
Stability Analysis and Hopf Bifurcation for the Brusselator Reaction–Diffusion System with Gene Expression Time Delay


Abstract
1. Introduction and Preliminaries
2. The Galerkin Method
3. The Dynamical Theoretical Formulation
4. Hopf Bifurcation
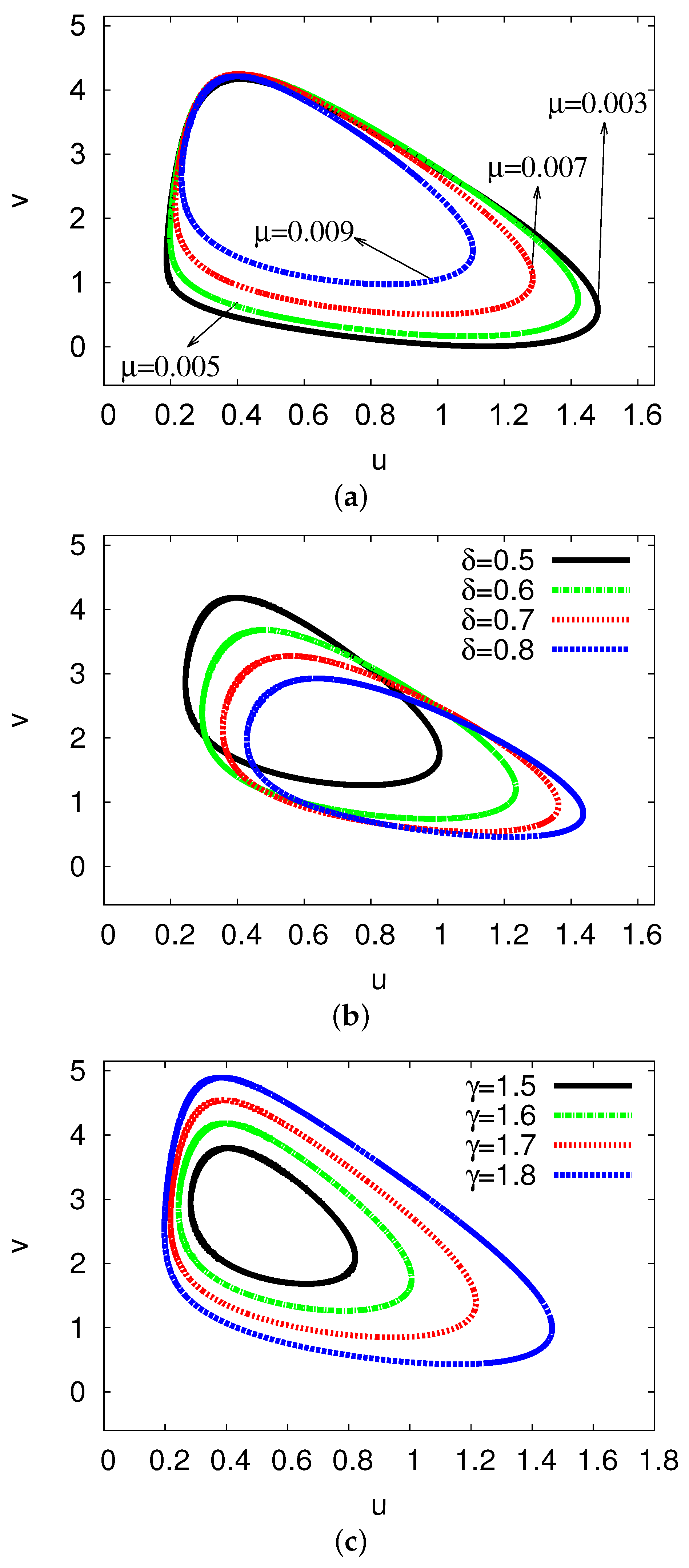
5. Bifurcation Diagrams
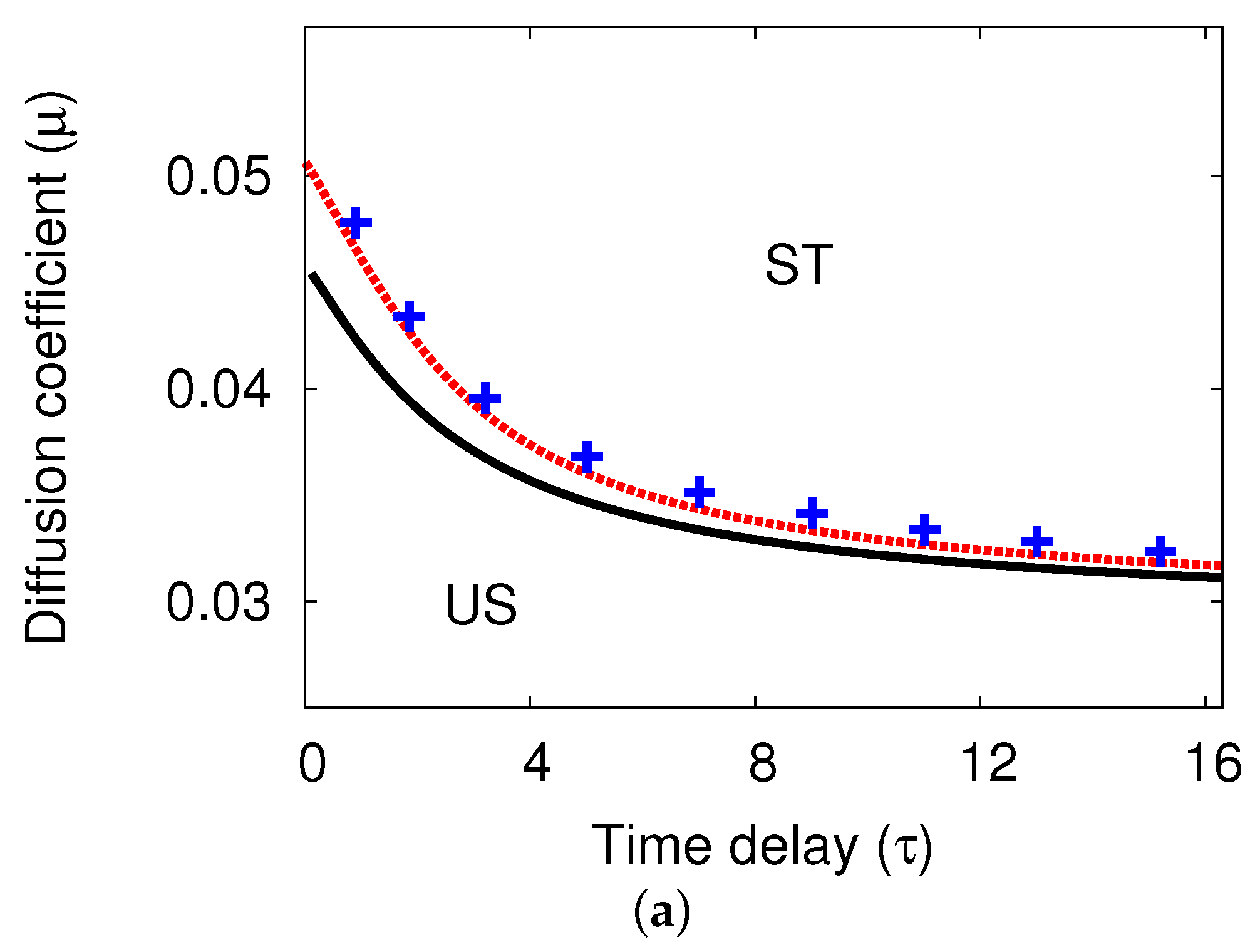
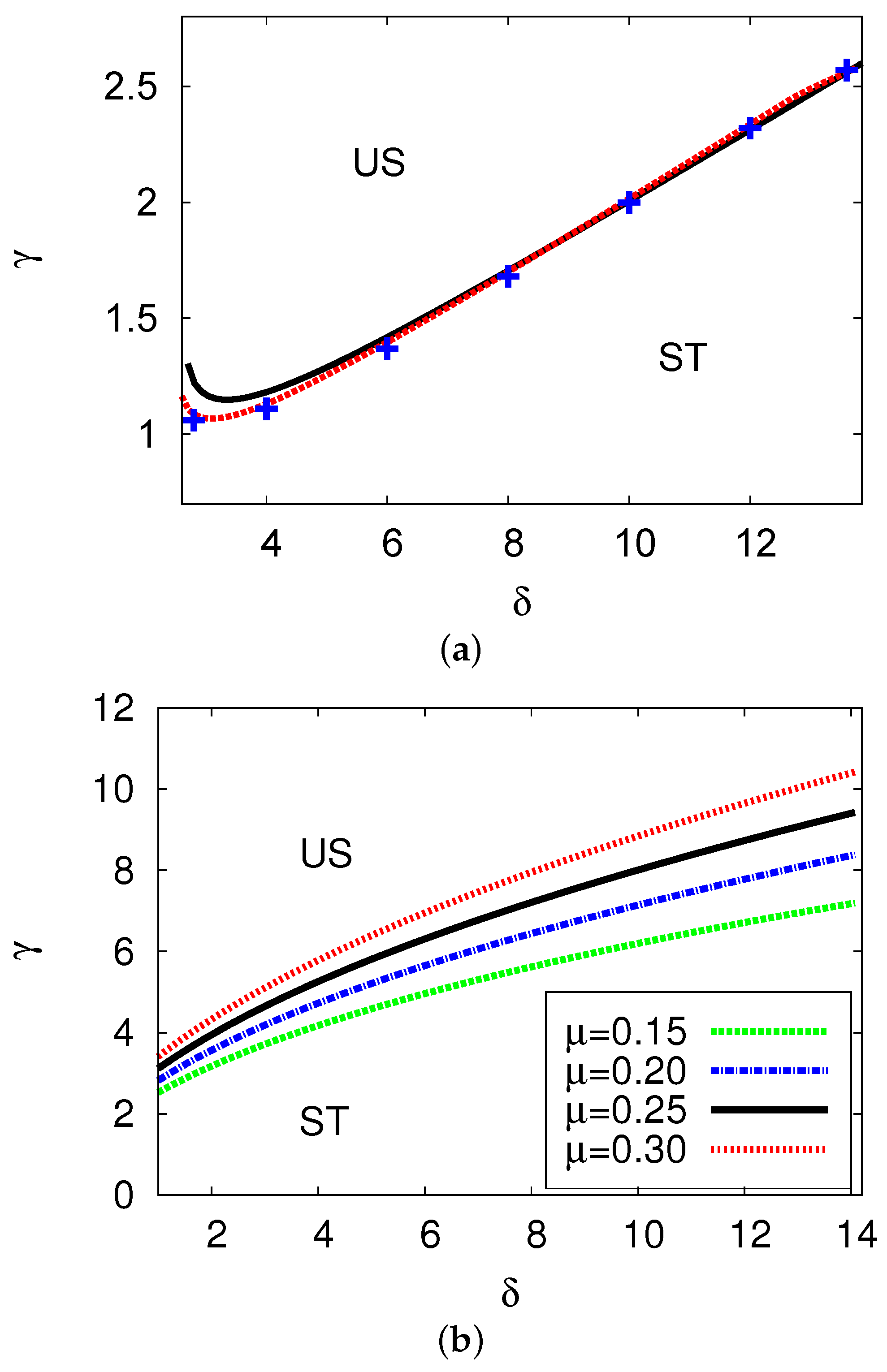
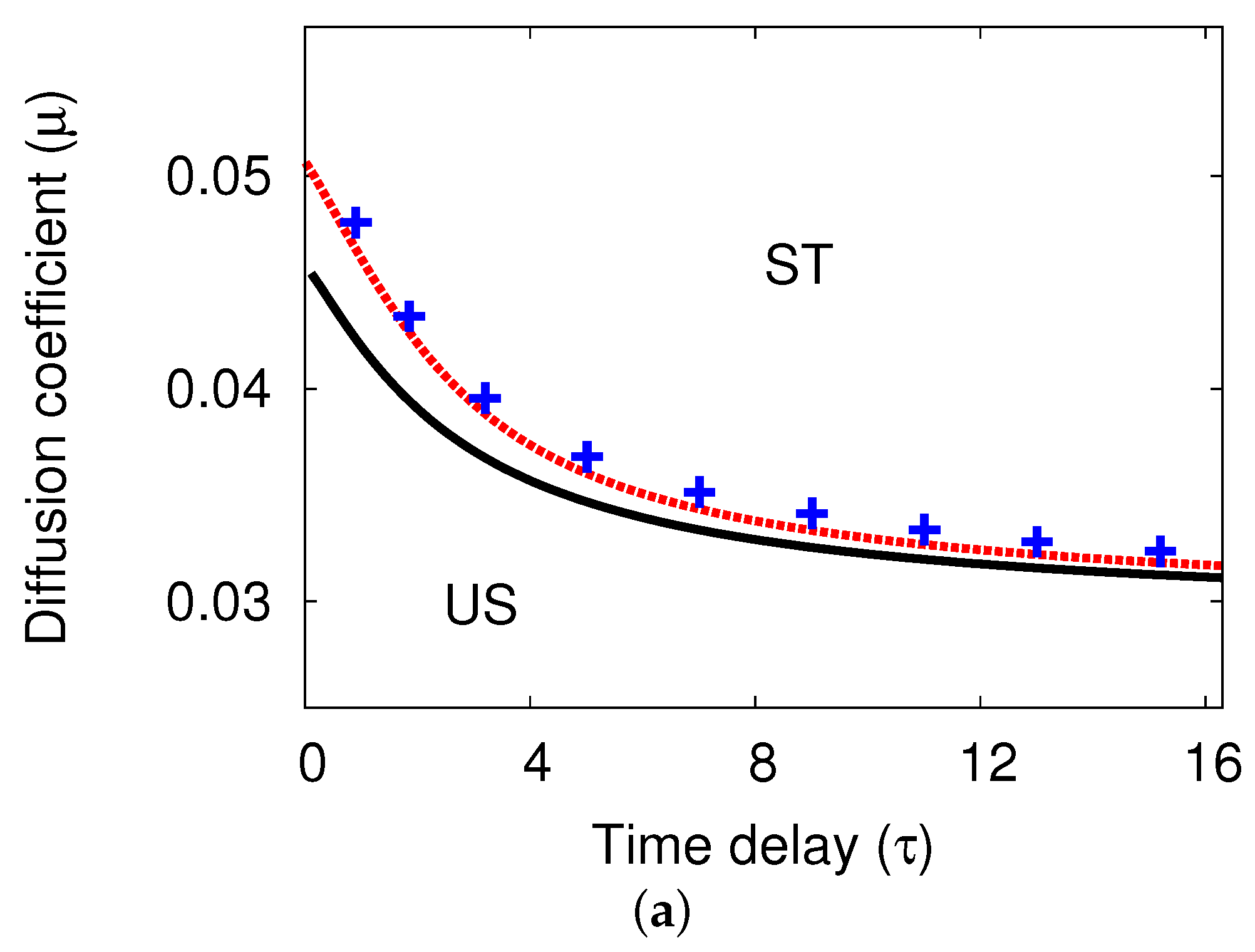
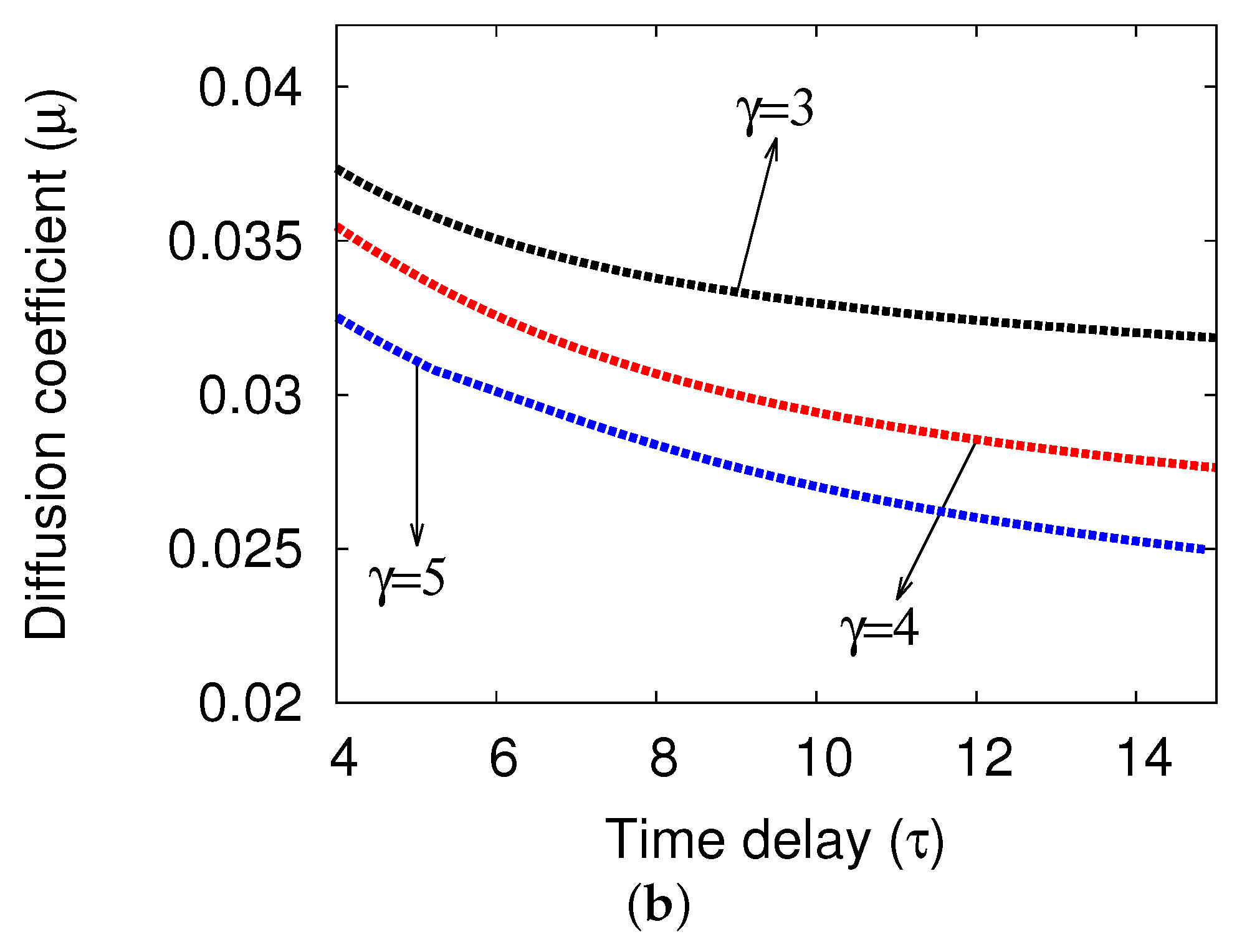
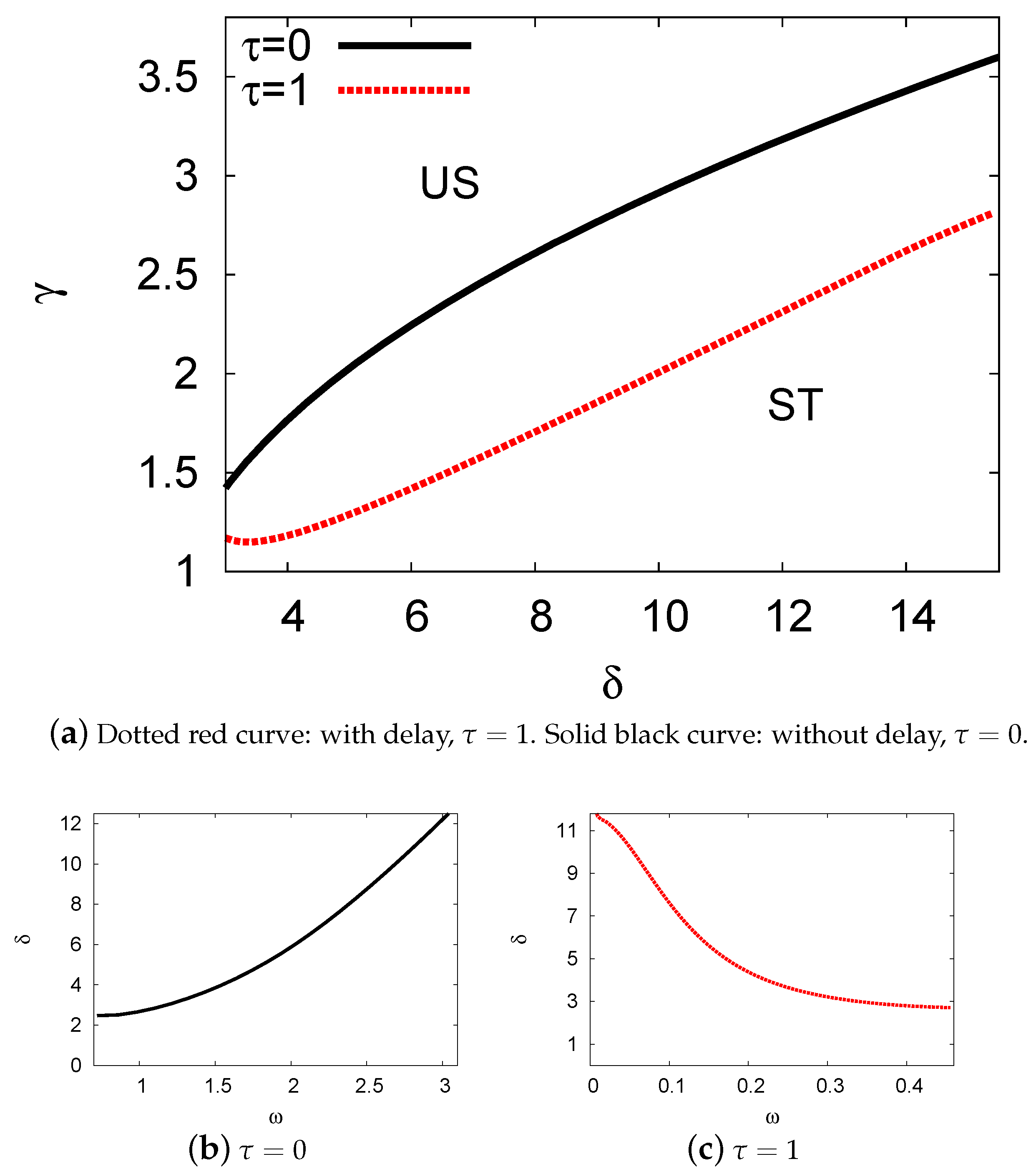
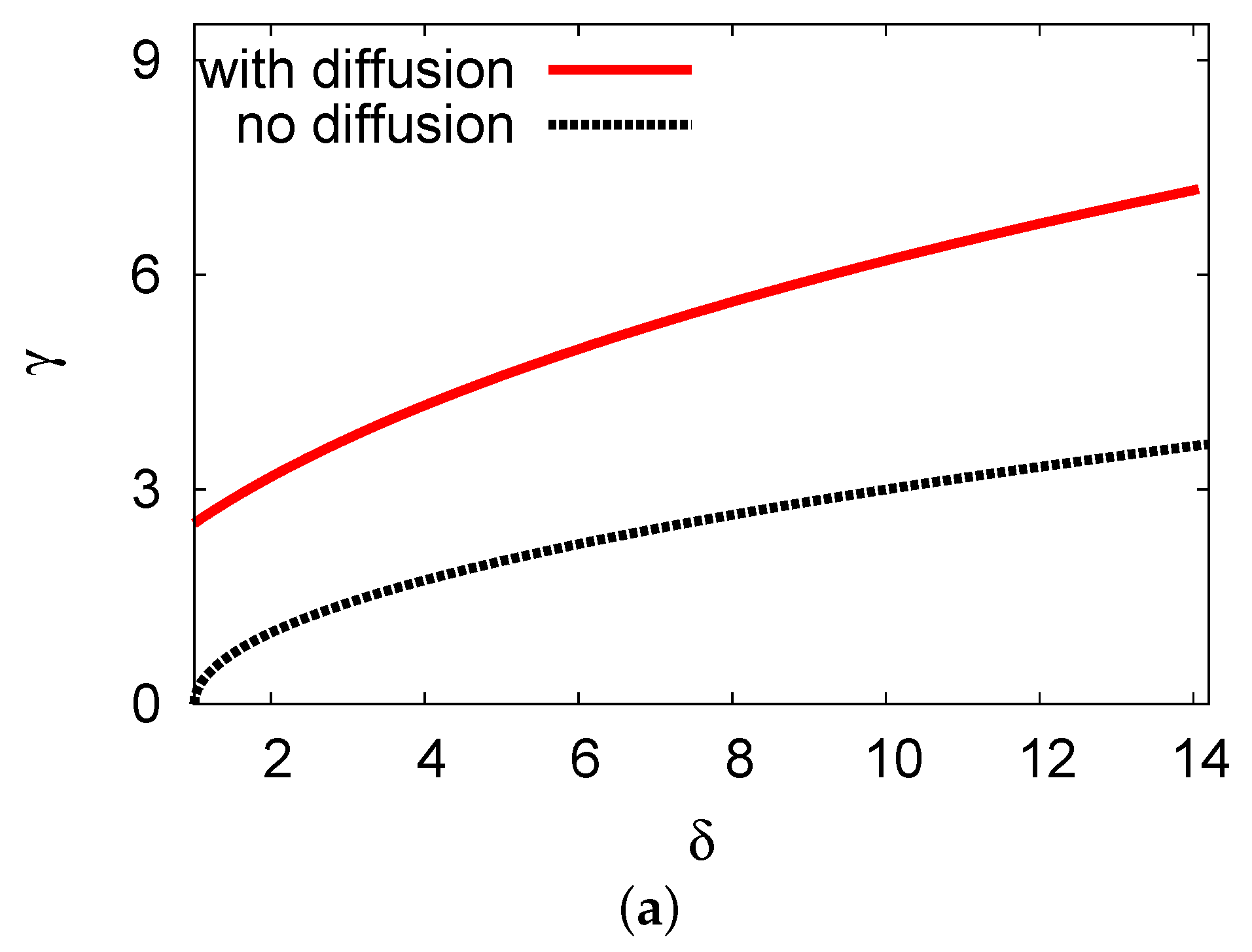
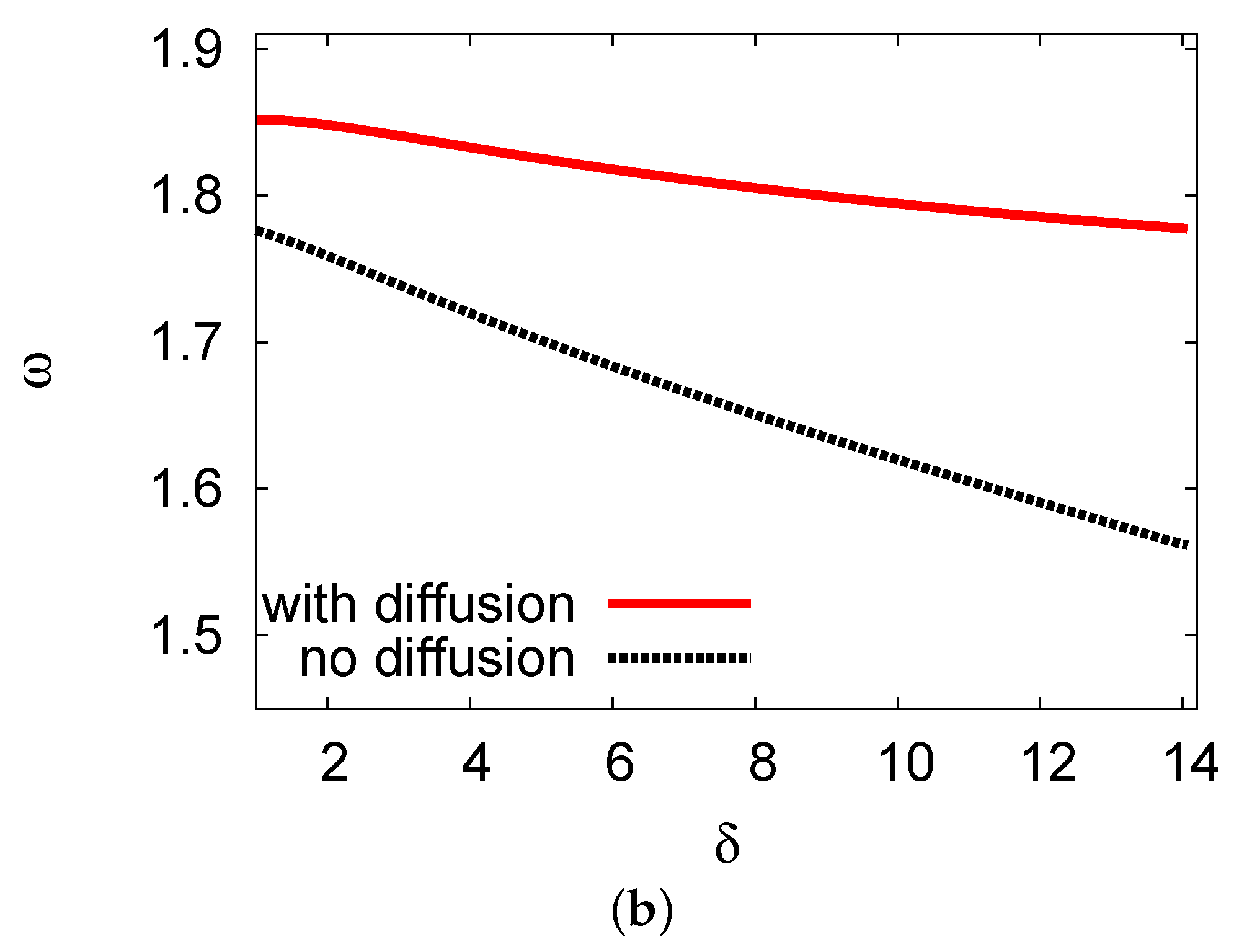
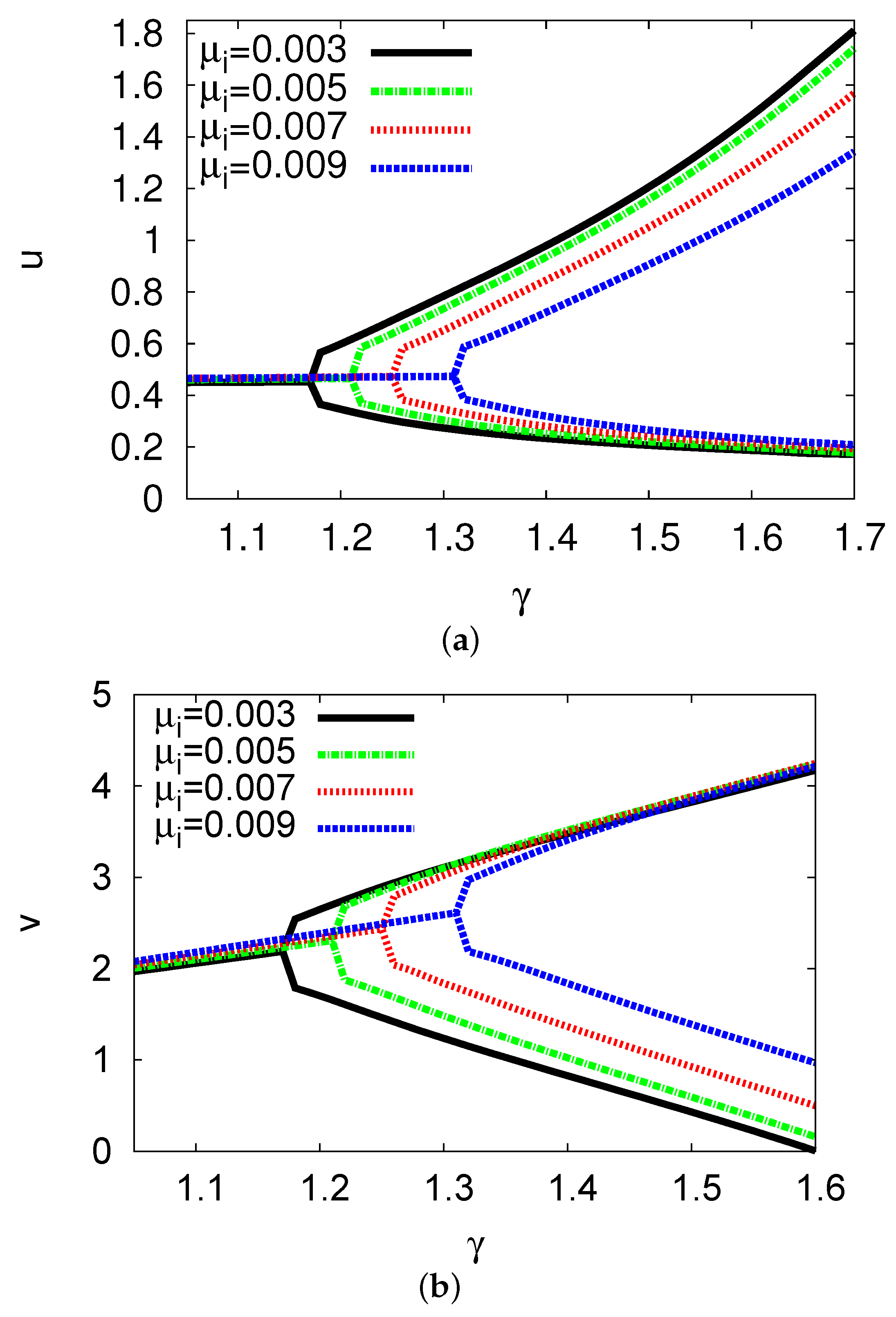
5.1. Bifurcation Diagrams at
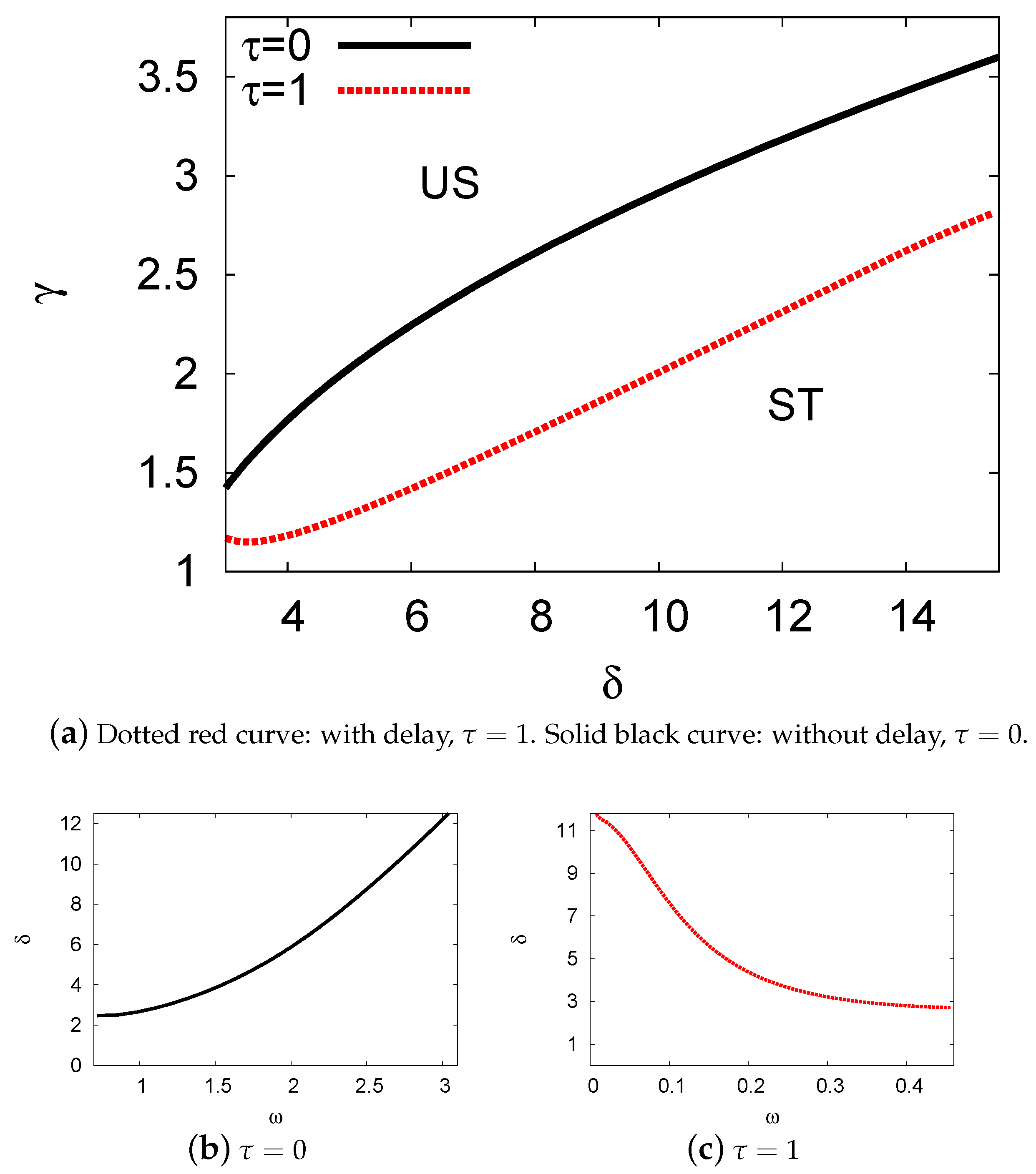
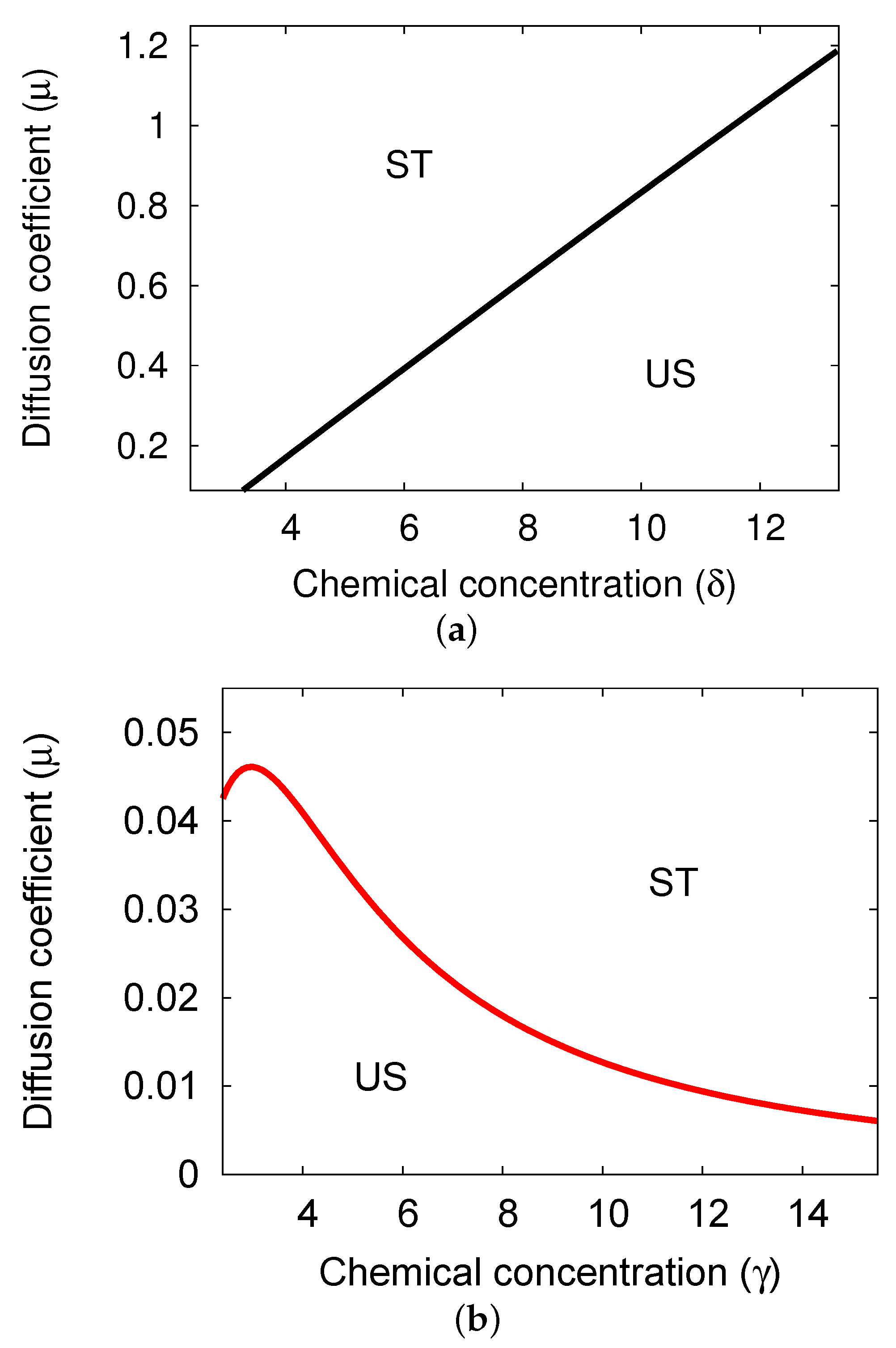
5.2. Bifurcation Diagrams at
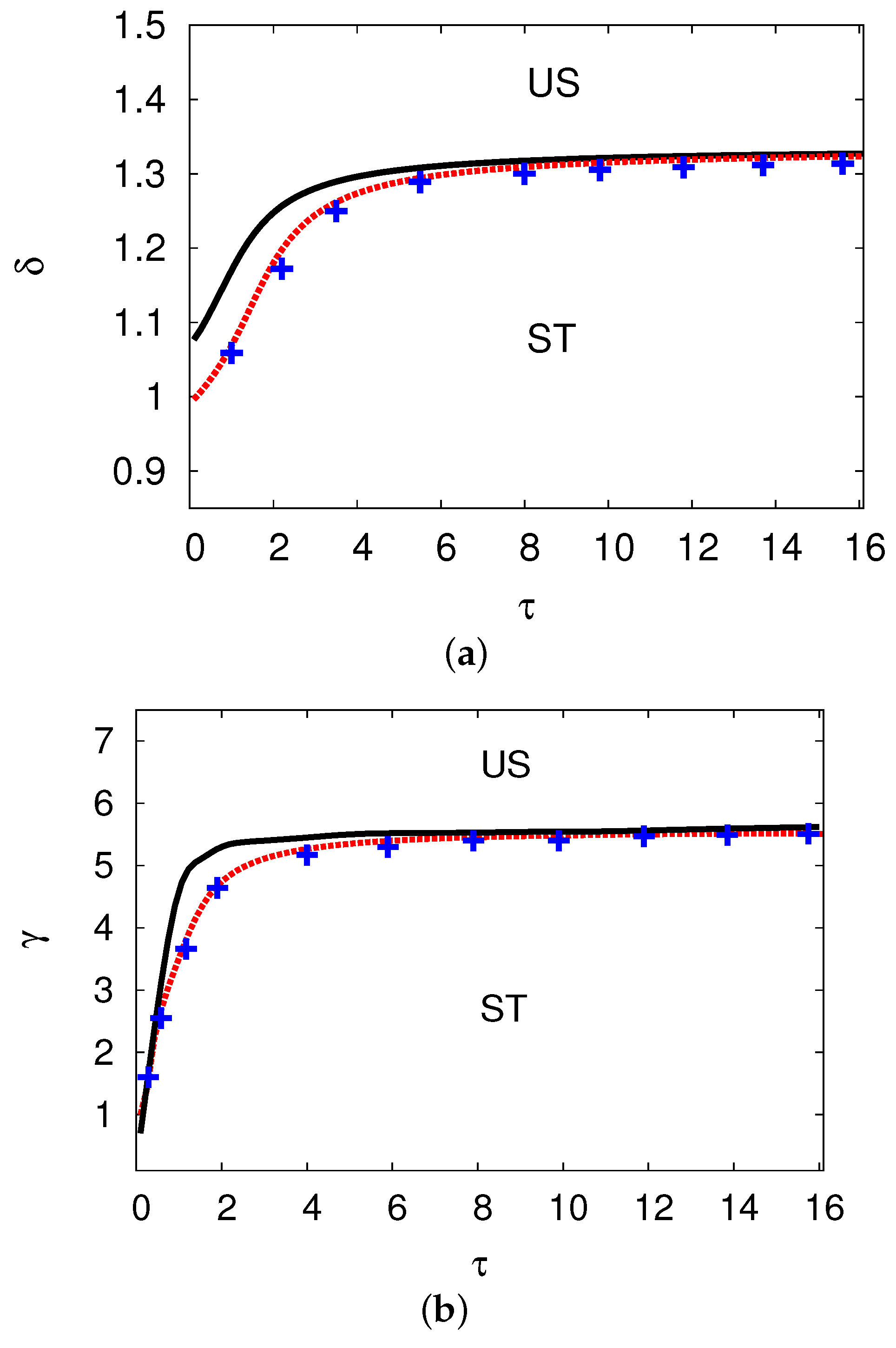
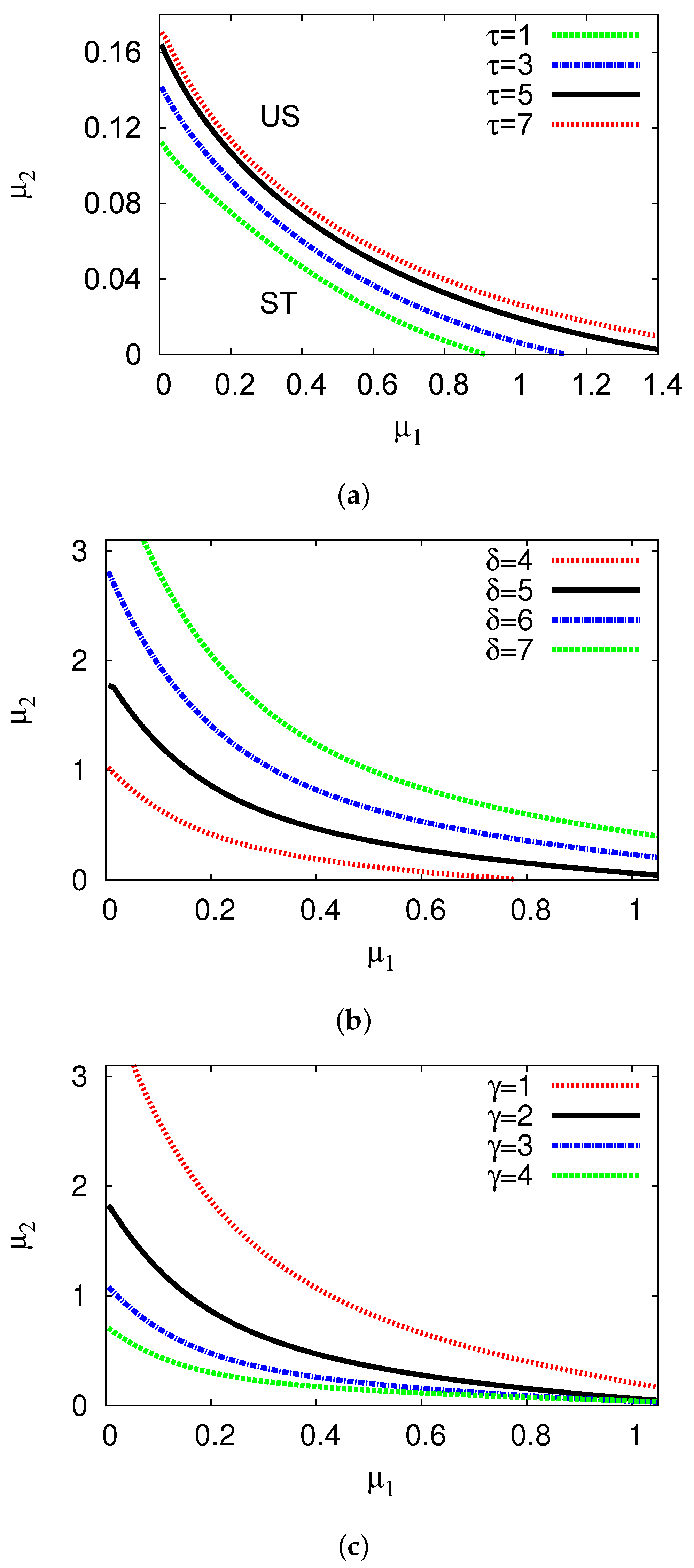
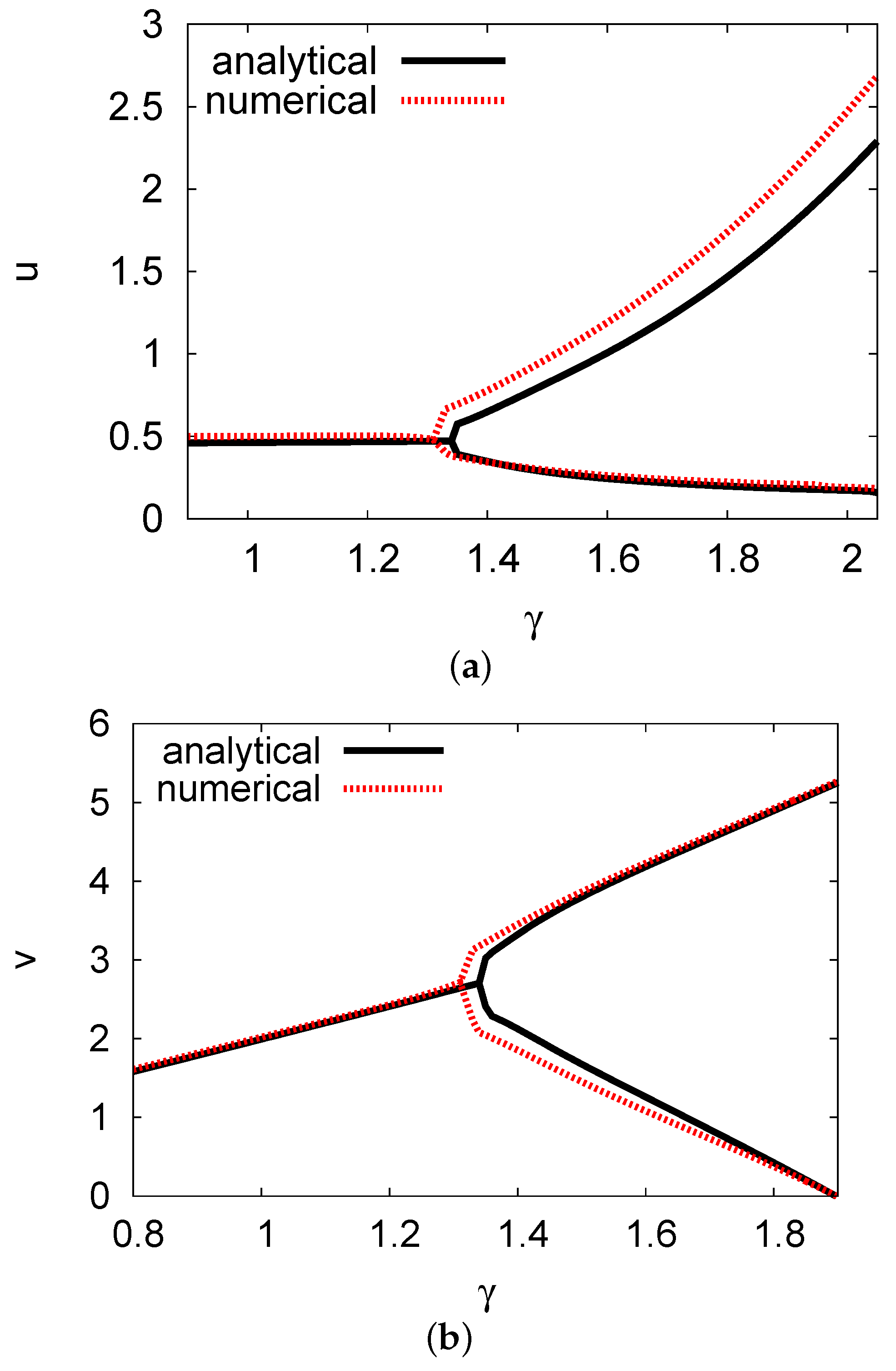
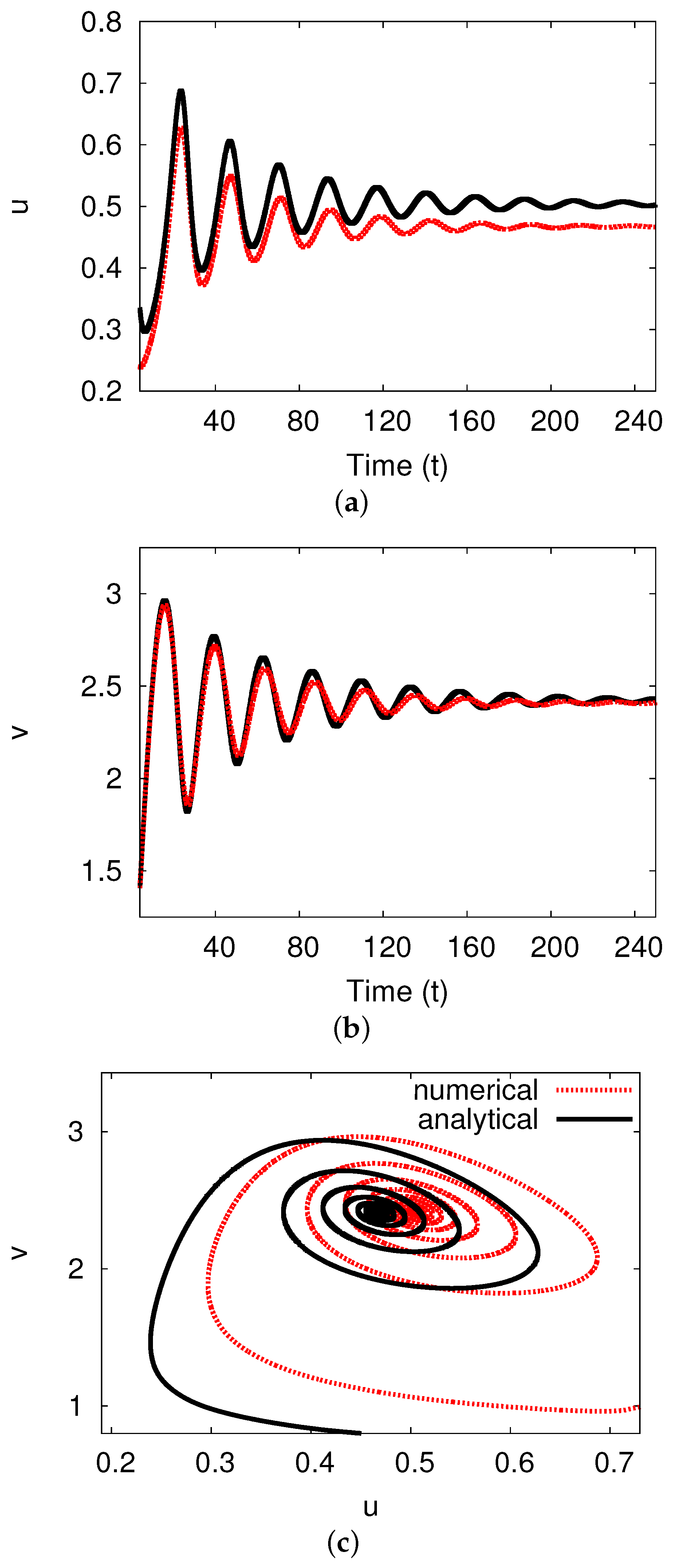
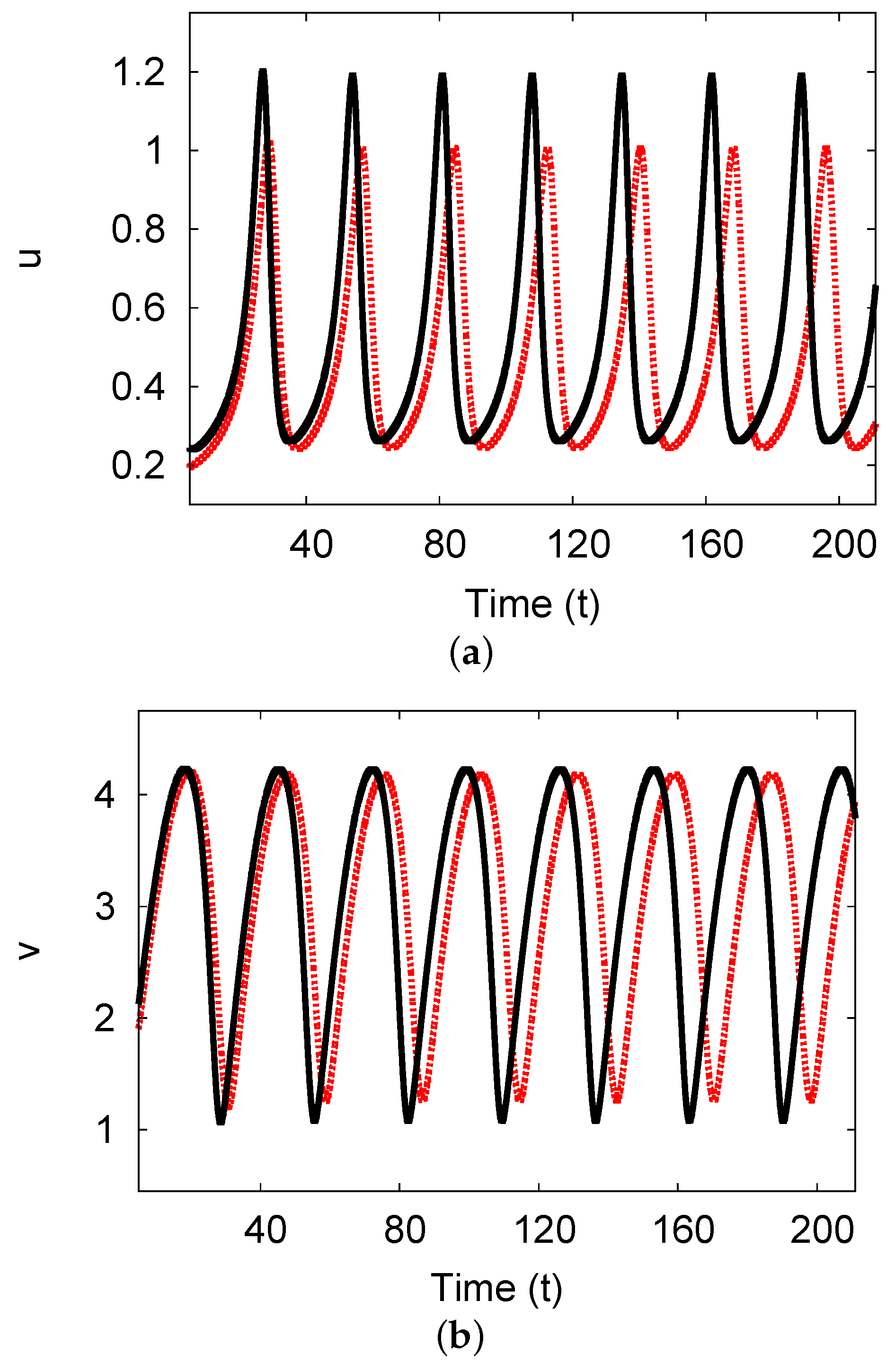
5.3. Examples and Numerical Simulations at Long Delay Time
6. Conclusions and Outlook
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CSTR | Continuous-flow stirred-tank reactor, |
| DDE | Delay differential equation, |
| DPDE | Delay partial differential equation, |
| BZ | Belousov–Zhabotinsky, |
| 4th | Fourth-order Runge–Kutta, |
| 2D | Two-dimensional. |
Appendix A. The Analytical System of Two-Term Scheme
References
- Alfifi, H.Y.; Marchant, T.R.; Nelson, M.I. Non-smooth feedback control for Belousov-Zhabotinskii reaction-diffusion equations: Semi-analytical solutions. J. Math Chem. 2016, 57, 157–178. [Google Scholar] [CrossRef]
- Ren, J.; Gao, J.; Yang, W. Computational simulation of Belousov-Zhabotinskii oscillating chemical reaction. Comput. Visual Sci. 2009, 12, 227–234. [Google Scholar] [CrossRef]
- Adomian, G. The diffusion-Brusselator equation. Comput. Math. Appl. 1995, 29, 1–3. [Google Scholar] [CrossRef]
- Alfifi, H.Y. Semi-analytical solutions for the Brusselator reaction-diffusion model. ANZIM J. 2017, 59, 167–182. [Google Scholar] [CrossRef]
- Alfifi, H.Y. Feedback Control for a Diffusive and Delayed Brusselator Model: Semi-Analytical Solutions. Symmetry 2021, 13, 725. [Google Scholar] [CrossRef]
- Guo, G.; Wu, J.; Ren, X. Hopf bifurcation in general Brusselator system with diffusion. Appl. Math. Mech. Engl. Ed. 2011, 32, 1177–1186. [Google Scholar] [CrossRef]
- Marchant, T.R.; Nelson, M.I. Semi-analytical solution for one-and two-dimensional pellet problems. Proc. R. Soc. Lond. 2004, A460, 2381–2394. [Google Scholar] [CrossRef]
- Al Noufaey, K. A semi-analytical approach for the reversible Schnakenberg reaction diffusion system. J. Results Phys. 2020, 16, 102858. [Google Scholar] [CrossRef]
- Al Noufaey, K.S.; Marchant, T.R. Semi-analytical solutions for the reversible Selkov model with feedback delay. Appl. Math. Comput. 2014, 232, 49–59. [Google Scholar] [CrossRef]
- Alharthi, M.R.; Marchant, T.R.; Nelson, M.I. Mixed quadratic-cubic autocatalytic reaction-diffusion equations: Semi-analytical solutions. Appl. Math. Model. 2014, 38, 5160–5173. [Google Scholar] [CrossRef]
- Marchant, T.R. Cubic autocatalytic reaction diffusion equations: Semi-analytical solutions. Proc. R. Soc. Lond. 2002, A458, 873–888. [Google Scholar] [CrossRef]
- Alfifi, H. Stability analysis for Schnakenberg reaction-diffusion model with gene expression time delay. Chaos Solitons Fractals 2022, 155, 111730. [Google Scholar] [CrossRef]
- Gray, P.; Griffiths, J.F.; Scott, K. Branched-chain reactions in open systems: Theory of the oscillatory ignition limit for the hydrogen+ oxygen reaction in a continuous-flow stirred-tank reactor. Proc. R. Soc. Lond. 1984, A394, 243–258. [Google Scholar]
- Gu, Y.; Ullah, S.; Khan, M.A.; Alshahrani, M.Y.; Abohassan, M.; Riaz, M.B. Mathematical modeling and stability analysis of the COVID-19 with quarantine and isolation. Results Phys. 2022, 34, 105284. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, Q.; Tian, C. Effect of Time Delay on Spatial Patterns in a Airal Infection Model with Diffusion. Math. Model. Anal. 2016, 21, 143–158. [Google Scholar] [CrossRef]
- Prigogine, I.; Lefever, R. Symmetry Breaking Instabilities in Dissipative Systems II. J. Chem. Phys. 1968, 48, 1665–1700. [Google Scholar] [CrossRef]
- Ang, W. The two-dimensional reaction-diffusion Brusselator system: A dual-reciprocity boundary element solution. Eng. Anal. Bound. Elem. 2003, 27, 897–903. [Google Scholar] [CrossRef]
- Kumar, S.; Jiwari, D.R.; Mittal, R.C. Numerical simulation for computational modelling of reaction-diffusion Brusselatormodel arising in chemical processes. J. Math. Chem. 2019, 57, 149–179. [Google Scholar] [CrossRef]
- Mittal, R.; Jiwari, R. Numerical study of two-dimensional reaction-diffusion Brusselator system. Appl. Math. Comput. 2011, 217, 5404–5415. [Google Scholar]
- Kumar, S.; Khan, Y.; Yildirim, A. A mathematical modeling arising in the chemical systems and its approximate numerical solution. Asia Pac. J. Chem. Eng. 2012, 7, 835–840. [Google Scholar] [CrossRef]
- Sarwar, S.; Iqbal, S. Stability analysis, dynamical behavior and analytical solutions of nonlinear fractional differential system arising in chemical reaction. Chin. J. Phys. 2017, 56, 374–384. [Google Scholar] [CrossRef]
- Arafa, A.; Rida, S.; Mohamed, H. Approximate analytical solutions of Schnakenberg systems by homotopy analysis method. Appl. Math. Model. 2012, 36, 4789–4796. [Google Scholar] [CrossRef]
- Michael, J.; Wei, J. The existence and stability of asymmetric spike patterns for the Schnakenberg model. Stud. Appl. Math. 2002, 109, 229C264. [Google Scholar]
- Schnakenberg, J. Simple chemical reaction systems with limit cycle behavior. J. Theor. Biol. 1979, 81, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Tyson, J.J. Some further studies of nonlinear oscillations in chemical systems. JCP 1973, 58, 3919–3930. [Google Scholar] [CrossRef]
- Ghergu, M.; Radulescu, V. Nonlinear PDEs: Mathematical Models in Biology, Chemistry and Population Genetics; Springer: Berlin/Heidelberg, Germany, 2012; Volume 160. [Google Scholar] [CrossRef]
- Yan, X.P.; Zhang, P.; Zhang, C.H. Turing instability and spatially homogeneous Hopf bifurcation in a diffusive Brusselator system. Nonlinear Anal. Model. Control 2020, 25, 638–657. [Google Scholar] [CrossRef]
- Twizell, E.; Gumel, A.; Cao, Q. A second-order scheme for the Brusselator reaction-diffusion system. J. Math. Chem. 1999, 26, 297–316. [Google Scholar] [CrossRef]
- Lv, Y.; Liu, Z. Turing-Hopf bifurcation analysis and normal form of a diffusive Brusselator model with gene expression time delay. Chaos Solitons Fractals 2021, 152, 111478. [Google Scholar] [CrossRef]
- Alfifi, H. Stability analysis and Hopf bifurcation for two-species reaction-diffusion-advection competition systems with two time delays. Appl. Math. Comput. 2024, 474, 128684. [Google Scholar] [CrossRef]
- Temimi, H.; Ben-Romdhane, M.; El-Borgi, S.; Cha, Y.J. Time-Delay Effects on Controlled Seismically Excited Linear and Nonlinear Structures. Int. J. Struct. Stab. Dyn. 2016, 16, 1550031. [Google Scholar] [CrossRef]
- Wazwaz, A. The decomposition method applied to systems of partial differential equations and to the reaction-diffusion Brusselator model. Appl. Math. Comput. 2000, 110, 251–264. [Google Scholar] [CrossRef]
- Fletcher, C.A. Computational Galerkin Methods; Springer: New York, NY, USA, 1984. [Google Scholar]
- Al Noufaey, K.S.; Marchant, T.R.; Edwards, M.P. The diffusive Lotka-Volterra predator-prey system with delay. Math. Biosci. 2015, 270, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Alfifi, H. Effects of diffusion and delayed immune response on dynamic behavior in a viral model. Appl. Math. Comput. 2023, 441, 127714. [Google Scholar] [CrossRef]
- Alfifi, H.Y.; Marchant, T.R.; Nelson, M.I. Semi-analytical solutions for the 1- and 2-D diffusive Nicholson’s blowflies equation. IMA J. Appl. Math. 2014, 79, 175–199. [Google Scholar] [CrossRef]
- Belousov, B.P. An Oscillating Reaction and Its Mechanism. In Collection of Abstracts on Radiation Medicine [Sborn. Referat. Radiat. Med.]; Medgiz: Moscow, Russia, 1959; p. 145. [Google Scholar]
- Erneux, T. Applied Delay Differential Equations; Springer: New York, NY, USA, 2009. [Google Scholar]
- Hale, J. Theory of Functional Differential Equations; Springer: New York, NY, USA, 1977. [Google Scholar]
- Looss, G.; Joseph, D.D. Elementary Stability and Bifurcation Theory, 2nd ed.; Springer: New York, NY, USA, 1990. [Google Scholar]
- Nowak, M.; May, R. Virus Dynamics; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Smith, G.D. Numerical Solution of Partial Differential Equations: Finite Difference Methods, 3rd ed.; Oxford: New York, NY, USA, 1985. [Google Scholar]
- Verwer, J.; Hundsdorfer, W.; Sommeijer, B. Convergence properties of the Runge-Kutta-Chebyshev method. Numer. Math. 1990, 57, 157–178. [Google Scholar] [CrossRef]
- Maplesoft, a Division of Waterloo Maple Inc. Maple; Maplesoft: Waterloo, ON, Canada, 2019. [Google Scholar]
- Al Noufaey, K.S. Stability Analysis of a Diffusive Three-Species Ecological System with Time Delays. Symmetry 2021, 13, 2217. [Google Scholar] [CrossRef]
- Canabarro, A.; Gléria, I.; Lyra, M. Periodic solutions and chaos in a non-linear model for the delayed cellular immune response. Phys. A Stat. Mech. Its Appl. 2004, 342, 234–241. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfifi, H.Y.; Almuaddi, S.M. Stability Analysis and Hopf Bifurcation for the Brusselator Reaction–Diffusion System with Gene Expression Time Delay. Mathematics 2024, 12, 1170. https://doi.org/10.3390/math12081170
Alfifi HY, Almuaddi SM. Stability Analysis and Hopf Bifurcation for the Brusselator Reaction–Diffusion System with Gene Expression Time Delay. Mathematics. 2024; 12(8):1170. https://doi.org/10.3390/math12081170
Chicago/Turabian StyleAlfifi, Hassan Y., and Saad M. Almuaddi. 2024. "Stability Analysis and Hopf Bifurcation for the Brusselator Reaction–Diffusion System with Gene Expression Time Delay" Mathematics 12, no. 8: 1170. https://doi.org/10.3390/math12081170
APA StyleAlfifi, H. Y., & Almuaddi, S. M. (2024). Stability Analysis and Hopf Bifurcation for the Brusselator Reaction–Diffusion System with Gene Expression Time Delay. Mathematics, 12(8), 1170. https://doi.org/10.3390/math12081170