1. Introduction
Disruption of secondary palate development will lead to cleft palate, a common birth defect that affects 1:700 births [
1]. The formation of the continuous secondary palate is a complex developmental process involving a series of steps, such as palate specification and initiation, vertical palatal growth, elevation and fusion [
2,
3]. During mouse embryogenesis and organogenesis, neural crest-derived mesenchymal cells and cranial ectoderm-originated epithelial cells form a cluster of bilateral facial primordia, including the two maxillary processes [
2,
4]. On Embryonic Day 11.5 (E11.5), a group of cells within these two maxillary processes are specified to become palatal mesenchymal cells, protrude into the primitive oral-nasal cavity and form two palatal outgrowths there [
5]. On E12.5, the palatal outgrowth further develops to form the characteristic palate shelf that contains a core mass of mesenchymal cells enclosed by epithelial cells [
1,
4,
5].
Vertical growth of the palatal shelves along the lateral aspects of the tongue continues into E13.5. On E14.5, however, the vertically-orientated palatal shelves undergo re-orientation and position themselves horizontally above the tongue, a process termed palate re-orientation or elevation [
1,
2,
3]. The two re-orientated palatal shelves grow horizontally towards each other to meet along the facial midline. This contact induces the merging of the medial edge epithelium (MEE) of the two shelves to form the medial edge seam (MES) that will subsequently undergo degeneration, leading to mesenchymal confluence and the formation of a continuous palate, a process called palate fusion. The resulting continuous palate separates the primitive oral-nasal cavity into nasal and oral cavities [
1,
2,
3].
The aforementioned processes have to be precisely regulated at the gene function level to assure the proper formation of secondary palate. Gene targeting technology has greatly advanced our understanding of the gene functions in palate growth, re-orientation and fusion, since it allows investigators to create mutations virtually in any genes of interest to study their function during development
in vivo [
1,
3,
6]. However, the mutant phenotype is often influenced by strain background. The effect of strain background on gene targeting was first reported in 1995 on the EGF receptor gene (
Egfr) knock-out [
7,
8]. Loss of
Egfr function leads to peri-implantation lethality on a CF-1 background [
8], death at mid-gestation stage on the 129 background [
7,
8], lethality at birth on C57B6 [
7] and post-natal lethality around 20 days of birth on MF-1 and CD-1 strain backgrounds [
7,
8]. Further studies uncovered a strain-dependent neurodegeneration defect in the
Egfr knock-out mice [
9]. Since then, the effect of strain background on knock-out mice has been supported by numerous studies and has become the consensus of the field [
10]. The effect of strain background on gene function during mouse secondary palate development has also been described recently [
11].The mechanisms underlying the strain effects are not yet well understood.
In contrast to gene function studies, little effort has been made to analyze the effects of strain background on gene expression during mouse embryogenesis, either in secondary palate formation or in embryonic development in general. We reasoned the possibility that different strains may bear sequence variation in a gene regulatory region, which could, in principle, affect its expression, and decided to test this idea by searching for genes that give distinct expression patterns on different strain backgrounds. As a result of this effort, we reported in this study that Meox-2 and Fgfr1 displayed different expression patterns in mouse secondary palate development in C57B6, Black Swiss and Swiss Webster, three strains commonly used in mouse developmental biology studies. This is the first study reporting a given gene that displays distinct expression patterns, not levels, on different strain background during embryonic development.
3. Results and Discussion
Meox-1 and
Meox-2 are two related mouse homeobox genes first discovered to be expressed specifically in paraxial mesoderm, such as somites, during development [
12]. Subsequent studies revealed
Meox-2 expression in mouse secondary palate mesenchymal cells, and the expression specifically marks the posterior soft palate region in the C57B6 strain [
13,
17]. Loss of
Meox-2 function in C57B6 mice leads to a posterior cleft palate in 10%–20% of mutant embryos due to a post-fusion defect [
14,
18,
19]. The present study explores whether strain background can affect
Meox-2 expression during mouse secondary palate development. Indeed,
Meox-2 displays distinct expression patterns on different strain backgrounds.
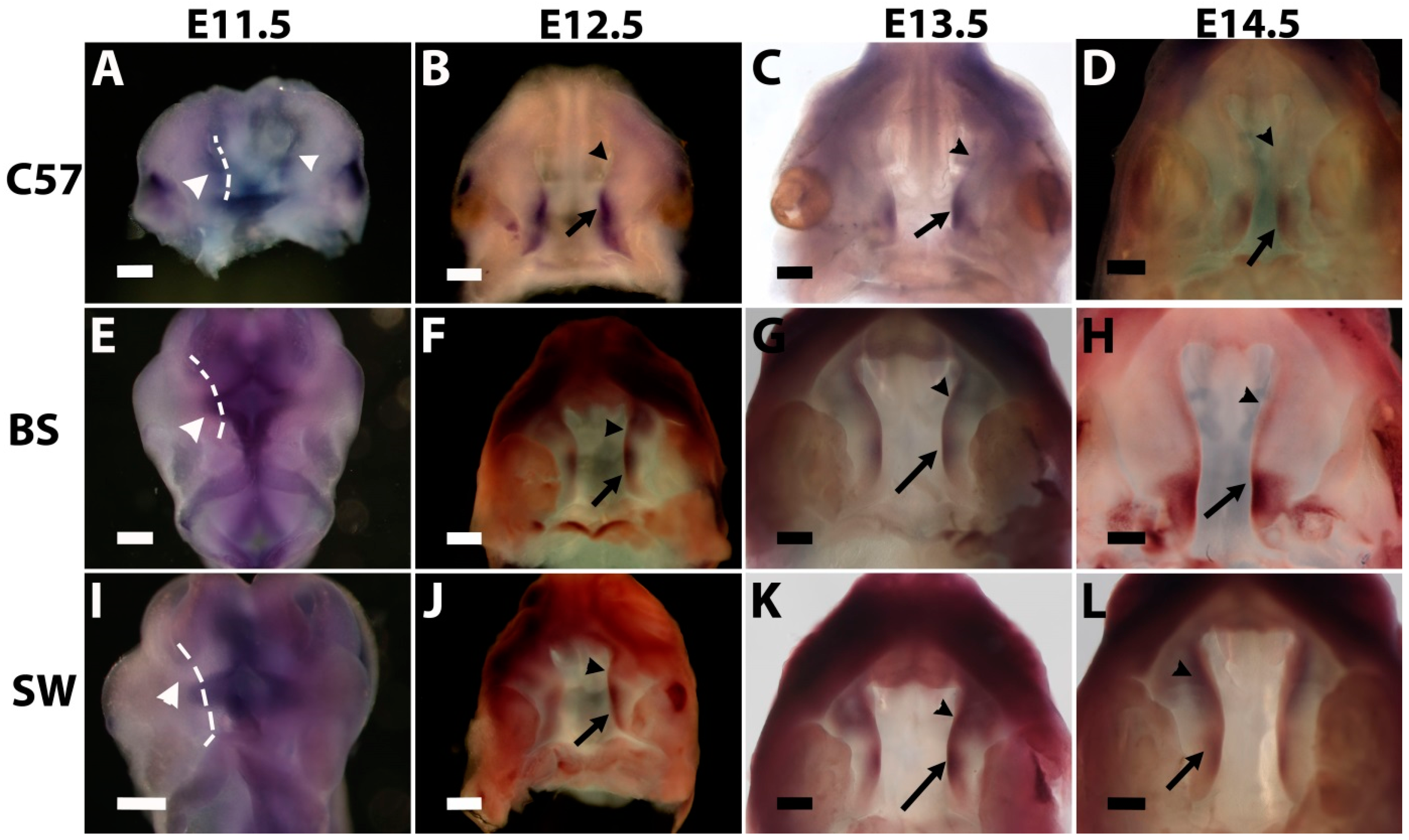
As mentioned above, palatogenesis is initiated by the formation of an outgrowth out of the maxillary process on E11.5 [
4,
5]. On the C57B6 strain background,
Meox-2 expression can be detected as early as E11.5 in the palate outgrowth covering both anterior and posterior regions (
Figure 1A). On E12.5, the expression of
Meox-2 is present only in the posterior two-thirds of the palate shelf (arrow in
Figure 1B), but absent in the anterior region (arrowhead in
Figure 1B). On E13.5, the expression further shifts to the posterior one-third of the palate shelf (arrow in
Figure 1C) and is completely absent in the anterior two-thirds (arrowhead in
Figure 1C). On E14.5, the expression is restricted to the posterior region corresponding to the soft palate (arrow in
Figure 1D). No expression is found in the anterior hard palate (arrowhead in
Figure 1D). This expression pattern of
Meox-2 confirmed the previous study using C57B [
13,
14].
On the Swiss Webster background, however, the expression of
Meox-2 is not detectable on E11.5 in the palate outgrowth (
Figure 1I). On E12.5, unlike C57B6,
Meox-2 is expressed along the entire palate A-P axis, although the intensity is stronger in the posterior region (arrow in
Figure 1J) than the anterior region (arrowhead in
Figure 1J). From E13.5, the intensity of anterior expression (arrowhead in
Figure 1K) is similar to that in the posterior expression (arrow in
Figure 1K). On E14.5,
Meox-2 expression covers both anterior and posterior regions with the same intensity (arrowhead and arrow in
Figure 1L).
On the Black Swiss background,
Meox-2 expression is not detectable in the palate outgrowth on E11.5 (
Figure 1E). On E12.5 and E13.5, expression is found in both the anterior and posterior regions, but the posterior expression level (arrow in
Figure 1F,G) is higher than the anterior expression (arrowhead in
Figure 1F,G). On E14.5, the expression in the posterior soft palate is increased and expanded (arrow in
Figure 1H), whereas the expression in the anterior hard palate is further reduced and narrowed, but not absent (arrowhead in
Figure 1H).
Post-hybridization sectioning of these embryos confirmed the previous studies [
13,
14] showing that the expression is restricted to the mesenchymal cells [
20].
Therefore, the expression of Meox-2 in secondary palate development is strain dependent. It is expressed on E11.5 only in the C57B6 strain, but not in Black Swiss and Swiss Webster strains. From E12.5 to E14.5, the expression of Meox-2 shows strong A-P polarity on the C57B6 background, since the expression is restricted to the posterior region and completely absent in the anterior region. The expression on the Swiss Webster background shows no A-P polarity on E14.5, although the expression on E12.5 and E13.5 is stronger in the posterior region; the expression is the same intensity from anterior to posterior on E14.5. The expression on the Black Swiss background on E14.5 is in an intermediate category between the C57B6 and Swiss Webster backgrounds; the posterior expression is strong in the posterior, whereas the anterior expression is weak and narrow, but not absent.
Figure 1.
The expression of Meox-2 during mouse secondary palate development on the backgrounds of C57B6 (A–D), Black Swiss (E–H) and Swiss Webster (I–L). Dashed lines in (E) and (I) indicate the unstained palate outgrowth areas. Arrows and arrowheads indicate to different regions in palate shelves, as illustrated in the Results and Discussion section. Scale bars represent 228 μm (B, D, F, J) and 285 µm (A, C, E, G–I, K and L); C57, BS and SW are the abbreviations for C57B6, Black Swiss and Swiss Webster.
Figure 1.
The expression of Meox-2 during mouse secondary palate development on the backgrounds of C57B6 (A–D), Black Swiss (E–H) and Swiss Webster (I–L). Dashed lines in (E) and (I) indicate the unstained palate outgrowth areas. Arrows and arrowheads indicate to different regions in palate shelves, as illustrated in the Results and Discussion section. Scale bars represent 228 μm (B, D, F, J) and 285 µm (A, C, E, G–I, K and L); C57, BS and SW are the abbreviations for C57B6, Black Swiss and Swiss Webster.
Since the
Meox-2 mutant line was studied only on the C57B6 background, the biological significance of this expression divergence among the three strains is not clear. However, all three strains express
Meox-2 in the posterior soft palate, and only Swiss Webster expresses the gene in both the anterior and posterior with the same intensity, suggesting that
Meox-2 is likely to be more important in the posterior. Consistent with this speculation, the cleft palates found in
Meox-2 mutant embryos on the C57B6 background are posterior clefts [
14]. In addition,
Meox-2 has been shown to play important roles in muscle differentiation during mouse embryonic development [
18,
19], and soft palate is composed mainly of muscle [
2].
The function of the FGF receptor 2 gene (
Fgfr2) in mouse secondary palate development has been studied by both loss-of-function and gain-of-function approaches. [
21,
22]. However, the function of
Fgfr1 in secondary palate has not been reported, since the
Fgfr1 null mutant embryos are early lethal due to gastrulation defects [
15]. Conditional deletion of the
Fgfr1 gene in mouse secondary palate has not yet been published. In the current study, we found that the
Fgfr1 gene shows divergent expression patterns among different strains during palate development, especially at early stages.
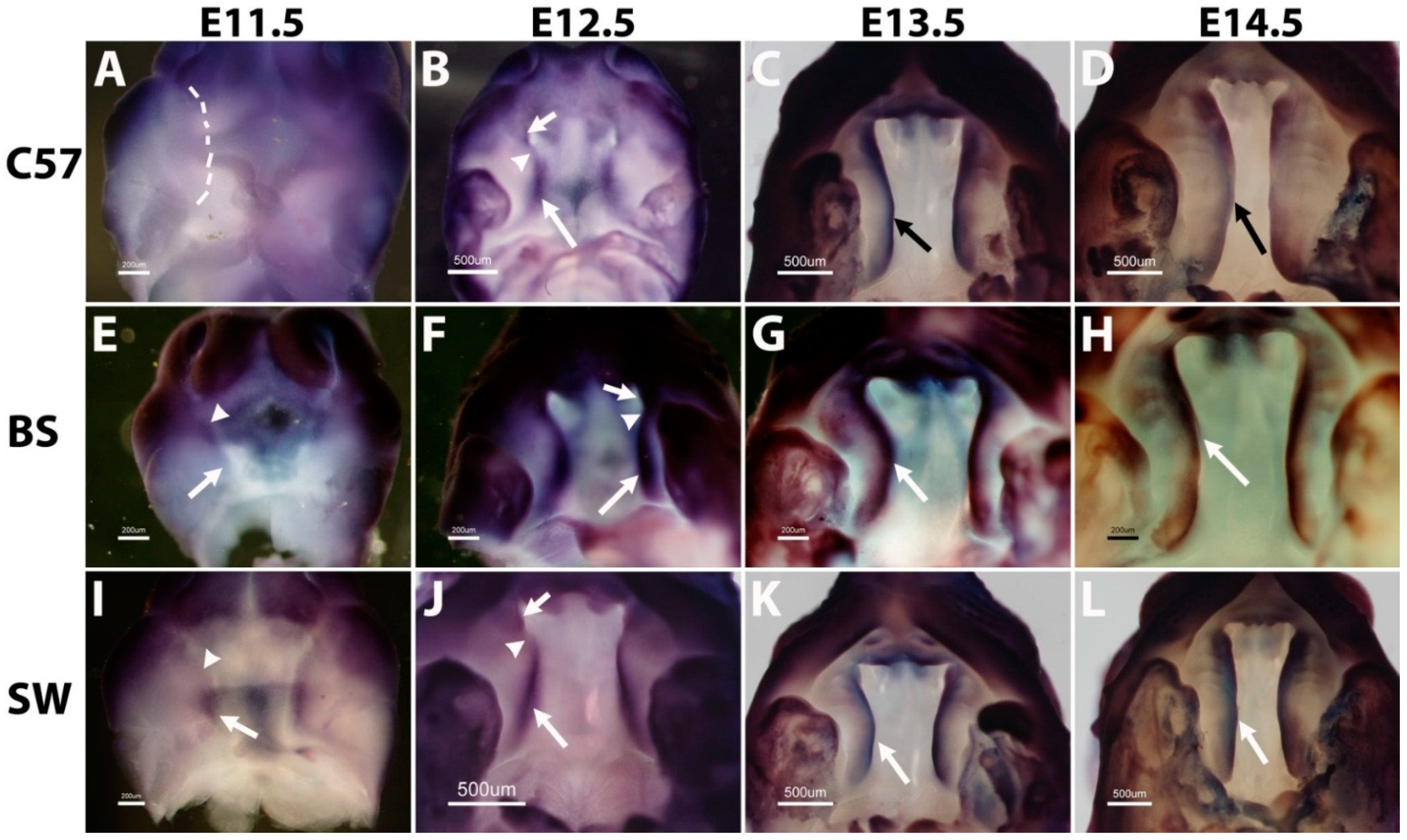
As shown in
Figure 2, on the C57B6 background,
Fgfr1 expression is undetectable in the palate outgrowth on E11.5 (
Figure 2A). On E12.5, high expression is found in the posterior region (arrow in
Figure 2B) and the very anterior tip (short arrow in
Figure 2B), but is absent in a portion of the anterior region (arrowhead in
Figure 2B). From E13.5 to E14.5, the expression covers the entire A-P axis with the same intensity (
Figure 2C,D).
Figure 2.
Fgfr1 gene displays divergent expression patterns during mouse secondary palate formation among C57B6 (A–D), Black Swiss (E–H) and Swiss Webster (I–L) strains. The dashed line in (A) indicates the unstained palate outgrowth. Scale bars represent 200 µm (A, E and I) and 500 µm (B–D, F–H and J–L); arrows and arrowheads indicate different regions in palate shelves, as illustrated in the Results and Discussion section. C57, BS and SW are the abbreviations for C57B6, Black Swiss and Swiss Webster.
Figure 2.
Fgfr1 gene displays divergent expression patterns during mouse secondary palate formation among C57B6 (A–D), Black Swiss (E–H) and Swiss Webster (I–L) strains. The dashed line in (A) indicates the unstained palate outgrowth. Scale bars represent 200 µm (A, E and I) and 500 µm (B–D, F–H and J–L); arrows and arrowheads indicate different regions in palate shelves, as illustrated in the Results and Discussion section. C57, BS and SW are the abbreviations for C57B6, Black Swiss and Swiss Webster.
Different from C57B6, the expression of
Fgfr1 in the Black Swiss strain is found in the palate outgrowth on E11.5 (
Figure 2E), and the expression is restricted to the anterior half (arrowhead in
Figure 2E). On E12.5, the expression covers the entire A-P axis of palate shelf with the same intensity in the anterior tip (short arrow in
Figure 2F), posterior region (arrow in
Figure 2F) and an anterior portion (arrowhead in
Figure 2F). This uniform A-P expression pattern extends to E13.5 and E14.5 (
Figure 2G,H).
In marked contrast to the Black Swiss strain, the expression of
Fgfr1 is only in the posterior region of the palate outgrowth on E11.5 in the Swiss Webster strain (arrow in
Figure 2I). On E12.5,
Fgfr1 is highly expressed in the posterior region (arrow in
Figure 2J), weakly expressed in the very anterior tip (short arrow in
Figure 2J), but is almost absent in a portion of the anterior region (arrowhead in
Figure 2J). From E13.5 to E14.5, similar to C57B6 and Black Swiss,
Fgfr1 expression on Swiss Webster is uniformly present along the entire palate A-P axis (
Figure 2K,L).
Therefore, the expression pattern of Fgfr1 in late palate development, E13.5 and E14.5, is similar among the three strains of C57B6, Black Swiss and Swiss Webster. However, the expression at early stages, especially the outgrowth stage on E11.5, shows considerable diversity in these three strains. Expression is undetectable on the C57B6 strain, in the anterior half only in the Black Swiss strain and the posterior half only in the Swiss Webster strain.
C57B6 and Black Swiss are two common inbred and outbred strains, respectively, used for gene targeting experiments. Swiss Webster is a common outbred strain widely used in gene expression studies. Despite the large number of studies regarding gene function on different strain backgrounds, very few studies explore the expression divergence of a given gene among different strains. It is likely that our findings with Meox-2 and Fgfr1 genes in secondary palate could also occur to other genes in other organs. Since the functions of Meox-2 and Fgfr1 in palate development have not been examined on different strains, the functional significance of this strain-dependent expression is not clear. However, the distinct expression of a given gene could, in principle, affect its function more or less and may account for the phenotype variations among different strains in gene targeting studies.


{kind=link}
{kind=link}

