5.24 | ||
| 90 | +Periodic | 1.597 | 0.299 | 0.025 | 2.83 | 1.11 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
San Martín, C.; Milne, A.E.; Webster, R.; Storkey, J.; Andújar, D.; Fernández-Quintanilla, C.; Dorado, J. Spatial Analysis of Digital Imagery of Weeds in a Maize Crop. ISPRS Int. J. Geo-Inf. 2018, 7, 61. https://doi.org/10.3390/ijgi7020061
San Martín C, Milne AE, Webster R, Storkey J, Andújar D, Fernández-Quintanilla C, Dorado J. Spatial Analysis of Digital Imagery of Weeds in a Maize Crop. ISPRS International Journal of Geo-Information. 2018; 7(2):61. https://doi.org/10.3390/ijgi7020061
Chicago/Turabian StyleSan Martín, Carolina, Alice E. Milne, Richard Webster, Jonathan Storkey, Dionisio Andújar, Cesar Fernández-Quintanilla, and José Dorado. 2018. "Spatial Analysis of Digital Imagery of Weeds in a Maize Crop" ISPRS International Journal of Geo-Information 7, no. 2: 61. https://doi.org/10.3390/ijgi7020061
APA StyleSan Martín, C., Milne, A. E., Webster, R., Storkey, J., Andújar, D., Fernández-Quintanilla, C., & Dorado, J. (2018). Spatial Analysis of Digital Imagery of Weeds in a Maize Crop. ISPRS International Journal of Geo-Information, 7(2), 61. https://doi.org/10.3390/ijgi7020061