
Alterations in the Proteome and Phosphoproteome Profiles of Rat Hippocampus after Six Months of Morphine Withdrawal: Comparison with the Forebrain Cortex


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Morphine Administration and Drug Withdrawal of Male Wistar Rats
2.3. Preparation of Samples
2.4. Detection of Phosphoproteins by Pro-Q® Diamond Staining
2.5. Scanning of Phosphorylated Proteins
2.6. Staining by Colloidal Coomassie Blue (CBB)
2.7. Statistical Analysis
2.8. nLC-MS/MS
3. Results
3.1. Proteomic Analysis of the Rat Hippocampus Isolated from Animals after 6 Months of Morphine Withdrawal
3.1.1. Pro-Q® Diamond Staining and Colloidal Coomassie Blue Staining of 2D Gels
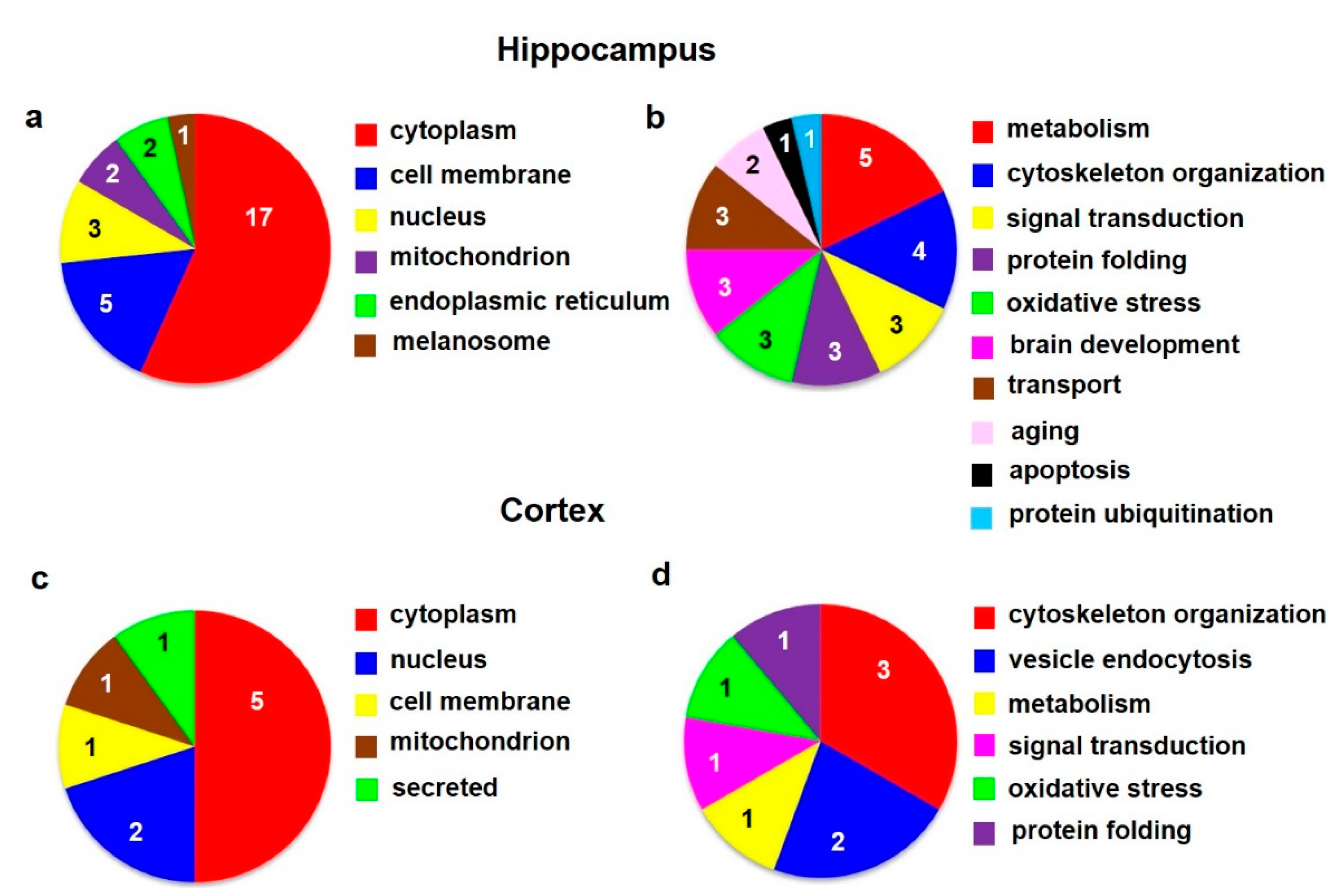
3.1.2. GO Enrichment Analysis of Altered Hippocampal Proteins
3.2. Proteomic Analysis of the Rat Forebrain Cortex Isolated from Animals after 6 Months of Morphine Withdrawal
3.2.1. Pro-Q® Diamond Staining and Colloidal Coomassie Blue Staining of 2D Gels
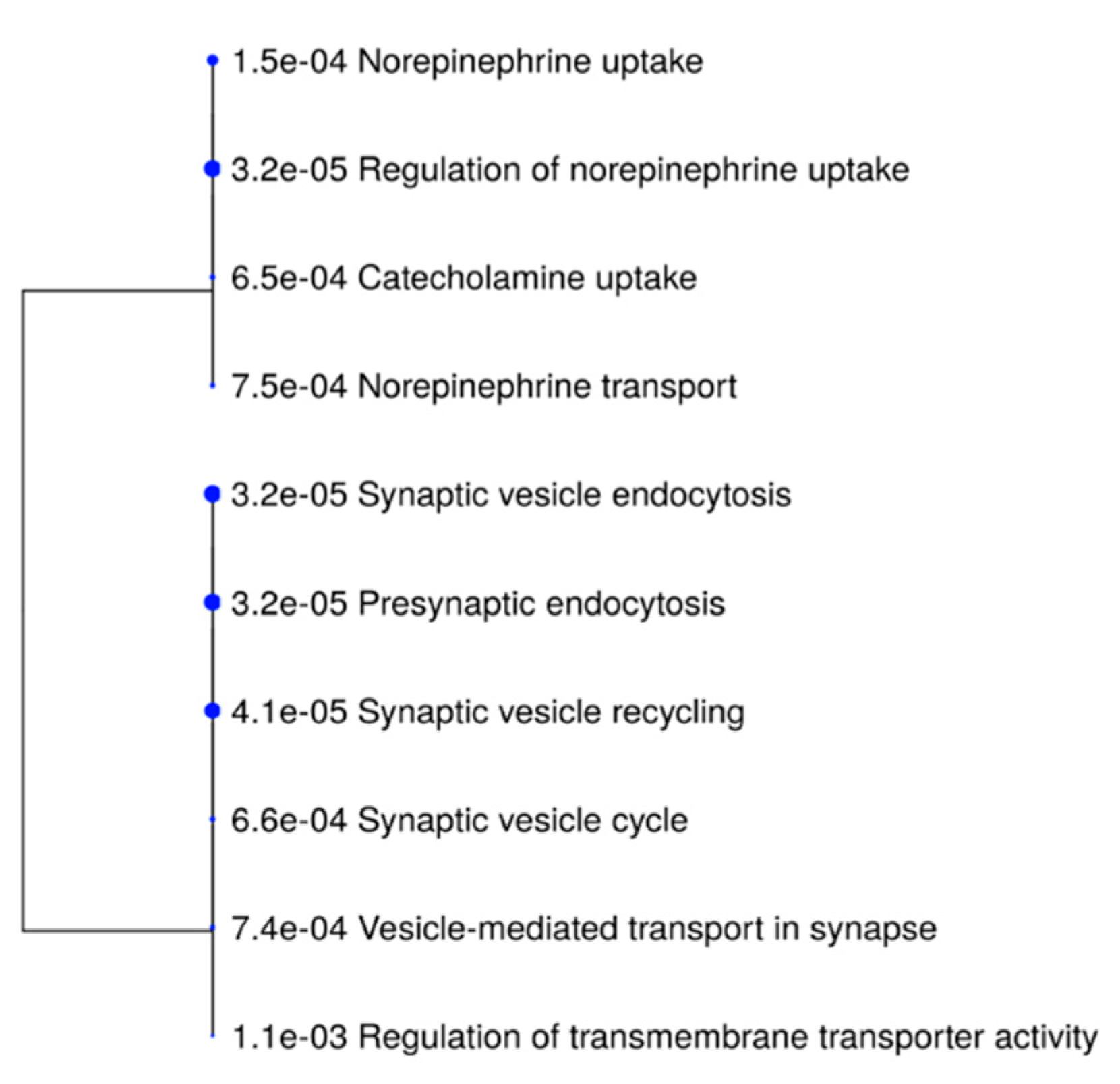
3.2.2. GO Enrichment Analysis of Altered Cortical Proteins
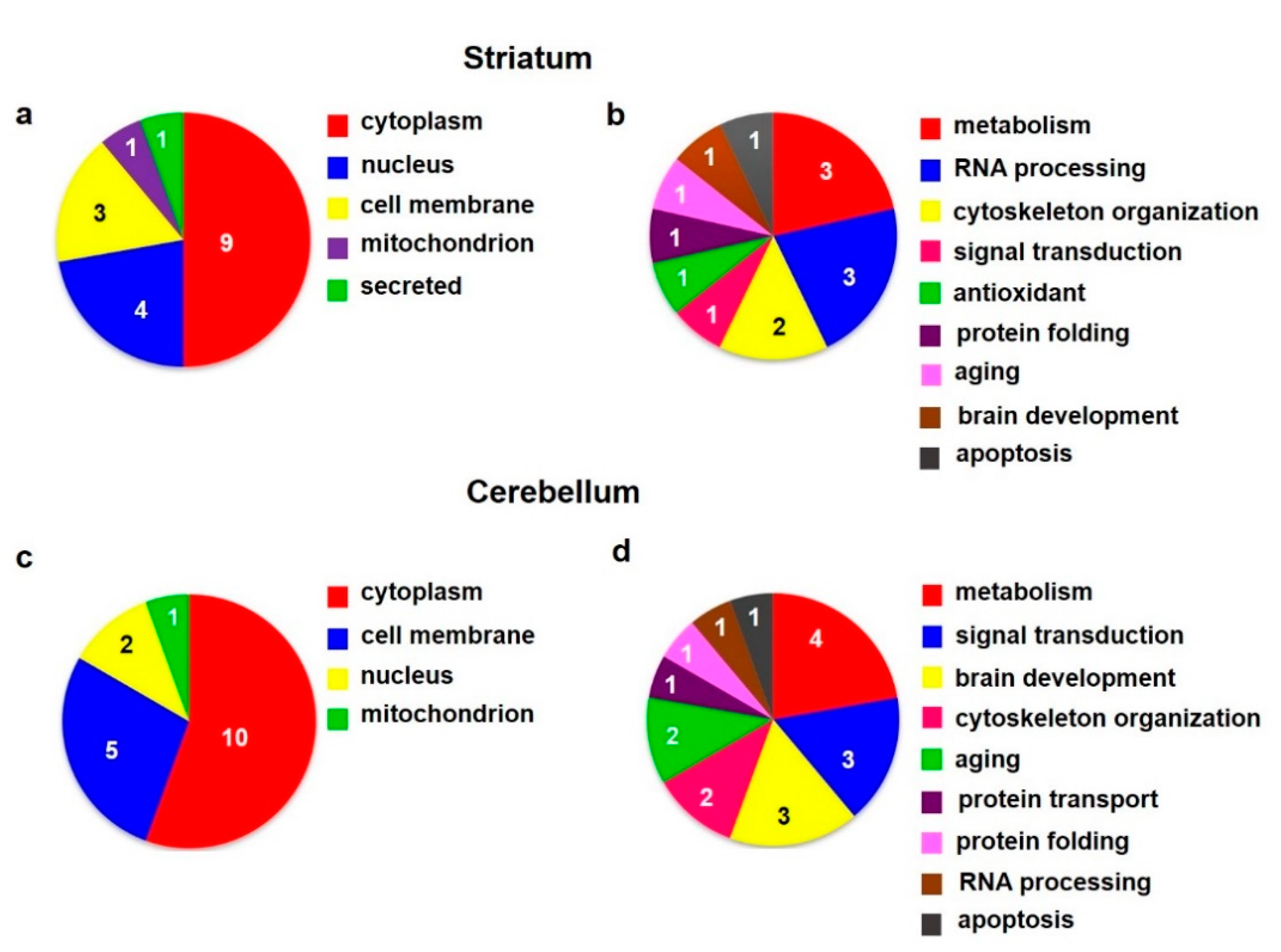
3.3. Proteomic Analysis of the Rat Striatum and Cerebellum Isolated from Animals after 6 Months of Morphine Withdrawal
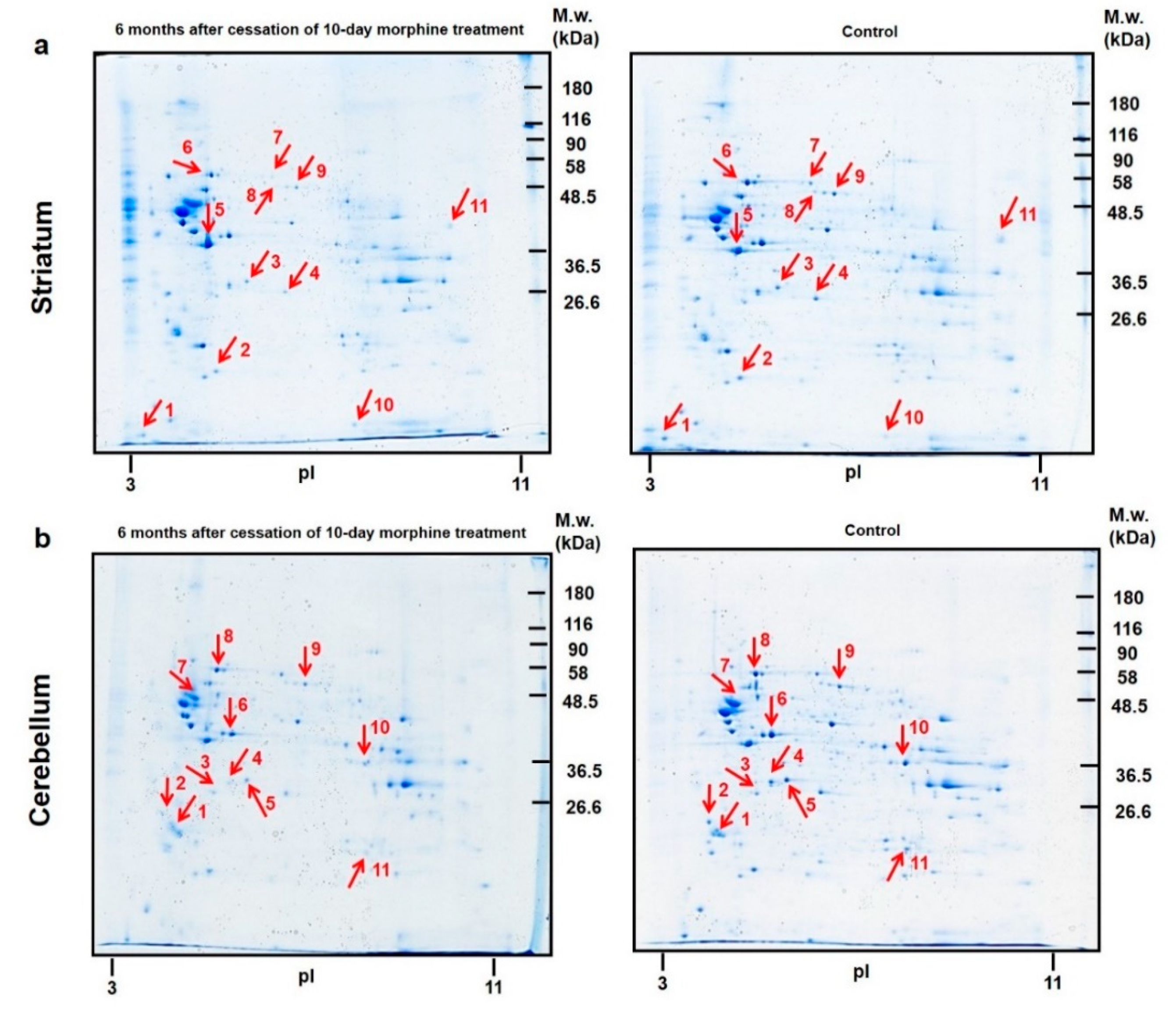
3.3.1. Colloidal Coomassie Blue Protein Staining of Striatal 2D Gels
3.3.2. Colloidal Coomassie Blue Protein Staining of Cerebellar 2D Gels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Azevedo Neto, J.; Constanzini, A.; De Giorgio, R.; Lambert, D.G.; Ruzza, C.; Calò, G. Biased versus partial agonism in the search for safer opioid analgesics. Molecules 2020, 25, 3870. [Google Scholar] [CrossRef]
- Paul, A.K.; Gueven, N.; Dietis, N. Profiling the effects of repetitive morphine administration on motor behavior in rats. Molecules 2021, 26, 4355. [Google Scholar] [CrossRef]
- Chen, Y.; Mestek, A.; Liu, J.; Hurley, J.A.; Yu, L. Molecular cloning and functional expression of a mu-opioid receptor from rat brain. Mol. Pharmacol. 1993, 44, 8–12. [Google Scholar] [CrossRef]
- Pacifici, G.M. Metabolism and pharmacokinetics of morphine in neonates: A review. Clinics 2016, 71, 474–480. [Google Scholar] [CrossRef]
- Bourova, L.; Vosahlikova, M.; Kagan, D.; Dlouha, K.; Novotny, J.; Svoboda, P. Long-term adaptation to high doses of morphine causes desensitization of µ-OR- and δ-OR-stimulated G-protein response in forebrain cortex but does not decrease the amount of G-protein alpha subunit. Med. Sci. Monit. 2010, 16, 260–270. [Google Scholar]
- Sim, L.J.; Selley, D.E.; Dworkin, S.I.; Childers, S.R. Effects of chronic morphine administration on mu opioid receptor-stimulated [35S] GTPgammaS autoradiography in rat brain. J. Neurosci. 1996, 16, 2684–2692. [Google Scholar] [CrossRef]
- Selley, D.E.; Liu, Q.; Childers, S.R. Signal transduction correlates of µ opioid agonist intrinsic efficacy: Receptor-stimulated [35S] GTPγS binding in mMOR-CHO cells and rat thalamus. J. Pharm. Exp. Ther. 1998, 285, 496–505. [Google Scholar]
- Selley, D.E.; Cao, Q.L.; Liu, Q.; Childers, S.R. Effect of sodium on agonist efficacy for G-protein activation in µ-opioid receptor-transfected CHO cells and rat thalamus. Br. J. Pharmacol. 2000, 130, 987–996. [Google Scholar] [CrossRef]
- Sim-Selley, L.J.; Selley, D.E.; Vogt, L.J.; Childers, S.R.; Martin, T.J. Chronic heroin self-administration desensitizes mu opioid receptor-activated G-proteins in specific regions of rat brain. J. Neurosci. 2000, 20, 4555–4562. [Google Scholar] [CrossRef] [PubMed]
- Maher, C.E.; Selley, D.E.; Childers, S.R. Relationship of mu opioid receptor binding to activation of G-proteins in specific rat brain regions. Biochem. Pharmacol. 2000, 59, 1395–1401. [Google Scholar] [CrossRef]
- Maher, C.E.; Martin, T.J.; Childers, S.R. Mechanisms of mu opioid receptor/G-protein desensitization in brain by chronic heroin administration. Life Sci. 2005, 77, 1140–1154. [Google Scholar] [CrossRef] [PubMed]
- Ujcikova, H.; Dlouha, K.; Roubalova, L.; Vosahlikova, M.; Kagan, D.; Svoboda, P. Up-regulation of adenylylcyclases I and II induced by long-term adaptation of rats to morphine fades away 20 days after morphine withdrawal. Biochim. Et Biophys. Acta 2011, 1810, 1220–1229. [Google Scholar] [CrossRef] [PubMed]
- Ujcikova, H.; Brejchova, J.; Vosahlikova, M.; Kagan, D.; Dlouha, K.; Sykora, J.; Merta, L.; Drastichova, Z.; Novotny, J.; Ostasov, P.; et al. Opioid-receptor (OR) signaling cascades in rat cerebral cortex and model cell lines: The role of plasma membrane structure. Physiol. Res. 2014, 63, S165–S176. [Google Scholar] [CrossRef] [PubMed]
- Ujcikova, H.; Eckhardt, A.; Kagan, D.; Roubalova, L.; Svoboda, P. Proteomic analysis of post-nuclear supernatant and percoll-purified membranes prepared from brain cortex of rats exposed to increasing doses of morphine. Proteome Sci. 2014, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Ujcikova, H.; Vosahlikova, M.; Roubalova, L.; Svoboda, P. Proteomic analysis of protein composition of rat forebrain cortex exposed to morphine for 10 days; comparison with animals exposed to morphine and subsequently nurtured for 20 days in the absence of this drug. J. Proteom. 2016, 145, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Ujcikova, H.; Cechova, K.; Jagr, M.; Roubalova, L.; Vosahlikova, M.; Svoboda, P. Proteomic analysis of protein composition of rat hippocampus exposed to morphine for 10 days; comparison with animals after 20 days of morphine withdrawal. PLoS ONE 2020, 15, e0231721. [Google Scholar] [CrossRef] [PubMed]
- Ujcikova, H.; Hejnova, L.; Eckhardt, A.; Roubalova, L.; Novotny, J.; Svoboda, P. Impact of three-month morphine withdrawal on rat brain cortex, hippocampus, striatum and cerebellum: Proteomic and phosphoproteomic studies. Neurochem. Int. 2021, 144, 104975. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Chudapongse, N.; Lee, S.M.; Levin, M.C.; Oh, J.T.; Park, H.J.; Ho, I.K. Proteomic analysis of phosphotyrosyl proteins in morphine-dependent rat brains. Mol. Brain Res. 2005, 133, 58–70. [Google Scholar] [CrossRef]
- Bierczynska-Krzysik, A.; Bonar, E.; Drabik, A.; Noga, M.; Suder, P.; Dylag, T.; Dubin, A.; Kotlinska, J.; Silberring, J. Rat brain proteome in morphine dependence. Neurochem. Int. 2006, 49, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Bierczynska-Krzysik, A.; Pradeep, J.J.P.; Silberring, J.; Kotlinska, J.; Dylag, T.; Cabatic, M.; Lubec, G. Proteomic analysis of rat cerebral cortex, hippocampus and striatum after exposure to morphine. Int. J. Mol. Med. 2006, 18, 775–784. [Google Scholar] [CrossRef]
- Li, K.W.; Jimenez, C.R.; van der Schors, R.C.; Hornshaw, M.P.; Schoffelmeer, A.N.M.; Smit, A.B. Intermittent administration of morphine alters protein expression in rat nucleus accumbens. Proteomics 2006, 6, 2003–2008. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.L.; Lu, G.; Gong, Y.X.; Zhao, L.C.; Chen, J.; Chi, Z.Q.; Yang, Y.M.; Chen, Z.; Li, Q.L.; Liu, J.G. Expression changes of hippocampal energy metabolism enzymes contribute to behavioral abnormalities during chronic morphine treatment. Cell Res. 2007, 17, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Li, J.; Ma, L. Metabolic enzymes link morphine withdrawal with metabolic disorder. Cell Res. 2007, 17, 741–743. [Google Scholar] [CrossRef]
- Bodzon-Kułakowska, A.; Suder, P.; Mak, P.; Bierczynska-Krzysik, A.; Lubec, G.; Walczak, B.; Kotlinska, J.; Lubec, G. Proteomic analysis of striatal neuronal cell cultures after morphine administration. J. Sep. Sci. 2009, 32, 1200–1210. [Google Scholar] [CrossRef] [PubMed]
- Bodzon-Kułakowska, A.; Kułakowski, K.; Drabik, A.; Moszczynski, A.; Silberring, J.; Suder, P. Morphinome—A meta-analysis applied to proteomics. Proteomics 2011, 11, 5–21. [Google Scholar] [CrossRef]
- Antolak, A.; Bodzon-Kułakowska, A.; Cetnarska, E.; Pietruszka, M.; Marszałek-Grabska, M.; Kotlińska, J.; Suder, P. Proteomic data in morphine addiction versus real protein activity: Metabolic enzymes. J. Cell. Biochem. 2017, 118, 4323–4330. [Google Scholar] [CrossRef]
- Bodzon-Kułakowska, A.; Padrtova, T.; Drabik, A.; Ner-Kluza, J.; Antolak, A.; Kułakowski, K.; Suder, P. Morphinome database—The database of proteins altered by morphine administration—An update. J. Proteom. 2019, 190, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Drastichova, Z.; Hejnova, L.; Moravcova, R.; Novotny, J. Proteomic analysis unveils expressional changes in cytoskeleton- and synaptic plasticity-associated proteins in rat brain six months after withdrawal from morphine. Life 2021, 11, 683. [Google Scholar] [CrossRef] [PubMed]
- Marcus, K.; Lelong, C.; Rabilloud, T. What room for two-dimensional gel-based proteomics in a shotgun proteomics world? Proteomes 2020, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Schulenberg, B.; Goodman, T.N.; Aggeler, R.; Capaldi, R.A.; Patton, W.F. Characterization of dynamic and steady-state protein phosphorylation using fluorescent phosphoprotein gel stain and mass spectrometry. Electrophoresis 2004, 25, 2526–2532. [Google Scholar] [CrossRef] [PubMed]
- Eckhardt, A.; Jagr, M.; Pataridis, S.; Miksik, I. Proteomic analysis of human tooth pulp: Proteomics of human tooth. J. Endod. 2014, 40, 1961–1966. [Google Scholar] [CrossRef] [PubMed]
- Jágr, M.; Eckhardt, A.; Pataridis, S.; Foltan, R.; Mysak, J.; Miksik, I. Proteomic analysis of human tooth pulp proteomes—Comparison of caries-resistant and caries-susceptible persons. J. Proteom. 2016, 145, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Dodge, P.W.; Takemori, A.E. Effects of morphine, nalnorphine and pentobarbital alone and combination on cerebral glycolytic substrates and cofactors of rats in vivo. Biochem. Pharmacol. 1972, 21, 287–294. [Google Scholar] [CrossRef]
- Sherman, A.D.; Mitchell, C.L. Effects of morphine and pain on brain intermediary metabolism. Neuropharmacology 1972, 11, 871–877. [Google Scholar] [CrossRef]
- Chuang, D.M.; Hough, C.; Senatorov, V.V. Glyceraldehyde-3-phosphate dehydrogenase, apoptosis, and neurodegenerative diseases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 269–290. [Google Scholar] [CrossRef] [PubMed]
- Tristan, C.; Shahani, N.; Sedlak, T.W.; Sawa, A. The diverse functions of GAPDH: Views from different subcellular compartments. Cell. Signal. 2011, 23, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Landino, L.M.; Hagedorn, T.D.; Kennett, K.L. Evidence for thiol/disulfide exchange reactions between tubulin and glyceraldehyde-3-phosphate dehydrogenase. Cytoskeleton 2014, 71, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Kunjithapatham, R.; Geschwind, J.F.; Devine, L.; Boronina, T.N.; O’Meally, R.N.; Cole, R.N.; Torbenson, R.S.; Ganapathy-Kanniappan, S. Occurence of a multimer high-molecular-weight glyceraldehyde-3-phosphate dehydrogenase in human serum. J. Proteome Res. 2015, 14, 1645–1656. [Google Scholar] [CrossRef] [PubMed]
- Tisdale, E.J.; Talati, N.K.; Artalejo, C.R.; Shisheva, A. GAPDH binds Akt to facilitate cargo transport in the early secretory pathway. Exp. Cell Res. 2016, 349, 310–319. [Google Scholar] [CrossRef]
- Lazarev, V.F.; Guzhova, I.V.; Margulis, B.A. Glyceraldehyde-3-phosphate dehydrogenase is a multifaceted therapeutic target. Pharmaceutics 2020, 12, 416. [Google Scholar] [CrossRef] [PubMed]
- Collel, A.; Green, D.R.; Ricci, J.E. Novel roles for GAPDH in cell death and carcinogenesis. Cell Death Differ. 2009, 16, 1573–1581. [Google Scholar] [CrossRef] [PubMed]
- Goasdoue, K.; Awabdy, D.; Bjorkman, S.T.; Miller, S. Standard loading controls are not reliable for Western blot quantification across brain development or in pathological conditions. Electrophoresis 2016, 37, 630–634. [Google Scholar] [CrossRef]
- Repici, M.; Giorgini, M. DJ-1 in Parkinson’s disease: Clinical insights and therapeutic perspectives. J. Clin. Med. 2019, 8, 1377. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, M.C.T.; Scheller, D.; Schulte, C.; Mesa, I.R.; Colman, P.; Bujac, S.R.; Bell, R.; Berteau, C.; Perez, L.T.; Lachmann, I.; et al. Evaluation of cerebrospinal fluid proteins as potential biomarkers for early stage Parkinson’s disease diagnosis. PLoS ONE 2018, 13, e0206536. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.S.; Geng, W.S.; Wang, Z.Q.; Jia, J.J. Morphine addiction and oxidative stress: The potential effects of thioredoxin-1. Front. Pharmacol. 2020, 11, 82. [Google Scholar] [CrossRef]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Zheng, Q.S.; Pan, J.; Zheng, R.L. Oxidative damage of biomolecules in mouse liver induced by morphine and protected by antioxidants. Basic Clin. Pharmacol. Toxicol. 2004, 95, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Ozmen, I.; Naziroglu, M.; Alici, H.A.; Sahin, F.; Cengiz, M.; Eren, I. Spinal morphine administration reduces the fatty acid contents in spinal cord and brain by increasing oxidative stress. Neurochem. Res. 2007, 32, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Motaghinejad, M.; Karimian, M.; Motaghinejad, O.; Shabab, B.; Yazdani, I.; Fatima, S. Protective effects of various dosage of curcumin against morphine induced apoptosis and oxidative stress in rat isolated hippocampus. Pharmacol. Rep. 2015, 67, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Zaher, A.O.; Mostafa, M.G.; Farghly, H.M.; Hamdy, M.M.; Omran, G.A.; Al-Shaibani, N.K.M. Inhibition of brain oxidative stress and inducible nitric oxide synthase expression by thymoquinone attenuates the development of morphine tolerance and dependence in mice. Eur. J. Pharmacol. 2013, 702, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Skrabalova, J.; Drastichova, Z.; Novotny, J. Morphine as a potential oxidative stress-causing agent. Mini Rev. Org. Chem. 2013, 10, 367–372. [Google Scholar] [CrossRef]
- Mattei, V.; Martellucci, S.; Santilli, F.; Manganelli, V.; Garofalo, T.; Candelise, N.; Caruso, A.; Sorice, M.; Scaccianoce, S.; Misasi, R. Morphine withdrawal modifies prion protein expression in rat hippocampus. PLoS ONE 2017, 12, e0169571. [Google Scholar]
- Bodzon-Kulakowska, A.; Suder, P.; Drabik, A.; Kotlinska, J.H.; Silberring, J. Constant activity of glutamine synthetase after morphine administration versus proteomic results. Anal. Bioanal. Chem. 2010, 398, 2939–2942. [Google Scholar] [CrossRef][Green Version]
- Everitt, B.J.; Robbins, T.W. From the ventral to the dorsal striatum: Devolving views of their roles in drug addiction. Neurosci. Biobehav. Rev. 2013, 37, 1946–1954. [Google Scholar] [CrossRef]
- Yager, L.M.; Garcia, A.F.; Wunsch, A.M.; Ferguson, S.M. The ins and outs of the striatum: Role in drug addiction. Neuroscience 2015, 301, 529–541. [Google Scholar] [CrossRef]
- Shen, S.; Jiang, X.; Li, J.; Straubinger, R.M.; Suarez, M.; Tu, C.; Duan, X.; Thompson, A.C.; Qu, J. Large-scale, ion-current-based proteomic investigation of the rat striatal proteome in a model of short- and long-term cocaine withdrawal. J. Proteome Res. 2016, 15, 1702–1716. [Google Scholar] [CrossRef]
- Miquel, M.; Vazquez-Sanroman, D.; Carbo-Gas, M.; Gil-Miravet, I.; Sanchis-Sequra, C.; Carulli, D.; Manzo, J.; Coria-Avila, G.A. Have we been ignoring the elephant in the room? Seven arguments for considering the cerebellum as part of addiction circuitry. Neurosci. Biobehav. Rev. 2016, 60, 1–11. [Google Scholar] [CrossRef]
- Ranjbar, H.; Soti, M.; Banazadeh, M.; Saleki, K.; Kohlmeier, K.A.; Shabani, M. Addiction and the cerebellum with a focus on actions of opioid receptors. Neurosci. Biobehav. Rev. 2021, 131, 229–247. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot | Accession | Gene | Protein Name | Mascot | Matched | SC a | MW b | pI c | Change | p Value |
|---|---|---|---|---|---|---|---|---|---|---|
| Number | Score | Peptides | [%] | (kDa) | (Fold) | |||||
| (a) HIPPOCAMPUS Pro-Q® Diamond staining | ||||||||||
| 1 | P0DP29 | Calm1 | Calmodulin-1 | 2299.5 | 28 | 93.3 | 16.8 | 3.9 | ↑2.3 | 0.0316 |
| 2 | P63102 | Ywhaz | 14-3-3 protein zeta/delta | 1654.2 | 25 | 74.7 | 27.8 | 4.6 | ↑2.4 | 0.0014 |
| 3 | Q6P9V9 | Tuba1b | Tubulin alpha-1B chain | 1866.7 | 27 | 57.9 | 50.1 | 4.8 | ↑3.2 | 0.0424 |
| 4 | O35814 | Stip1 | Stress-induced phosphoprotein 1 | 1728.2 | 32 | 39.4 | 62.5 | 6.4 | ↓3.5 | 0.0309 |
| 5 | P16617 | Pgk1 | Phosphoglycerate kinase 1 | 2026.8 | 36 | 57.8 | 44.5 | 9.0 | ↓2.2 | 0.0195 |
| 6 | P05065 | Aldoa | Fructose-bisphosphate aldolase A | 2243.8 | 35 | 80.2 | 39.3 | 9.2 | ↓2.7 | 0.0084 |
| (b) HIPPOCAMPUS Colloidal Coommassie Blue staining | ||||||||||
| 1 | P35704 | Prdx2 | Peroxiredoxin-2 | 429.6 | 8 | 33.8 | 21.8 | 5.2 | ↓3.1 | 0.0036 |
| 2 | P31044 | Pebp1 | Phosphatidylethanolamine-binding | 988.5 | 13 | 62.6 | 20.8 | 5.4 | ↓2.6 | 0.0004 |
| protein 1 | ||||||||||
| 3 | Q00981 | Uchl1 | Ubiquitin carboxyl-terminal hydrolase | 1462.4 | 24 | 59.6 | 24.8 | 5.0 | ↓2.0 | 0.0394 |
| isozyme L1 | ||||||||||
| 4 | F8WFM2 | Napb | Beta-soluble NSF attachment protein | 1276.3 | 20 | 55.4 | 33.5 | 5.2 | ↓2.4 | 0.0015 |
| 5 | P54311 | Gnb1 | Guanine nucleotide-binding protein | 728.3 | 12 | 33.2 | 37.4 | 5.6 | ↓2.4 | 0.0045 |
| G(I)/G(S)/G(T) subunit beta-1 | ||||||||||
| 6 | O35179 | Sh3gl2 | Endophilin-A1 | 1368.9 | 28 | 34.4 | 39.9 | 5.1 | ↓7.8 | 0.0363 |
| 7 | P60711 | Actb | Actin, cytoplasmic 1 | 2492.6 | 38 | 62.7 | 41.7 | 5.2 | ↓1.8 | 0.0293 |
| 8 | P07335 | Ckb | Creatine kinase B-type | 2008.6 | 27 | 57 | 42.7 | 5.3 | ↓1.6 | 0.0402 |
| 9 | P07335 | Ckb | Creatine kinase B-type | 2337.6 | 34 | 65.4 | 42.7 | 5.3 | ↓2.3 | 0.0026 |
| 10 | G3V7C6 | Tubb4b | Tubulin beta chain | 2983.9 | 37 | 49.8 | 61.1 | 4.6 | ↓1.9 | 0.0358 |
| 11 | P23565 | Ina | Alpha-internexin | 2688.1 | 41 | 75.6 | 56.1 | 5.1 | ↓2.4 | 0.0117 |
| 12 | F1M953 | Hspa9 | Stress-70 protein, mitochondrial | 2772.9 | 43 | 41.7 | 73.7 | 5.8 | ↓3.1 | 0.0127 |
| 13 | G3V7C6 | Tubb4b | Tubulin beta chain | 1809.5 | 23 | 40.5 | 61.1 | 4.6 | ↓6.1 | 0.0175 |
| 14 | P31399 | Atp5pd | ATP synthase subunit d, mitochondrial | 1680.6 | 23 | 67.7 | 18.8 | 6.2 | ↑6.7 | 0.0011 |
| 15 | O88767 | Park7 | Parkinson disease protein 7 homolog | 1106.3 | 15 | 70.4 | 20 | 6.4 | ↑2.0 | 0.0324 |
| 16 | P48500 | Tpi1 | Triosephosphate isomerase | 829.1 | 12 | 53 | 26.8 | 7.7 | ↑1.9 | 0.0348 |
| (c) CORTEX Pro-Q® Diamond staining | ||||||||||
| 1 | P85108 | Tubb2a | Tubulin beta-2A chain | 3071.4 | 37 | 73 | 49.9 | 4.6 | ↓2.4 | 0.0103 |
| 2 | P68370 | Tuba1a | Tubulin alpha-1A chain | 1538.3 | 21 | 47 | 50.1 | 4.8 | ↓3.7 | 0.0406 |
| 3 | P60711 | Actb | Actin, cytoplasmic 1 | 1532.3 | 23 | 50.9 | 41.7 | 5.2 | ↓2.7 | 0.0284 |
| (d) CORTEX Colloidal Coommassie Blue staining | ||||||||||
| 1 | P0DP29 | Calm1 | Calmodulin-1 | 1670.9 | 18 | 74.5 | 16.8 | 3.9 | ↓2.0 | 0.0478 |
| 2 | P37377 | Snca | Alpha-synuclein | 1287.2 | 16 | 80 | 14.5 | 4.6 | ↓1.4 | 0.0114 |
| 3 | Q99NA5 | Idh3a | Isocitrate dehydrogenase [NAD] subunit | 830.1 | 9 | 28 | 39.6 | 6.5 | ↑2.4 | 0.0252 |
| alpha, mitochondrial | ||||||||||
| Spot | Accession | Protein Name | Change | Subcellular Localization | GO-Molecular Functions, Biological Processes |
|---|---|---|---|---|---|
| Number | (Fold) | ||||
| (a) HIPPOCAMPUS Pro-Q® Diamond staining | |||||
| 1 | P0DP29 | Calmodulin-1 | ↑2.3 | Cytoplasm, cytoskeleton | Calcium-mediated signaling, activation |
| of adenylate cyclase activity, regulation | |||||
| of cytokinesis | |||||
| 2 | P63102 | 14-3-3 protein zeta/delta | ↑2.4 | Cytoplasm, melanosome | Signal transducing adaptor protein |
| 3 | Q6P9V9 | Tubulin alpha-1B chain | ↑3.2 | Cytoplasm, cytoskeleton | Cell shape and movement |
| 4 | O35814 | Stress-induced phosphoprotein 1 | ↓3.5 | Cytoplasm, nucleus | Chaperone binding |
| 5 | P16617 | Phosphoglycerate kinase 1 | ↓2.2 | Cytoplasm | Energy metabolism (glycolysis) |
| 6 | P05065 | Fructose-bisphosphate aldolase A | ↓2.7 | Cytoplasm | Energy metabolism (glycolysis) |
| (b) HIPPOCAMPUS Colloidal Coommassie Blue staining | |||||
| 1 | P35704 | Peroxiredoxin-2 | ↓3.1 | Cytoplasm | Antioxidant, response to oxidative stress |
| 2 | P31044 | Phosphatidylethanolamine-binding | ↓2.6 | Cytoplasm, cell membrane | Hippocampus development, aging, response |
| protein 1 | to oxidative stress, MAPK cascade | ||||
| 3 | Q00981 | Ubiquitin carboxyl-terminal hydrolase | ↓2.0 | Cytoplasm, endoplasmic reticulum | Protein ubiquitination, axonogenesis |
| isozyme L1 | |||||
| 4 | F8WFM2 | Beta-soluble NSF attachment protein | ↓2.4 | Cell membrane | ER-Golgi transport, protein transport |
| 5 | P54311 | Guanine nucleotide-binding protein | ↓2.4 | Cell membrane, cytoplasm | Signal transducer |
| G(I)/G(S)/G(T) subunit beta-1 | |||||
| 6 | O35179 | Endophilin-A1 | ↓7.8 | Cytoplasm, endosome, cell membrane | Endocytosis, regulation of receptor internalization |
| 7 | P60711 | Actin, cytoplasmic 1 | ↓1.8 | Cytoplasm, cytoskeleton, nucleus | Cell shape and movement |
| 8,9 | P07335 | Creatine kinase B-type | ↓1.6, ↓2.3 | Cytoplasm | Brain development, creatine metabolism |
| 10,13 | G3V7C6 | Tubulin beta chain | ↓1.9, ↓6.1 | Cytoplasm, cytoskeleton | Cell shape and movement |
| 11 | P23565 | Alpha-internexin | ↓2.4 | Cytoplasm, cytoskeleton | Cytoskeleton organization, developmental protein |
| 12 | F1M953 | Stress-70 protein, mitochondrial | ↓3.1 | Mitochondrion | Chaperone |
| 14 | P31399 | ATP synthase subunit d, mitochondrial | ↑6.7 | Mitochondrion | ATP metabolic process, hydrogen ion transport |
| 15 | O88767 | Parkinson disease protein 7 homolog | ↑2.0 | Cell membrane, cytoplasm, nucleus, | Chaperone, aging, inflammatory response, stress |
| endoplasmic reticulum | response, negative regulation of apoptosis | ||||
| 16 | P48500 | Triosephosphate isomerase | ↑1.9 | Cytoplasm | Energy metabolism (glycolysis) |
| (c) CORTEX Pro-Q® Diamond staining | |||||
| 1 | P85108 | Tubulin beta-2A chain | ↓2.4 | Cytoplasm, cytoskeleton | Cell shape and movement |
| 2 | P68370 | Tubulin alpha-1A chain | ↓3.7 | Cytoplasm, cytoskeleton | Cell shape and movement |
| 3 | P60711 | Actin, cytoplasmic 1 | ↓2.7 | Cytoplasm, cytoskeleton, nucleus | Cell shape and movement |
| (d) CORTEX Colloidal Coommassie Blue staining | |||||
| 1 | P0DP29 | Calmodulin-1 | ↓2.0 | Cytoplasm, cytoskeleton | Calcium-mediated signaling, activation of adenylate cyclase activity, regulation of cytokinesis |
| 2 | P37377 | Alpha-synuclein | ↓1.4 | Cytoplasm, cell membrane, nucleus, secreted | Chaperone, response to oxidative stress, regulation of synaptic vesicle trafficking, regulation of neurotransmitter release |
| 3 | Q99NA5 | Isocitrate dehydrogenase [NAD] subunit alpha, mitochondrial | ↑2.4 | Mitochondrion | Krebs cycle |
| Pathways | Enrichment FDR | Pathway | Genes | Name of Genes in List |
|---|---|---|---|---|
| Genes | in List | |||
| Phosphorus metabolic process | 0.000454130956700965 | 2813 | 12 | Atp5pd, Aldoa, Uchl1, Prdx2, Calm1, Sh3gl2, Ywhaz, Ckb, Tpi1, Park7, Pgk1, Actb |
| Phosphate-containing compound metabolic process | 0.000454130956700965 | 2798 | 12 | Atp5pd, Aldoa, Uchl1, Prdx2, Calm1, Sh3gl2, Ywhaz, Ckb, Tpi1, Park7, Pgk1, Actb |
| Phosphorylation | 0.00109269868079895 | 2072 | 10 | Aldoa, Uchl1, Prdx2, Sh3gl2, Ywhaz, Tpi1, Park7, Pgk1, Actb, Atp5pd |
| ATP metabolic process | 0.00109269868079895 | 272 | 5 | Atp5pd, Aldoa, Tpi1, Park7, Pgk1 |
| Methylglyoxal metabolic process | 0.001813238441941 | 5 | 2 | Park7, Aldoa |
| Generation of precursor metabolites and energy | 0.00432203205106559 | 397 | 5 | Aldoa, Tpi1, Park7, Pgk1, Atp5pd |
| Nucleoside phosphate metabolic process | 0.00464706222526931 | 428 | 5 | Atp5pd, Aldoa, Tpi1, Park7, Pgk1 |
| Nucleotide metabolic process | 0.00464706222526931 | 419 | 5 | Atp5pd, Aldoa, Tpi1, Park7, Pgk1 |
| Postsynaptic cytoskeleton organization | 0.00556835599926725 | 13 | 2 | Ina, Actb |
| Organophosphate metabolic process | 0.00556835599926725 | 820 | 6 | Atp5pd, Aldoa, Ckb, Tpi1, Park7, Pgk1 |
| Establishment of localization in cell | 0.00556835599926725 | 1630 | 8 | Calm1, Ywhaz, Uchl1, Napb, Park7, Hspa9, Actb, Atp5pd |
| Nucleobase-containing small molecule metabolic process | 0.00556835599926725 | 473 | 5 | Atp5pd, Aldoa, Tpi1, Park7, Pgk1 |
| Postsynaptic actin cytoskeleton organization | 0.00556835599926725 | 13 | 2 | Ina, Actb |
| Pyruvate metabolic process | 0.00698656482715899 | 121 | 3 | Aldoa, Tpi1, Pgk1 |
| Glycolytic process | 0.00698656482715899 | 102 | 3 | Aldoa, Tpi1, Pgk1 |
| Nucleoside diphosphate phosphorylation | 0.00698656482715899 | 120 | 3 | Aldoa, Tpi1, Pgk1 |
| ATP generation from ADP | 0.00698656482715899 | 103 | 3 | Aldoa, Tpi1, Pgk1 |
| Purine nucleoside diphosphate metabolic process | 0.00698656482715899 | 113 | 3 | Aldoa, Tpi1, Pgk1 |
| Purine ribonucleoside diphosphate metabolic process | 0.00698656482715899 | 113 | 3 | Aldoa, Tpi1, Pgk1 |
| Ribonucleoside diphosphate metabolic process | 0.00698656482715899 | 116 | 3 | Aldoa, Tpi1, Pgk1 |
| Removal of superoxide radicals | 0.00698656482715899 | 18 | 2 | Prdx2, Park7 |
| Response to chemical | 0.00698656482715899 | 4423 | 12 | Ywhaz, Gnb1, Uchl1, Prdx2, Calm1, Park7, Ina, Stip1, Actb, Tuba1b, Atp5pd, Aldoa |
| Response to superoxide | 0.00698656482715899 | 21 | 2 | Prdx2, Park7 |
| Response to oxygen radical | 0.00698656482715899 | 21 | 2 | Prdx2, Park7 |
| ADP metabolic process | 0.00698656482715899 | 108 | 3 | Aldoa, Tpi1, Pgk1 |
| Nucleotide phosphorylation | 0.00698656482715899 | 122 | 3 | Aldoa, Tpi1, Pgk1 |
| Cellular response to chemical stimulus | 0.00698656482715899 | 2498 | 9 | Gnb1, Uchl1, Prdx2, Park7, Ina, Stip1, Actb, Tuba1b, Atp5pd |
| Cellular response to oxygen radical | 0.00698656482715899 | 19 | 2 | Prdx2, Park7 |
| Cellular response to superoxide | 0.00698656482715899 | 19 | 2 | Prdx2, Park7 |
| Purine ribonucleotide metabolic process | 0.00726512653650191 | 327 | 4 | Atp5pd, Aldoa, Tpi1, Pgk1 |
| Pathways | Enrichment FDR | Pathway | Genes | Name of Genes in List |
|---|---|---|---|---|
| Genes | in List | |||
| Presynaptic endocytosis | 0.0000317791916695507 | 41 | 3 | Snca, Actb, Calm1 |
| Synaptic vesicle endocytosis | 0.0000317791916695507 | 41 | 3 | Snca, Actb, Calm1 |
| Regulation of norepinephrine uptake | 0.0000317791916695507 | 2 | 2 | Snca, Actb |
| Synaptic vesicle recycling | 0.0000411603443103439 | 49 | 3 | Snca, Actb, Calm1 |
| Norepinephrine uptake | 0.00014973418795375 | 6 | 2 | Snca, Actb |
| Catecholamine uptake | 0.000648303871273433 | 13 | 2 | Snca, Actb |
| Synaptic vesicle cycle | 0.000655582495341169 | 147 | 3 | Snca, Actb, Calm1 |
| Vesicle-mediated transport in synapse | 0.000739942307588717 | 160 | 3 | Snca, Actb, Calm1 |
| Norepinephrine transport | 0.000753222666338899 | 17 | 2 | Snca, Actb |
| Regulation of transmembrane transporter activity | 0.00113861317223078 | 199 | 3 | Calm1, Snca, Actb |
| Spot | Accession | Gene | Protein Name | Mascot | Matched | SC a | MW b | pI c | Change | p Value |
|---|---|---|---|---|---|---|---|---|---|---|
| Number | Score | Peptides | [%] | (kDa) | (Fold) | |||||
| (a) STRIATUM Colloidal Coommassie Blue staining | ||||||||||
| 1 | P0DP29 | Calm1 | Calmodulin-1 | 1050.8 | 15 | 74.5 | 16.8 | 3.9 | ↓2.3 | 0.0079 |
| 2 | P35704 | Prdx2 | Peroxiredoxin-2 | 388.2 | 7 | 31.3 | 21.8 | 5.2 | ↑1.8 | 0.0214 |
| 3 | P42123 | Ldhb | L-lactate dehydrogenase B chain | 1159.9 | 20 | 41.3 | 36.6 | 5.6 | ↓1.4 | 0.0483 |
| 4 | O88989 | Mdh1 | Malate dehydrogenase, cytoplasmic | 808.2 | 14 | 44.9 | 36.5 | 6.2 | ↓1.8 | 0.0326 |
| 5 | P60711 | Actb | Actin, cytoplasmic 1 | 1304.1 | 24 | 44.5 | 41.7 | 5.2 | ↑3.3 | 0.0258 |
| 6 | P63018 | Hspa8 | Heat shock cognate 71 kDa protein | 1336.6 | 23 | 31.6 | 70.8 | 5.2 | ↑2.5 | 0.0050 |
| 7 | P02770 | Alb | Albumin | 651.8 | 12 | 18.3 | 68.7 | 6.1 | ↓2.4 | 0.0115 |
| 8 | P47942 | Dpysl2 | Dihydropyrimidinase-related protein 2 | 1104.7 | 17 | 35.5 | 62.2 | 5.9 | ↓1.7 | 0.0352 |
| 9 | P47942 | Dpysl2 | Dihydropyrimidinase-related protein 2 | 1997.6 | 31 | 56.3 | 62.2 | 5.9 | ↓2.0 | 0.0244 |
| 10 | P19804 | Nme2 | Nucleoside diphosphate kinase B | 575.2 | 11 | 59.2 | 17.3 | 7.8 | ↓2.0 | 0.0458 |
| 11 | M0R757 | LOC100360413 | Elongation factor 1-alpha | 548.1 | 12 | 21.2 | 50.1 | 9.7 | ↓3.1 | 0.0030 |
| (b) CEREBELLUM Colloidal Coommassie Blue staining | ||||||||||
| 1 | P61983 | Ywhag | 14-3-3 protein gamma | 1065.1 | 20 | 49.4 | 28.3 | 4.7 | ↓3.8 | 0.0451 |
| 2 | P62260 | Ywhae | 14-3-3 protein epsilon | 1373.3 | 23 | 49.8 | 29.2 | 4.5 | ↓2.7 | 0.0214 |
| 3 | P85969 | Napb | Beta-soluble NSF attachment protein | 418.7 | 9 | 26.6 | 33.4 | 5.2 | ↓1.6 | 0.0172 |
| 4 | P54311 | Gnb1 | Guanine nucleotide-binding protein | 497.7 | 9 | 22.9 | 37.4 | 5.6 | ↓4.4 | 0.0035 |
| G(I)/G(S)/G(T) subunit beta-1 | ||||||||||
| 5 | P42123 | Ldhb | L-lactate dehydrogenase B chain | 1288.1 | 22 | 43.7 | 36.6 | 5.6 | ↓1.9 | 0.0471 |
| 6 | P07335 | Ckb | Creatine kinase B-type | 1531.4 | 22 | 53.8 | 42.7 | 5.3 | ↓1.5 | 0.0246 |
| 7 | Q6P9V9 | Tuba1b | Tubulin alpha-1B chain | 1209.4 | 21 | 53.7 | 50.1 | 4.8 | ↓2.1 | 0.0041 |
| 8 | P63018 | Hspa8 | Heat shock cognate 71 kDa protein | 2437.8 | 37 | 45.5 | 70.8 | 5.2 | ↓2.1 | 0.0072 |
| 9 | P47942 | Dpysl2 | Dihydropyrimidinase-related protein 2 | 1684.2 | 25 | 48.8 | 62.2 | 5.9 | ↓1.8 | 0.0174 |
| 10 | P09117 | Aldoc | Fructose-bisphosphate aldolase C | 1956.9 | 28 | 59 | 39.3 | 6.8 | ↓3.0 | 0.0053 |
| 11 | P48500 | Tpi1 | Triosephosphate isomerase | 859.3 | 12 | 50.6 | 26.8 | 7.7 | ↓2.2 | 0.0200 |
| Spot | Accession | Protein Name | Change | Subcellular Localization | GO-Molecular Functions, Biological Processes |
|---|---|---|---|---|---|
| Number | (Fold) | ||||
| (a) STRIATUM Colloidal Coommassie Blue staining | |||||
| 1 | P0DP29 | Calmodulin-1 | ↓2.3 | Cytoplasm, cytoskeleton | Calcium-mediated signaling, activation |
| of adenylate cyclase activity, regulation | |||||
| of cytokinesis | |||||
| 2 | P35704 | Peroxiredoxin-2 | ↑1.8 | Cytoplasm | Antioxidant, response to oxidative stress |
| 3 | P42123 | L-lactate dehydrogenase B chain | ↓1.4 | Cytoplasm, mitochondrion | Pyruvate metabolic process |
| 4 | O88989 | Malate dehydrogenase, cytoplasmic | ↓1.8 | Cytoplasm | Krebs cycle |
| 5 | P60711 | Actin, cytoplasmic 1 | ↑3.3 | Cytoplasm, cytoskeleton, nucleus | Cell shape and movement |
| 6 | P63018 | Heat shock cognate 71 kDa protein | ↑2.5 | Cell membrane, cytoplasm, nucleus | Protein folding, RNA processing, aging |
| 7 | P02770 | Albumin | ↓2.4 | Secreted | Transporter, apoptosis |
| 8,9 | P47942 | Dihydropyrimidinase-related protein 2 | ↓1.7,↓2.0 | Cytoplasm, cytoskeleton, cell | Brain development, neurogenesis, cell |
| membrane | movement | ||||
| 10 | P19804 | Nucleoside diphosphate kinase B | ↓2.0 | Cytoplasm, nucleus | Nucleotide metabolism, transcription |
| 11 | M0R757 | Elongation factor 1-alpha | ↓3.1 | Cell membrane, cytoplasm, nucleus | Translation |
| (b) CEREBELLUM Colloidal Coommassie Blue staining | |||||
| 1 | P61983 | 14-3-3 protein gamma | ↓3.8 | Cytoplasm | Signal transducing adaptor protein |
| 2 | P62260 | 14-3-3 protein epsilon | ↓2.6 | Cytoplasm, nucleus | Signal transducing adaptor protein, brain |
| development | |||||
| 3 | P85969 | Beta-soluble NSF attachment protein | ↓1.6 | Cell membrane | ER-Golgi transport, protein transport |
| 4 | P54311 | Guanine nucleotide-binding protein | ↓4.4 | Cell membrane, cytoplasm | Signal transduction |
| G(I)/G(S)/G(T) subunit beta-1 | |||||
| 5 | P42123 | L-lactate dehydrogenase B chain | ↓1.9 | Cytoplasm, cell membrane, | Pyruvate metabolic process |
| mitochondrion | |||||
| 6 | P07335 | Creatine kinase B-type | ↓1.5 | Cytoplasm | Brain development, creatine metabolism |
| 7 | Q6P9V9 | Tubulin alpha-1B chain | ↓2.1 | Cytoplasm, cytoskeleton | Cell shape and movement |
| 8 | P63018 | Heat shock cognate 71 kDa protein | ↓2.1 | Cell membrane, cytoplasm, nucleus | Protein folding, RNA processing, aging |
| 9 | P47942 | Dihydropyrimidinase-related protein 2 | ↓1.8 | Cytoplasm, cytoskeleton, cell | Brain development, neurogenesis, cell |
| membrane | movement | ||||
| 10 | P09117 | Fructose-bisphosphate aldolase C | ↓3.0 | Cytoplasm | Energy metabolism (glycolysis), aging, apoptosis |
| 11 | P48500 | Triosephosphate isomerase | ↓2.2 | Cytoplasm | Energy metabolism (glycolysis) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ujcikova, H.; Eckhardt, A.; Hejnova, L.; Novotny, J.; Svoboda, P. Alterations in the Proteome and Phosphoproteome Profiles of Rat Hippocampus after Six Months of Morphine Withdrawal: Comparison with the Forebrain Cortex. Biomedicines 2022, 10, 80. https://doi.org/10.3390/biomedicines10010080
Ujcikova H, Eckhardt A, Hejnova L, Novotny J, Svoboda P. Alterations in the Proteome and Phosphoproteome Profiles of Rat Hippocampus after Six Months of Morphine Withdrawal: Comparison with the Forebrain Cortex. Biomedicines. 2022; 10(1):80. https://doi.org/10.3390/biomedicines10010080
Chicago/Turabian StyleUjcikova, Hana, Adam Eckhardt, Lucie Hejnova, Jiri Novotny, and Petr Svoboda. 2022. "Alterations in the Proteome and Phosphoproteome Profiles of Rat Hippocampus after Six Months of Morphine Withdrawal: Comparison with the Forebrain Cortex" Biomedicines 10, no. 1: 80. https://doi.org/10.3390/biomedicines10010080
APA StyleUjcikova, H., Eckhardt, A., Hejnova, L., Novotny, J., & Svoboda, P. (2022). Alterations in the Proteome and Phosphoproteome Profiles of Rat Hippocampus after Six Months of Morphine Withdrawal: Comparison with the Forebrain Cortex. Biomedicines, 10(1), 80. https://doi.org/10.3390/biomedicines10010080