
Ameliorating Fibrosis in Murine and Human Tissues with END55, an Endostatin-Derived Fusion Protein Made in Plants
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Measurement of END55 Targets to Develop a Potency Assay
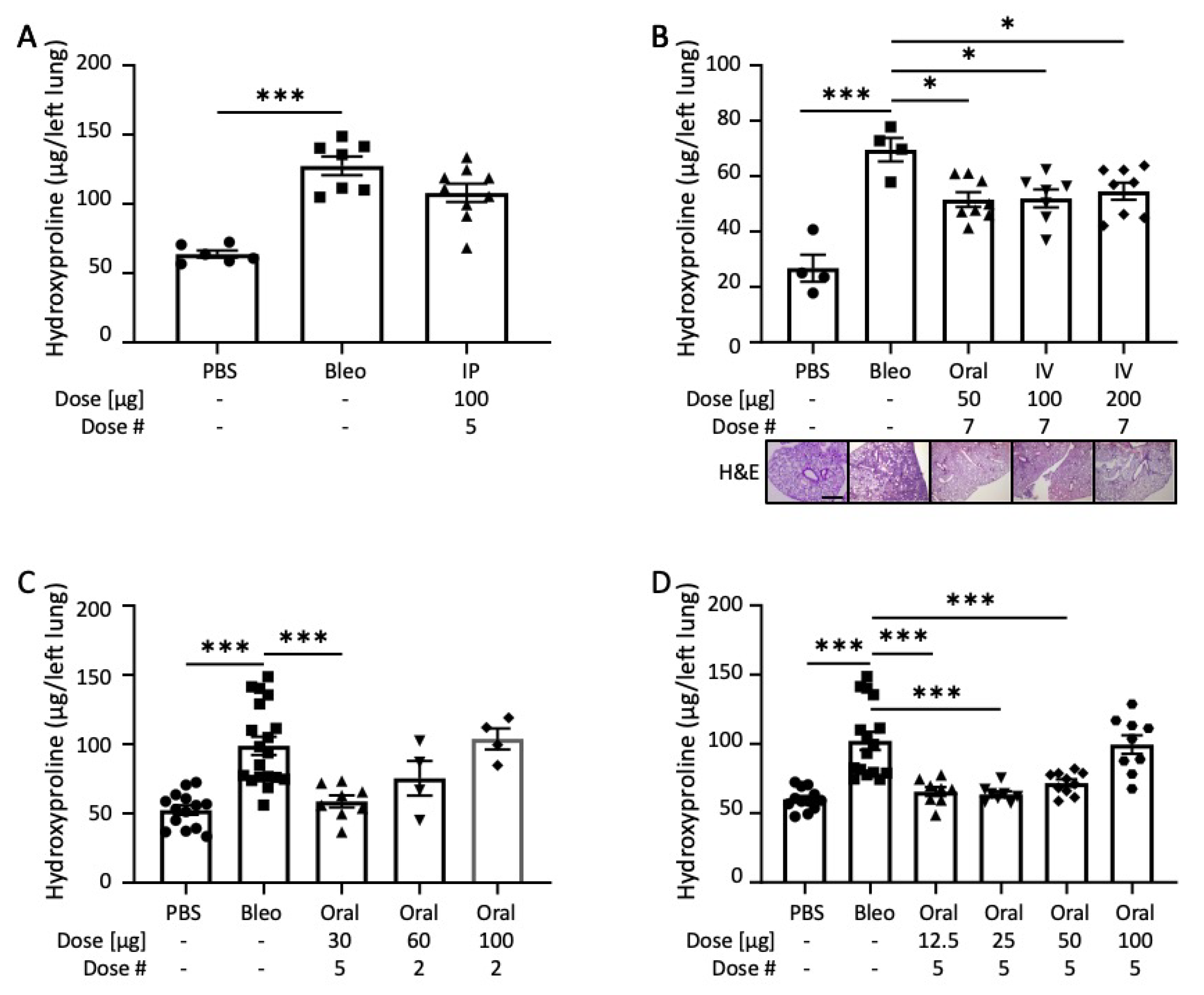
2.2. END55 Ameliorates Bleomycin-Induced Lung Fibrosis
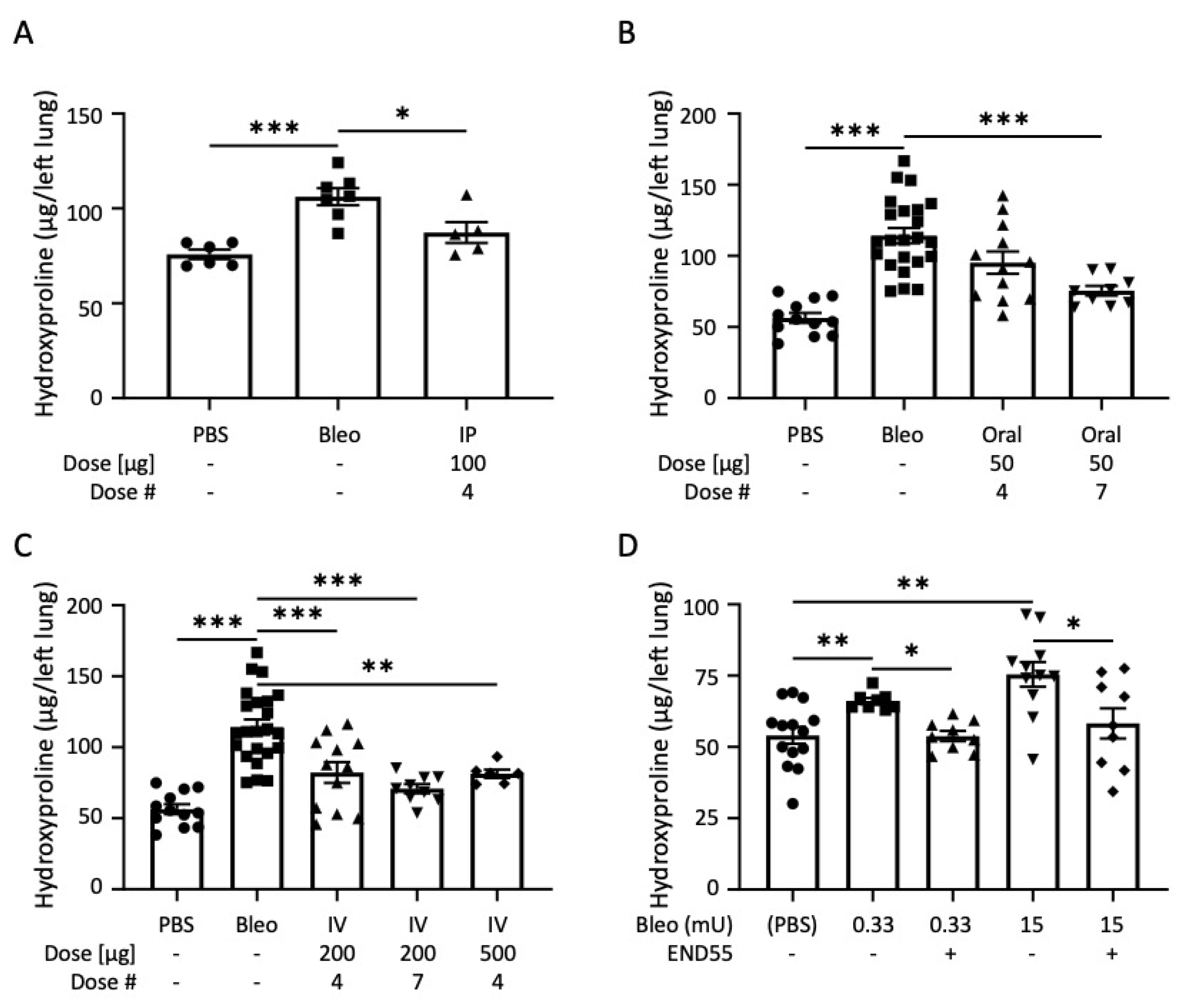
2.3. END55 Reverses Bleomycin-Induced Lung Fibrosis
2.4. END55 Reverses Established Bleomycin Lung Fibrosis
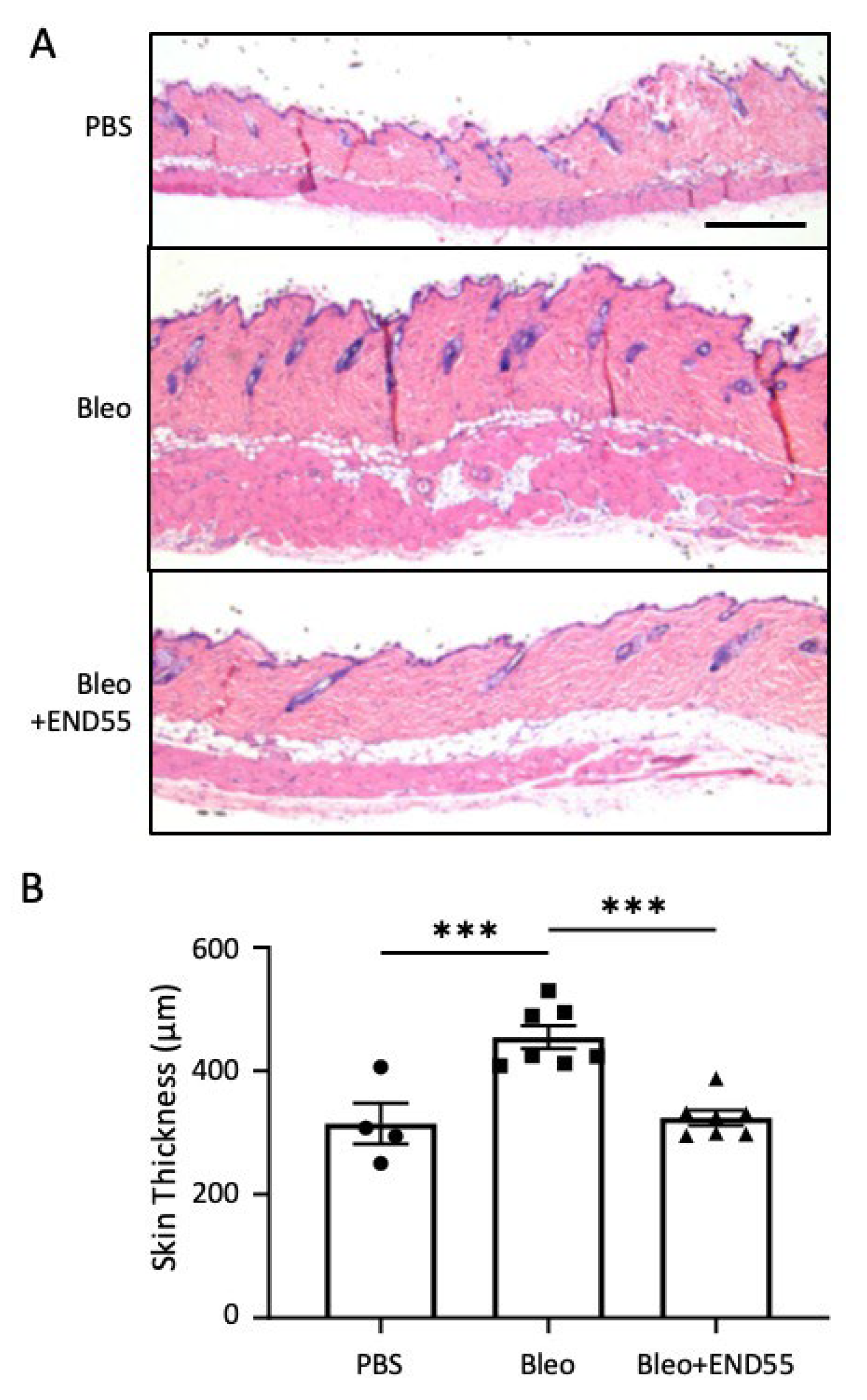
2.5. END55 Reduces Skin Thickness in Mice with Established Fibrosis
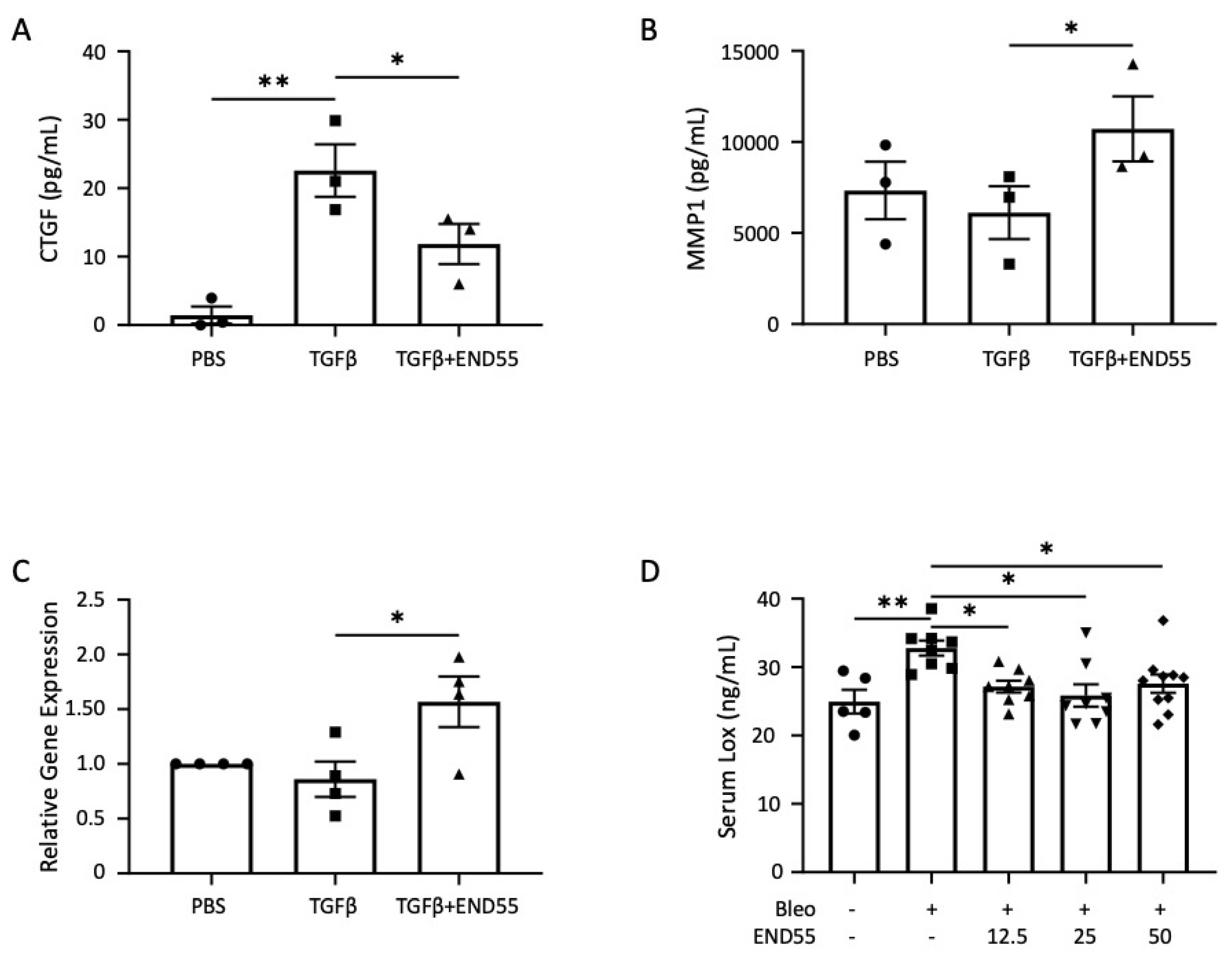
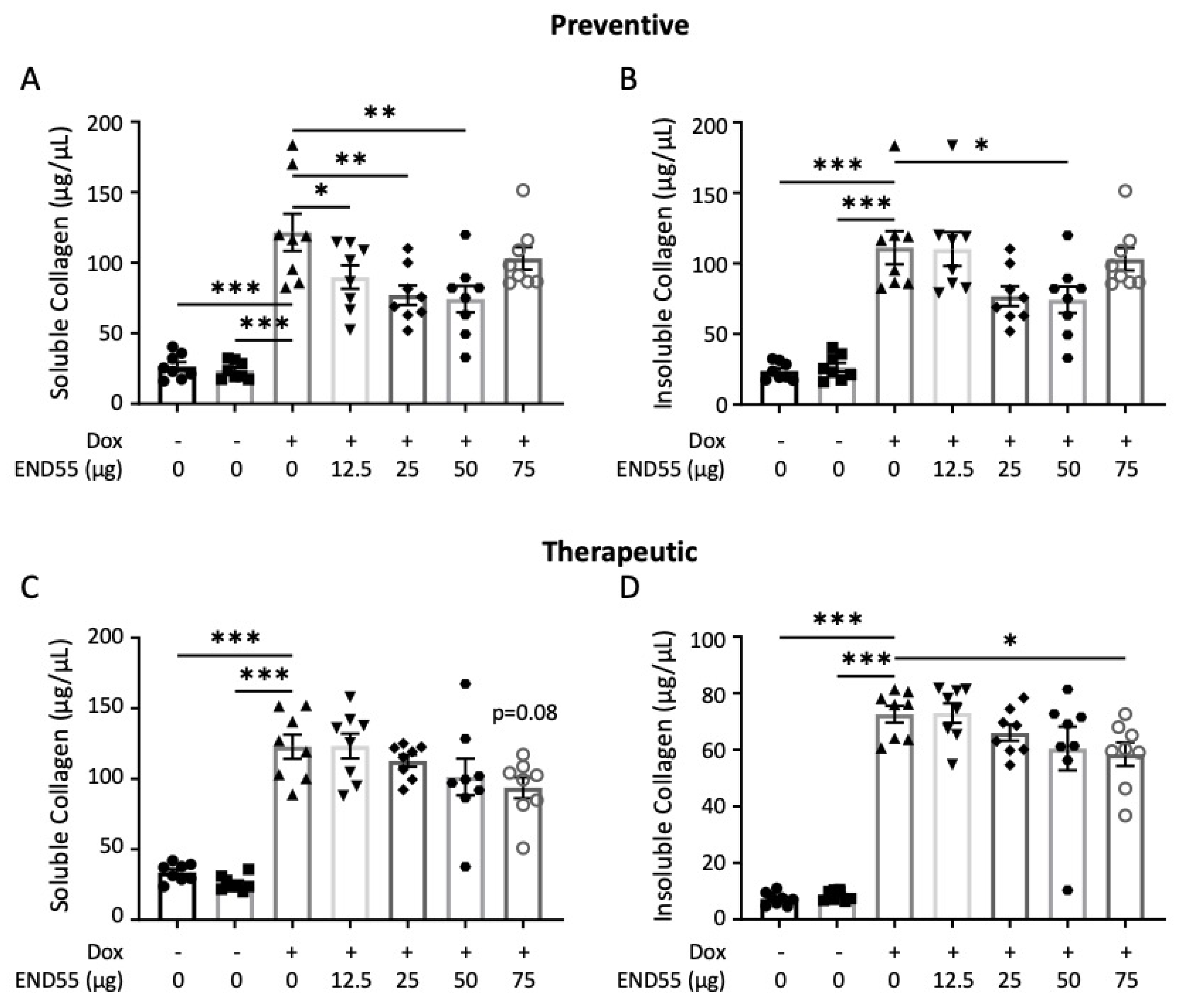
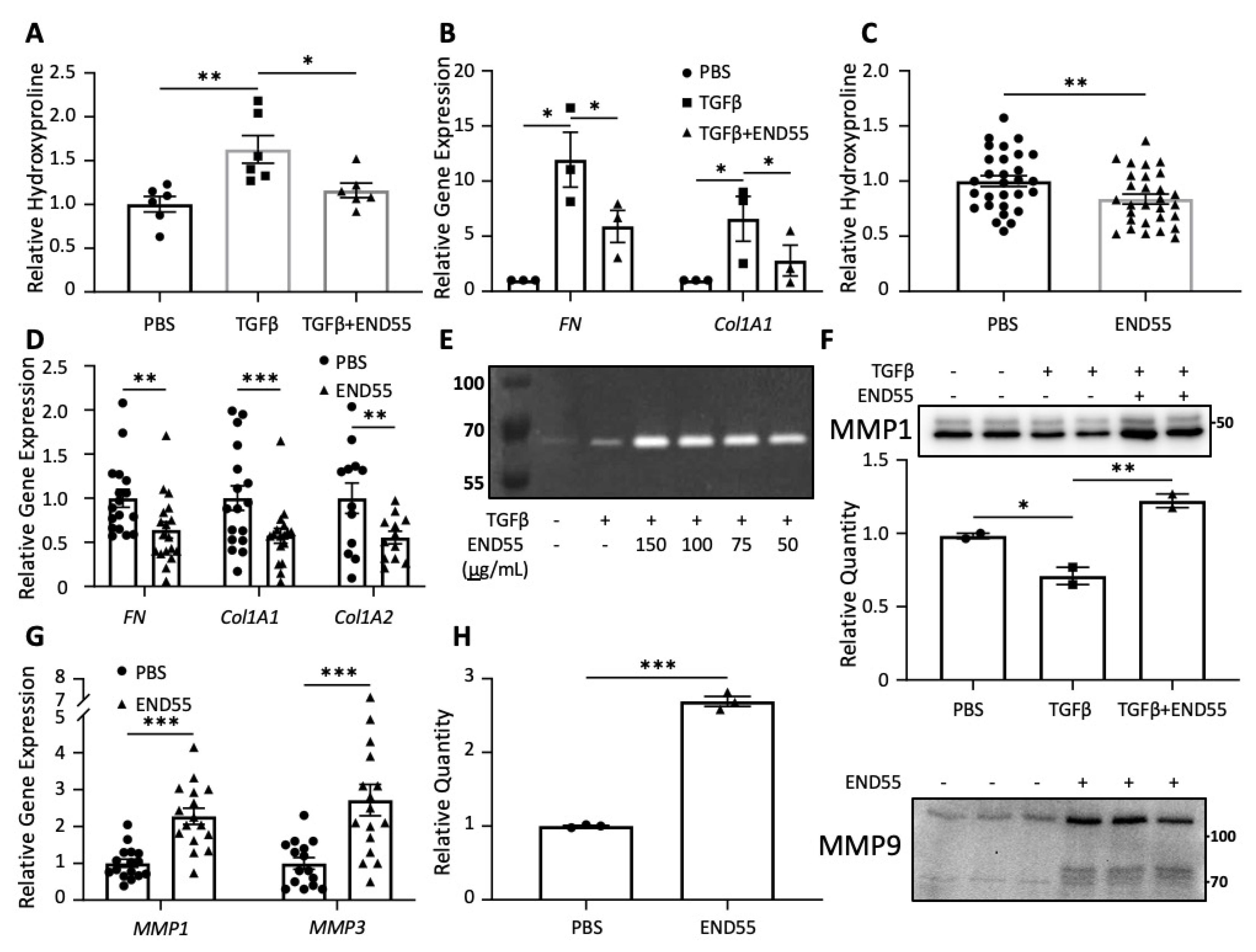
2.6. END55 Prevents and Ameliorates TGFβ-Induced Fibrosis In Vivo
2.7. END55 Reduces Markers of Fibrosis in Human Lung Tissue
2.8. END55 Increases Levels of Matrix Metalloproteases
2.9. END55 Decreases Fibrosis in Human Skin
3. Discussion
4. Materials and Methods
4.1. Peptide
4.2. Animal Studies
4.3. Bleomycin-Fibrosis Model
4.4. Collagen Assays
4.5. Histology
4.6. In Vitro and Ex Vivo Assays
4.7. RNA Extraction and Real Time Polymerase Chain Reaction
4.8. Potency Assays
4.9. Zymography
4.10. Immunoblotting
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Declaration of Interests
References
- Foundation PF. Available online: https://www.pulmonaryfibrosis.org/ (accessed on 4 June 2020).
- Association AL. Available online: https://www.lung.org/ (accessed on 4 June 2020).
- Wynn, T.A. Common and unique mechanisms regulate fibrosis in various fibroproliferative diseases. J. Clin. Investig. 2007, 117, 524–529. [Google Scholar] [CrossRef] [Green Version]
- Herzog, E.L.; Mathur, A.; Tager, A.M.; Feghali-Bostwick, C.; Schneider, F.; Varga, J. Review: Interstitial lung disease associated with systemic sclerosis and idiopathic pulmonary fibrosis: How similar and distinct? Arthritis Rheumatol. 2014, 66, 1967–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghu, G.; Chen, S.-Y.; Hou, Q.; Yeh, W.-S.; Collard, H.R. Incidence and prevalence of idiopathic pulmonary fibrosis in US adults 18–64 years old. Eur. Respir. J. 2016, 48, 179–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perelas, A.; Silver, R.M.; Arrossi, A.V.; Highland, K.B. Systemic sclerosis-associated interstitial lung disease. Lancet Respir. Med. 2020, 8, 304–320. [Google Scholar] [CrossRef]
- Garrett, S.M.; Frost, D.B.; Feghali-Bostwick, C. The mighty fibroblast and its utility in scleroderma research. J. Scleroderma Relat. Disord. 2017, 2, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Watanabe, T.; Frost, D.B.; Mlakar, L.; Heywood, J.; da Silveira, W.A.; Hardiman, G.; Feghali-Bostwick, C. A Human Skin Model Recapitulates Systemic Sclerosis Dermal Fibrosis and Identifies COL22A1 as a TGFβ Early Response Gene that Mediates Fibroblast to Myofibroblast Transition. Genes 2019, 10, 75. [Google Scholar] [CrossRef] [Green Version]
- Denton, C.P.; Merkel, P.A.; Furst, D.E.; Khanna, D.; Emery, P.; Hsu, V.M.; Silliman, N.; Streisand, J.; Powell, J.; Åkesson, A.; et al. Recombinant human anti–transforming growth factor β1 antibody therapy in systemic sclerosis: A multicenter, randomized, placebo-controlled phase I/II trial of CAT-192. Arthritis Care Res. 2006, 56, 323–333. [Google Scholar] [CrossRef]
- Rice, L.M.; Padilla, C.M.; McLaughlin, S.R.; Mathes, A.L.; Ziemek, J.; Goummih, S.; Nakerakanti, S.; York, M.; Farina, G.; Whitfield, M.L.; et al. Fresolimumab treatment decreases biomarkers and improves clinical symptoms in systemic sclerosis patients. J. Clin. Investig. 2015, 125, 2795–2807. [Google Scholar] [CrossRef] [PubMed]
- King, T.E., Jr.; Bradford, W.Z.; Castro-Bernardini, S.; Fagan, E.A.; Glaspole, I.; Glassberg, M.K.; Gorina, E.; Hopkins, P.M.; Kardatzke, D.; Lancaster, L.; et al. A phase 3 trial of pirfenidone in patients with idiopathic pulmonary fibrosis. N. Engl. J. Med. 2014, 370, 2083–2092. [Google Scholar] [CrossRef]
- Richeldi, L.; Kolb, M.; Jouneau, S.; Wuyts, W.A.; Schinzel, B.; Stowasser, S.; Quaresma, M.; Raghu, G. Efficacy and safety of nintedanib in patients with advanced idiopathic pulmonary fibrosis. BMC Pulm. Med. 2020, 20, 3. [Google Scholar] [CrossRef] [Green Version]
- Richeldi, L.; Du Bois, R.M.; Raghu, G.; Azuma, A.; Brown, K.K.; Costabel, U.; Cottin, V.; Flaherty, K.R.; Hansell, D.M.; Inoue, Y.; et al. Efficacy and safety of nintedanib in idiopathic pulmonary fibrosis. N. Engl. J. Med. 2014, 370, 2071–2082. [Google Scholar] [CrossRef] [Green Version]
- Distler, O.; Highland, K.B.; Gahlemann, M.; Azuma, A.; Fischer, A.; Mayes, M.D.; Raghu, G.; Sauter, W.; Girard, M.; Alves, M.; et al. Nintedanib for Systemic Sclerosis–Associated Interstitial Lung Disease. N. Engl. J. Med. 2019, 380, 2518–2528. [Google Scholar] [CrossRef] [PubMed]
- Bathori, G.; Cervenak, L.; Karadi, I. Caveolae—An alternative endocytotic pathway for targeted drug delivery. Crit. Rev. Ther. Drug Carr. Syst. 2004, 21, 30–96. [Google Scholar] [CrossRef]
- Loh, H.-S.; Green, B.J.; Yusibov, V. Using transgenic plants and modified plant viruses for the development of treatments for human diseases. Curr. Opin. Virol. 2017, 26, 81–89. [Google Scholar] [CrossRef]
- Yao, J.; Weng, Y.; Dickey, A.; Wang, K.Y. Plants as Factories for Human Pharmaceuticals: Applications and Challenges. Int. J. Mol. Sci. 2015, 16, 28549–28565. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.T.; Li, Y.; Wang, S.D.; Han, C.M. Effects of endostatin pretreatment on fibrosis of human skin fibroblasts and the mechanisms. Zhonghua Shao Shang Za Zhi. 2017, 33, 694–698. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Takihara, T.; Chambers, R.A.; Veraldi, K.L.; Larregina, A.T.; Feghali-Bostwick, C.A. A Peptide Derived from Endostatin Ameliorates Organ Fibrosis. Sci. Transl. Med. 2012, 4, 136ra71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Reilly, M.S.; Boehm, T.; Shing, Y.; Fukai, N.; Vasios, G.; Lane, W.S.; Flynn, E.; Birkhead, J.R.; Olsen, B.R.; Folkman, J. Endostatin: An endogenous inhibitor of angiogenesis and tumor growth. Cell 1997, 88, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.-H.; Javier, J.A.; Chang, G.-Y.; Oliveira, H.B.; Azar, D.T. Functional characterization of neostatins, the MMP-derived, enzymatic cleavage products of type XVIII collagen. FEBS Lett. 2005, 579, 3601–3606. [Google Scholar] [CrossRef]
- Ichinose, K.; Maeshima, Y.; Yamamoto, Y.; Kitayama, H.; Takazawa, Y.; Hirokoshi, K.; Sugiyama, H.; Yamasaki, Y.; Eguchi, K.; Makino, H. Antiangiogenic endostatin peptide ameliorates renal alterations in the early stage of a type 1 diabetic nephropathy model. Diabetes 2005, 54, 2891–2903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimoto, T.; Mlakar, L.; Takihara, T.; Feghali-Bostwick, C. An endostatin-derived peptide orally exerts anti-fibrotic activity in a murine pulmonary fibrosis model. Int. Immunopharmacol. 2015, 28, 1102–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assassi, S.; Radstake, T.R.; Mayes, M.D.; Martin, J. Genetics of scleroderma: Implications for personalized medicine? BMC Med. 2013, 11, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef]
- Tjin, G.; White, E.S.; Faiz, A.; Sicard, D.; Tschumperlin, D.J.; Mahar, A.; Kable, E.P.W.; Burgess, J.K. Lysyl oxidases regulate fibrillar collagen remodelling in idiopathic pulmonary fibrosis. Dis. Model. Mech. 2017, 10, 1301–1312. [Google Scholar] [CrossRef] [Green Version]
- Philp, C.J.; Siebeke, I.; Clements, D.; Miller, S.; Habgood, A.; John, A.E.; Navaratnam, V.; Hubbard, R.B.; Jenkins, G.; Johnson, S.R. Extracellular Matrix Cross-Linking Enhances Fibroblast Growth and Protects against Matrix Proteolysis in Lung Fibrosis. Am. J. Respir. Cell Mol. Biol. 2018, 58, 594–603. [Google Scholar] [CrossRef]
- Rimar, D.; Rosner, I.; Nov, Y.; Slobodin, G.; Rozenbaum, M.; Halasz, K.; Haj, T.; Jiries, N.; Kaly, L.; Boulman, N.; et al. Brief report: Lysyl oxidase is a potential biomarker of fibrosis in systemic sclerosis. Arthritis Rheumatol. 2013, 66, 726–730. [Google Scholar] [CrossRef]
- Nguyen, X.-X.M.; Nishimoto, T.; Takihara, T.; Mlakar, L.; Bradshaw, A.D.; Feghali-Bostwick, C.A. Lysyl oxidase directly contributes to extracellular matrix production and fibrosis in systemic sclerosis. Am. J. Physiol. Cell Mol. Physiol. 2021, 320, L29–L40. [Google Scholar] [CrossRef] [PubMed]
- Ashcroft, T.; Simpson, J.M.; Timbrell, V. Simple method of estimating severity of pulmonary fibrosis on a numerical scale. J. Clin. Pathol. 1988, 41, 467–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.G.; Cho, S.J.; Kang, M.J.; Chapoval, S.P.; Lee, P.J.; Noble, P.W.; Yehualaeshet, T.; Lu, B.; Flavell, R.A.; Milbrandt, J.; et al. Early growth response gene 1–mediated apoptosis is essential for transforming growth factor β1–induced pulmonary fibrosis. J. Exp. Med. 2004, 200, 377–389. [Google Scholar] [CrossRef]
- Su, Y.; Nishimoto, T.; Hoffman, S.; Nguyen, X.-X.; Pilewski, J.M.; Feghali-Bostwick, C. Insulin-like growth factor binding protein-4 exerts antifibrotic activity by reducing levels of connective tissue growth factor and the C-X-C chemokine receptor 4. FASEB BioAdv. 2019, 1, 167–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, X.-X.; Muhammad, L.; Nietert, P.J.; Feghali-Bostwick, C. IGFBP-5 Promotes Fibrosis via Increasing Its Own Expression and That of Other Pro-fibrotic Mediators. Front. Endocrinol. 2018, 9, 00601. [Google Scholar] [CrossRef] [PubMed]
- Morais, A.; Beltrão, M.; Sokhatska, O.; Costa, D.; Melo, N.; Mota, P.; Marques, A.; Delgado, L. Serum metalloproteinases 1 and 7 in the diagnosis of idiopathic pulmonary fibrosis and other interstitial pneumonias. Respir. Med. 2015, 109, 1063–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahalanobish, S.; Saha, S.; Dutta, S.; Sil, P.C.; Mahalanobish, S.; Saha, S.; Dutta, S.; Sil, P.C. Matrix metalloproteinase: An upcoming therapeutic approach for idiopathic pulmonary fibrosis. Pharmacol. Res. 2020, 152, 104591. [Google Scholar] [CrossRef]
- Popov, Y.V.; Sverdlov, D.Y.; Bhaskar, K.R.; Sharma, A.K.; Millonig, G.; Patsenker, E.; Krahenbuhl, S.; Krahenbuhl, L.; Schuppan, D. Macrophage-mediated phagocytosis of apoptotic cholangiocytes contributes to reversal of experimental biliary fibrosis. Am. J. Physiol. Liver Physiol. 2010, 298, G323–G334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirastschijski, U.; Lupše, B.; Maedler, K.; Sarma, B.; Radtke, A.; Belge, G.; Dorsch, M.; Wedekind, D.; McCawley, L.J.; Boehm, G.; et al. Matrix Metalloproteinase-3 is Key Effector of TNF-α-Induced Collagen Degradation in Skin. Int. J. Mol. Sci. 2019, 20, 5234. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Watanabe, T.; Nishimoto, T.; Takihara, T.; Mlakar, L.; Nguyen, X.-X.; Sanderson, M.; Su, Y.; Chambers, R.A.; Feghali-Bostwick, C. E4 engages uPAR and enolase-1 and activates urokinase to exert antifibrotic effects. JCI Insight 2021, 6, 144935. [Google Scholar] [CrossRef]
- Sumi, M.; Satoh, H.; Kagohashi, K.; Ishikawa, H.; Sekizawa, K. Increased serum levels of endostatin in patients with idiopathic pulmonary fibrosis. J. Clin. Lab. Anal. 2005, 19, 146–149. [Google Scholar] [CrossRef]
- Hummers, L.K.; Hall, A.; Wigley, F.M.; Simons, M. Abnormalities in the regulators of angiogenesis in patients with scleroderma. J. Rheumatol. 2009, 36, 576–582. [Google Scholar] [CrossRef]
- Reiseter, S.; Molberg, Ø.; Gunnarsson, R.; Lund, M.B.; Aalokken, T.M.; Aukrust, P.; Ueland, T.; Garen, T.; Brunborg, C.; Michelsen, A.; et al. Associations between circulating endostatin levels and vascular organ damage in systemic sclerosis and mixed connective tissue disease: An observational study. Arthritis Res. Ther. 2015, 17, 231. [Google Scholar] [CrossRef] [PubMed]
- Dziankowska-Bartkowiak, B.; Waszczykowska, E.; Zalewska, A.; Sysa-Jędrzejowska, A. Correlation of endostatin and tissue inhibitor of metalloproteinases 2 (TIMP2) serum levels with cardiovascular involvement in systemic sclerosis patients. Mediat. Inflamm. 2005, 2005, 144–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldman, A.; Tamarkin, L.; Paciotti, G.F.; Simpson, B.W.; Linehan, W.M.; Yang, J.C.; Fogler, W.E.; Turner, E.M.; Alexander, H.R.; Libutti, S.K. Serum endostatin levels are elevated and correlate with serum vascular endothelial growth factor levels in patients with stage IV clear cell renal cancer. Clin. Cancer Res. 2000, 6, 4628–4634. [Google Scholar] [PubMed]
- Bono, P.; Teerenhovi, L.; Joensuu, H. Elevated serum endostatin is associated with poor outcome in patients with non-Hodgkin lymphoma. Cancer 2003, 97, 2767–2775. [Google Scholar] [CrossRef] [PubMed]
- Feldman, A.; Pak, H.; Yang, J.C.; Alexander, H.R.; Libutti, S.K. Serum endostatin levels are elevated in patients with soft tissue sarcoma. Cancer 2001, 91, 1525–1529. [Google Scholar] [CrossRef]
- Yao, J.; Fan, L.; Peng, C.; Huang, A.; Liu, T.; Lin, Z.; Yang, Q.; Zhang, T.; Ma, H. Clinical efficacy of endostar combined with chemotherapy in the treatment of peritoneal carcinomatosis in gastric cancer: Results from a retrospective study. Oncotarget 2017, 8, 70788–70797. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Lv, D.; Meng, Y.; Wang, M.; Wang, W.; Zhou, C.; Zhou, S.; Chen, X.; Yang, H. Endostar improved efficacy of concurrent chemoradiotherapy with vinorelbine plus carboplatin in locally advanced lung squamous cell carcinoma patients with high serum Lp(a) concentration. Ann. Palliat. Med. 2020, 9, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Honglian, M.; Zhouguang, H.; Fang, P.; Lujun, Z.; Dongming, L.; Yujin, X.; Yong, B.; Liming, X.; YiRui, Z.; Xiao, H.; et al. Different administration routes of recombinant human endostatin combined with concurrent chemoradiotherapy might lead to different efficacy and safety profile in unresectable stage III non-small cell lung cancer: Updated follow-up results from two phase II trials. Thorac. Cancer 2020, 11, 898–906. [Google Scholar] [CrossRef]
- Li, Y.; Tian, Y.; Jin, F.; Wu, W.; Long, J.; Ouyang, J.; Zhou, Y. A phase II multicenter randomized controlled trial to compare standard chemoradiation with or without recombinant human endostatin injection (Endostar) therapy for the treatment of locally advanced nasopharyngeal carcinoma: Long-term outcomes update. Curr. Probl. Cancer 2019, 44, 100492. [Google Scholar] [CrossRef]
- Szentirmai, O.; Baker, C.H.; Bullain, S.S.; Lin, N.; Takahashi, M.; Folkman, J.; Mulligan, R.C.; Carter, B.S. Successful inhibition of intracranial human glioblastoma multiforme xenograft growth via systemic adenoviral delivery of soluble endostatin and soluble vascular endothelial growth factor receptor-2. J. Neurosurg. 2008, 108, 979–988. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Ding, M.; Qiao, Y.; Liu, Y.; Liu, L. Recombinant human endostatin combined with radiotherapy in the treatment of brain metastases of non-small cell lung cancer. Clin. Transl. Oncol. 2013, 16, 630–636. [Google Scholar] [CrossRef]
- Wan, Y.-Y.; Tian, G.-Y.; Guo, H.-S.; Kang, Y.-M.; Yao, Z.-H.; Li, X.-L.; Liu, Q.-H.; Lin, D.-J. Endostatin, an angiogenesis inhibitor, ameliorates bleomycin-induced pulmonary fibrosis in rats. Respir. Res. 2013, 14, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Liu, D.-G.; Yang, G.; Kong, L.-J.; Du, Y.-J.; Wang, H.-Y.; Li, F.-D.; Pei, F.-H.; Song, J.-T.; Fan, Y.-J.; et al. Endostar, a novel human recombinant endostatin, attenuates liver fibrosis in CCl4-induced mice. Exp. Biol. Med. 2014, 239, 998–1006. [Google Scholar] [CrossRef] [PubMed]
- Zhiyong, W.; Fei, S.; Lianju, X.; Yingkai, L.; Chun, Q.; Shuliang, L.; Xiqiao, W. Endostar injection inhibits rabbit ear hypertrophic scar formation. Int. J. Low. Extrem. Wounds 2012, 11, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.-T.; Hu, H.; Li, Y.; Jiang, H.-F.; Hu, X.-L.; Han, C.-M. Endostatin inhibits hypertrophic scarring in a rabbit ear model. J. Zhejiang Univ. Sci. B 2013, 14, 224–230. [Google Scholar] [CrossRef]
- Bai, X.; Li, X.; Tian, J.; Zhou, Z. Antiangiogenic treatment diminishes renal injury and dysfunction via regulation of local AKT in early experimental diabetes. PLoS ONE 2014, 9, e96117. [Google Scholar] [CrossRef] [Green Version]
- Baig, M.H.; Ahmad, K.; Saeed, M.; Alharbi, A.M.; Barreto, G.E.; Ashraf, G.M.; Choi, I. Peptide based therapeutics and their use for the treatment of neurodegenerative and other diseases. Biomed. Pharmacother. 2018, 103, 574–581. [Google Scholar] [CrossRef]
- Danho, W.; Swistok, J.; Khan, W.; Chu, X.-J.; Cheung, A.; Fry, D.; Sun, H.; Kurylko, G.; Rumennik, L.; Cefalu, J.; et al. Opportunities and challenges of developing peptide drugs in the pharmaceutical industry. Adv. Exp. Med. Biol. 2009, 611, 467–469. [Google Scholar] [CrossRef]
- Lau, J.L.; Dunn, M.K. Therapeutic peptides: Historical perspectives, current development trends, and future directions. Bioorg. Med. Chem. 2018, 26, 2700–2707. [Google Scholar] [CrossRef]
- Knight, P.D.; Karamanos, T.; Radford, S.; Ashcroft, A.E. Identification of a novel site of interaction between ataxin-3 and the amyloid aggregation inhibitor polyglutamine binding peptide 1. Eur. J. Mass Spectrom. 2017, 24, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Sragovich, S.; Amram, N.; Yeheskel, A.; Gozes, I. VIP/PACAP-Based Drug Development: The ADNP/NAP-Derived Mirror Peptides SKIP and D-SKIP Exhibit Distinctive in vivo and in silico Effects. Front. Cell Neurosci. 2020, 13, 589. [Google Scholar] [CrossRef]
- Igarashi, H.; Ito, T.; Mantey, S.A.; Pradhan, T.K.; Hou, W.; Coy, D.H.; Jensen, R.T. Development of simplified vasoactive intestinal peptide analogs with receptor selectivity and stability for human vasoactive intestinal peptide/pituitary adenylate Cyclase-activating polypeptide receptors. J. Pharmacol. Exp. Ther. 2005, 315, 370–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, N.; Ferreira, D.; Rodrigues, M.C.; Cintra, A.; Santos, N.; Sampaio, S.; Santos, A. Low-molecular-mass peptides from the venom of the Amazonian viper Bothrops atrox protect against brain mitochondrial swelling in rat: Potential for neuroprotection. Toxicon 2010, 56, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, B.H.; Fox, A.W.; Stewart, A.J.; Gold, M. Davunetide: A review of safety and efficacy data with a focus on neurodegenerative diseases. Expert Rev. Clin. Pharmacol. 2013, 6, 483–502. [Google Scholar] [CrossRef] [PubMed]
- Disatnik, M.; Ferreira, J.C.; Campos, J.C.; Gomes, K.S.; Dourado, P.M.; Qi, X.; Mochly-Rosen, D. Acute inhibition of excessive mitochondrial fission after myocardial infarction prevents long-term cardiac dysfunction. J. Am. Heart Assoc. 2013, 2, e000461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filichia, E.; Hoffer, B.; Qi, X.; Luo, Y. Inhibition of Drp1 mitochondrial translocation provides neural protection in dopaminergic system in a Parkinson’s disease model induced by MPTP. Sci. Rep. 2016, 6, 32656. [Google Scholar] [CrossRef] [Green Version]
- Barish, C.; Dorn, S.; Fogel, R.P.; Patel, R.; Rosenberg, J. Plecanatide Is Effective and Safe in the Treatment for Chronic Idiopathic Constipation: Results of a Phase II Trial. Am. J. Dig. Dis. 2020, 66, 537–540. [Google Scholar] [CrossRef]
- Engel, J.B.; Tinneberg, H.-R.; Rick, F.G.; Berkes, E.; Schally, A.V. Targeting of Peptide Cytotoxins to LHRH Receptors for Treatment of Cancer. Curr. Drug Targets 2016, 17, 488–494. [Google Scholar] [CrossRef]
- Enokida, H.; Yamada, Y.; Tatarano, S.; Yoshino, H.; Yonemori, M.; Sakaguchi, T.; Nishimura, H.; Eura, R.; Nakagawa, M. Oncological outcome of neoadjuvant low-dose estramustine plus LHRH agonist/antagonist followed by extended radical prostatectomy for Japanese patients with high-risk localized prostate cancer: A prospective single-arm study. Jpn. J. Clin. Oncol. 2019, 50, 66–72. [Google Scholar] [CrossRef]
- Schally, A.V.; Wang, H.; He, J.; Cai, R.; Sha, W.; Popovics, P.; Perez, R.; Vidaurre, I.; Zhang, X. Agonists of growth hormone-releasing hormone (GHRH) inhibit human experimental cancers in vivo by down-regulating receptors for GHRH. Proc. Natl. Acad. Sci. USA 2018, 115, 12028–12033. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.X.; Cheng, A.S.; To, K.F.; Tong, J.H.; Li, M.S.; Shen, J.; Wong, C.C.; Zhang, L.; Chan, R.L.; Wang, X.J.; et al. Host immune defense peptide LL-37 activates caspase-independent apoptosis and suppresses colon cancer. Cancer Res. 2012, 72, 6512–6523. [Google Scholar] [CrossRef]
- Valentinis, B.; Porcellini, S.; Asperti, C.; Cota, M.; Zhou, D.; Di Matteo, P.; Garau, G.; Zucchelli, C.; Avanzi, N.R.; Rizzardi, G.P.; et al. Mechanism of Action of the Tumor Vessel Targeting Agent NGR-hTNF: Role of Both NGR Peptide and hTNF in Cell Binding and Signaling. Int. J. Mol. Sci. 2019, 20, 4511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Wu, W.K.K.; Gallo, R.L.; Fang, E.F.; Hu, W.; Ling, T.K.W.; Shen, J.; Chan, R.L.Y.; Lu, L.; Luo, X.M.; et al. Critical Role of Antimicrobial Peptide Cathelicidin for Controlling Helicobacter pylori Survival and Infection. J. Immunol. 2016, 196, 1799–1809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.-M.; Lin, L.-C.; Wang, C.-F.; Lee, Y.-J.; Chen, Y.-T.; Liao, Y.-D. Antimicrobial Properties of an Immunomodulator—15 kDa Human Granulysin. PLoS ONE 2016, 11, e0156321. [Google Scholar] [CrossRef] [Green Version]
- Akram, A.R.; Avlonitis, N.; Scholefield, E.; Vendrell, M.; McDonald, N.; Aslam, T.; Craven, T.H.; Gray, C.; Collie, D.S.; Fisher, A.J.; et al. Enhanced avidity from a multivalent fluorescent antimicrobial peptide enables pathogen detection in a human lung model. Sci. Rep. 2019, 9, 8422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, A.C.; Janson, H.; Wold, H.; Fugelli, A.; Andersson, K.; Håkangård, C.; Olsson, P.; Olsen, W.M. LTX-109 is a novel agent for nasal decolonization of methicillin-resistant and -sensitive Staphylococcus aureus. Antimicrob. Agents Chemother. 2015, 59, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Manandhar, B.; Ahn, J.-M. Glucagon-like peptide-1 (GLP-1) analogs: Recent advances, new possibilities, and therapeutic implications. J. Med. Chem. 2014, 58, 1020–1037. [Google Scholar] [CrossRef] [Green Version]
- Morieri, M.L.; Frison, V.; Rigato, M.; D’Ambrosio, M.; Tadiotto, F.; Paccagnella, A.; Simioni, N.; Lapolla, A.; Avogaro, A.; Fadini, G.P. Effectiveness of Dulaglutide in the Real World and in Special Populations of Type 2 Diabetic Patients. J. Clin. Endocrinol. Metab. 2020, 105, e2617–e2625. [Google Scholar] [CrossRef]
- Pozzilli, P.; Bosi, E.; Cirkel, D.; Harris, J.; Leech, N.; Tinahones, F.J.; Vantyghem, M.-C.; Vlasakakis, G.; Ziegler, A.-G.; Janmohamed, S. Randomized 52-week Phase 2 Trial of Albiglutide Versus Placebo in Adult Patients with Newly Diagnosed Type 1 Diabetes. J. Clin. Endocrinol. Metab. 2020, 105, e2192–e2206. [Google Scholar] [CrossRef]
- Drucker, D.J. Advances in oral peptide therapeutics. Nat. Rev. Drug Discov. 2019, 19, 277–289. [Google Scholar] [CrossRef]
- Shaji, J.; Patole, V. Protein and peptide drug delivery: Oral approaches. Indian J. Pharm. Sci. 2008, 70, 269–277. [Google Scholar] [CrossRef]
- Boehm, T.; Folkman, J.; Browder, T.; O’Reilly, M.S. Antiangiogenic therapy of experimental cancer does not induce acquired drug resistance. Nature 1997, 390, 404–407. [Google Scholar] [CrossRef] [PubMed]
- Musiychuk, K.; Stephenson, N.; Bi, H.; Farrance, C.E.; Orozovic, G.; Brodelius, M.; Brodelius, P.; Horsey, A.; Ugulava, N.; Shamloul, A.; et al. A launch vector for the production of vaccine antigens in plants. Influenza Other Respir. Viruses 2007, 1, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Nishimoto, T.; Mlakar, L.; Heywood, J.; Malaab, M.; Hoffman, S.; Feghali-Bostwick, C. Optimization of a murine and human tissue model to recapitulate dermal and pulmonary features of systemic sclerosis. PLoS ONE 2017, 12, e0179917. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mlakar, L.; Garrett, S.M.; Watanabe, T.; Sanderson, M.; Nishimoto, T.; Heywood, J.; Helke, K.L.; Pilewski, J.M.; Herzog, E.L.; Feghali-Bostwick, C. Ameliorating Fibrosis in Murine and Human Tissues with END55, an Endostatin-Derived Fusion Protein Made in Plants. Biomedicines 2022, 10, 2861. https://doi.org/10.3390/biomedicines10112861
Mlakar L, Garrett SM, Watanabe T, Sanderson M, Nishimoto T, Heywood J, Helke KL, Pilewski JM, Herzog EL, Feghali-Bostwick C. Ameliorating Fibrosis in Murine and Human Tissues with END55, an Endostatin-Derived Fusion Protein Made in Plants. Biomedicines. 2022; 10(11):2861. https://doi.org/10.3390/biomedicines10112861
Chicago/Turabian StyleMlakar, Logan, Sara M. Garrett, Tomoya Watanabe, Matthew Sanderson, Tetsuya Nishimoto, Jonathan Heywood, Kristi L. Helke, Joseph M. Pilewski, Erica L. Herzog, and Carol Feghali-Bostwick. 2022. "Ameliorating Fibrosis in Murine and Human Tissues with END55, an Endostatin-Derived Fusion Protein Made in Plants" Biomedicines 10, no. 11: 2861. https://doi.org/10.3390/biomedicines10112861