
Plasma Metabolomic Profiling Reveals Four Possibly Disrupted Mechanisms in Systemic Sclerosis

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Group and Sample Collection
2.2. Sample Preparation
2.3. HPLC-MS Analysis
2.4. Data Pre-Processing
2.5. Statistical Analysis
3. Results
3.1. Study Group and Sample Analysis
3.2. Targeted Metabolomics
3.3. Untargeted Metabolomics
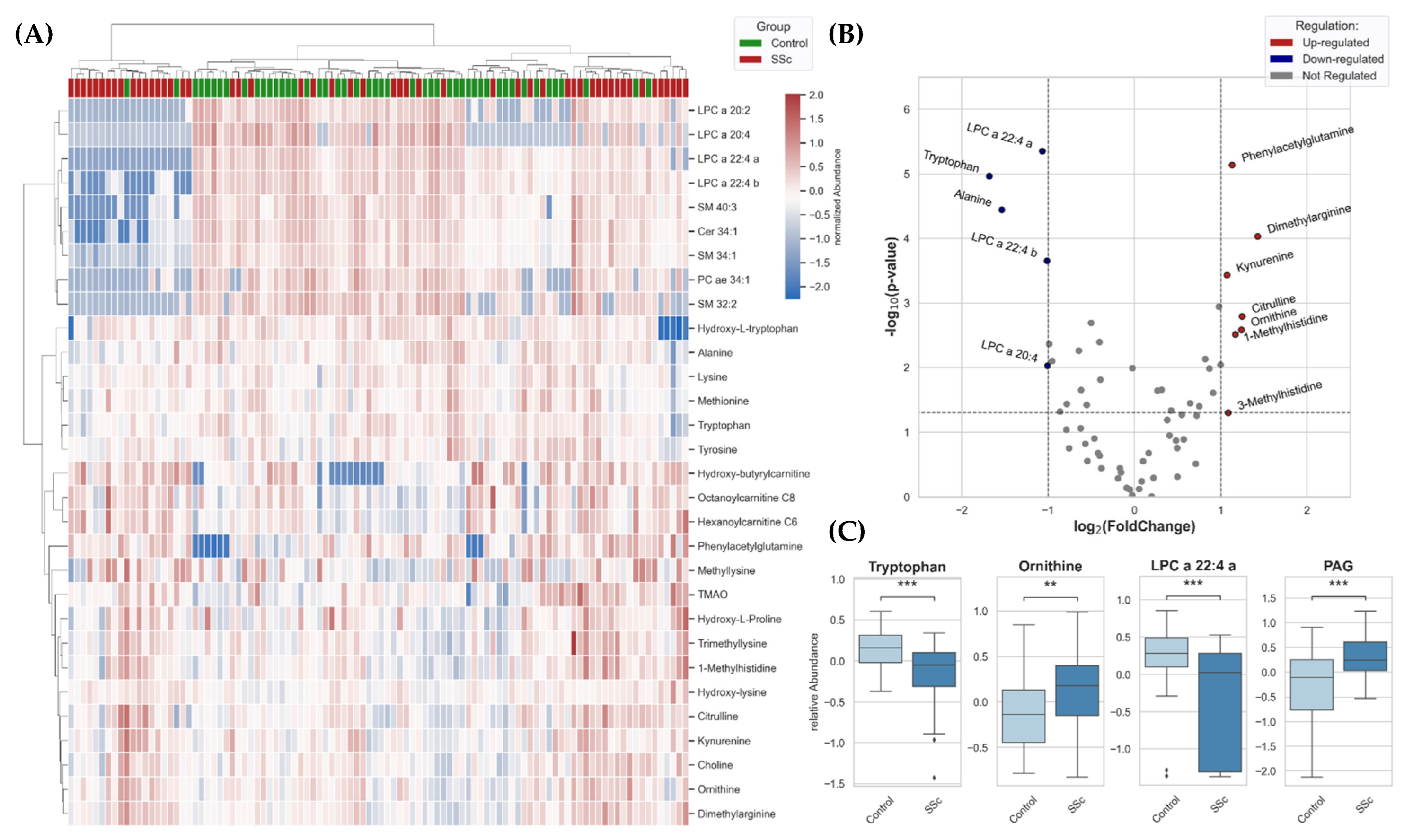
3.4. Metabolomic Patterns Altered in SSc Patients
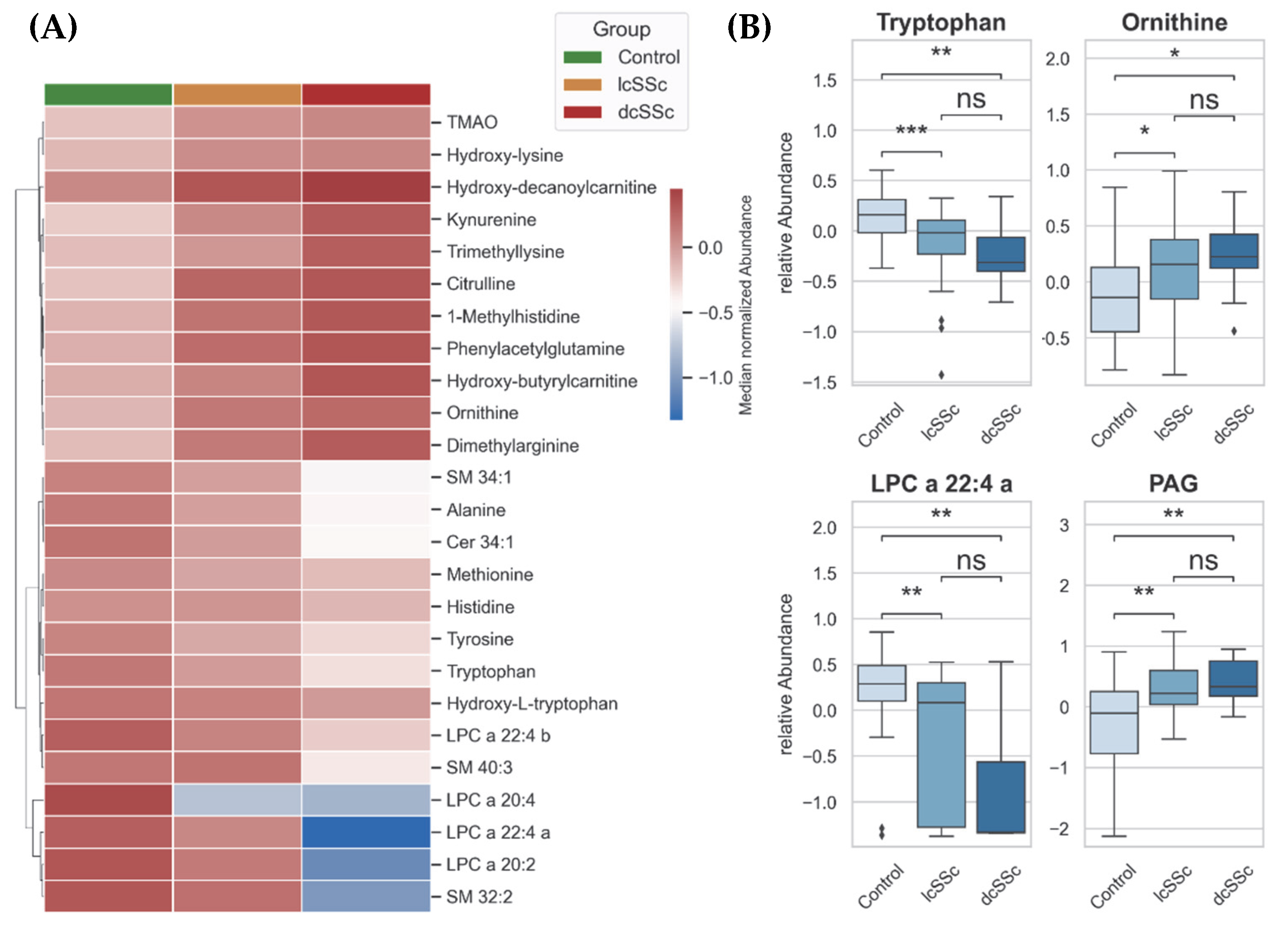
3.5. Altered Metabolism in lcSSc and dcSSc Patients
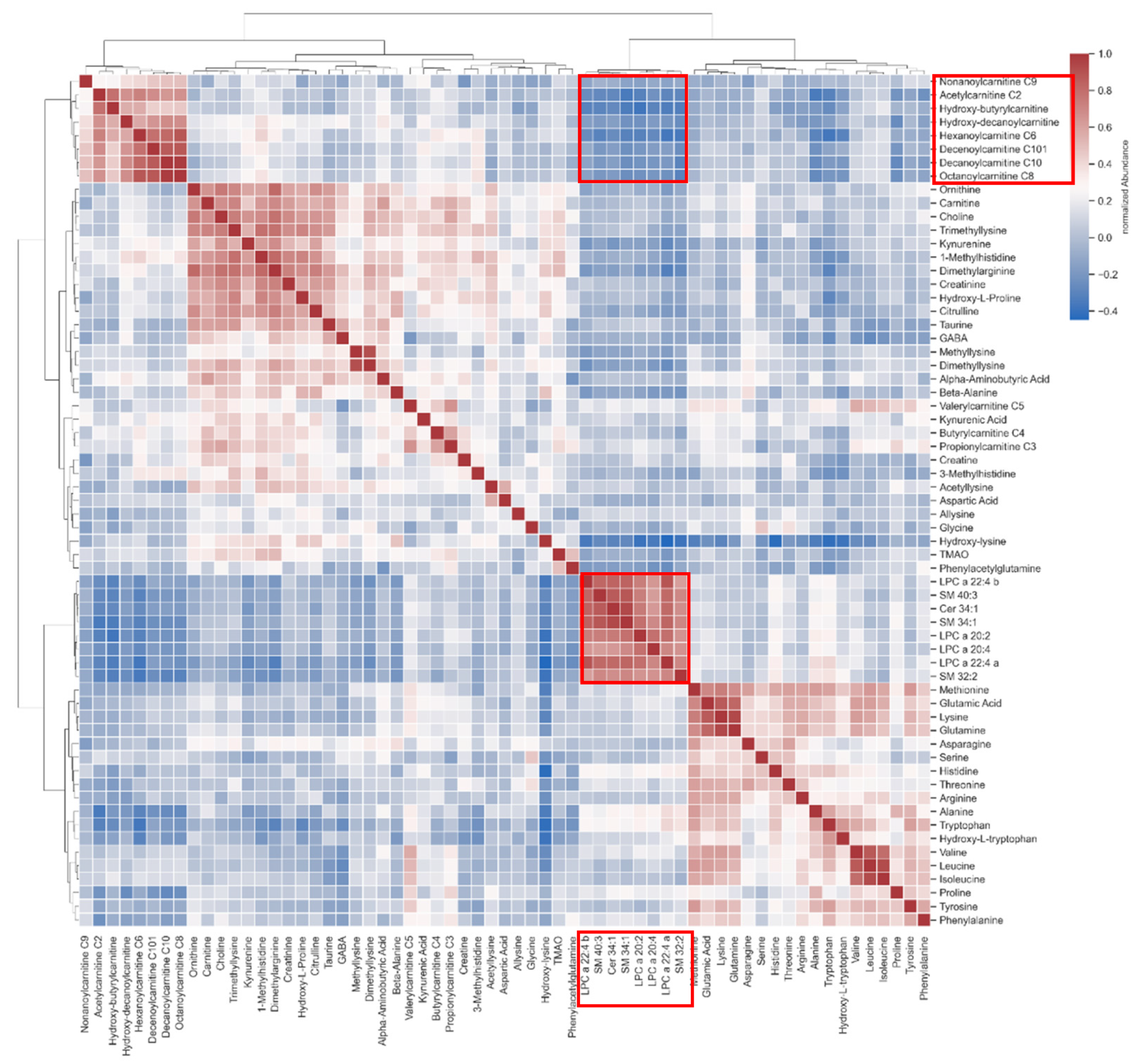
3.6. Cross-Correlation between Lipids and Carnitines
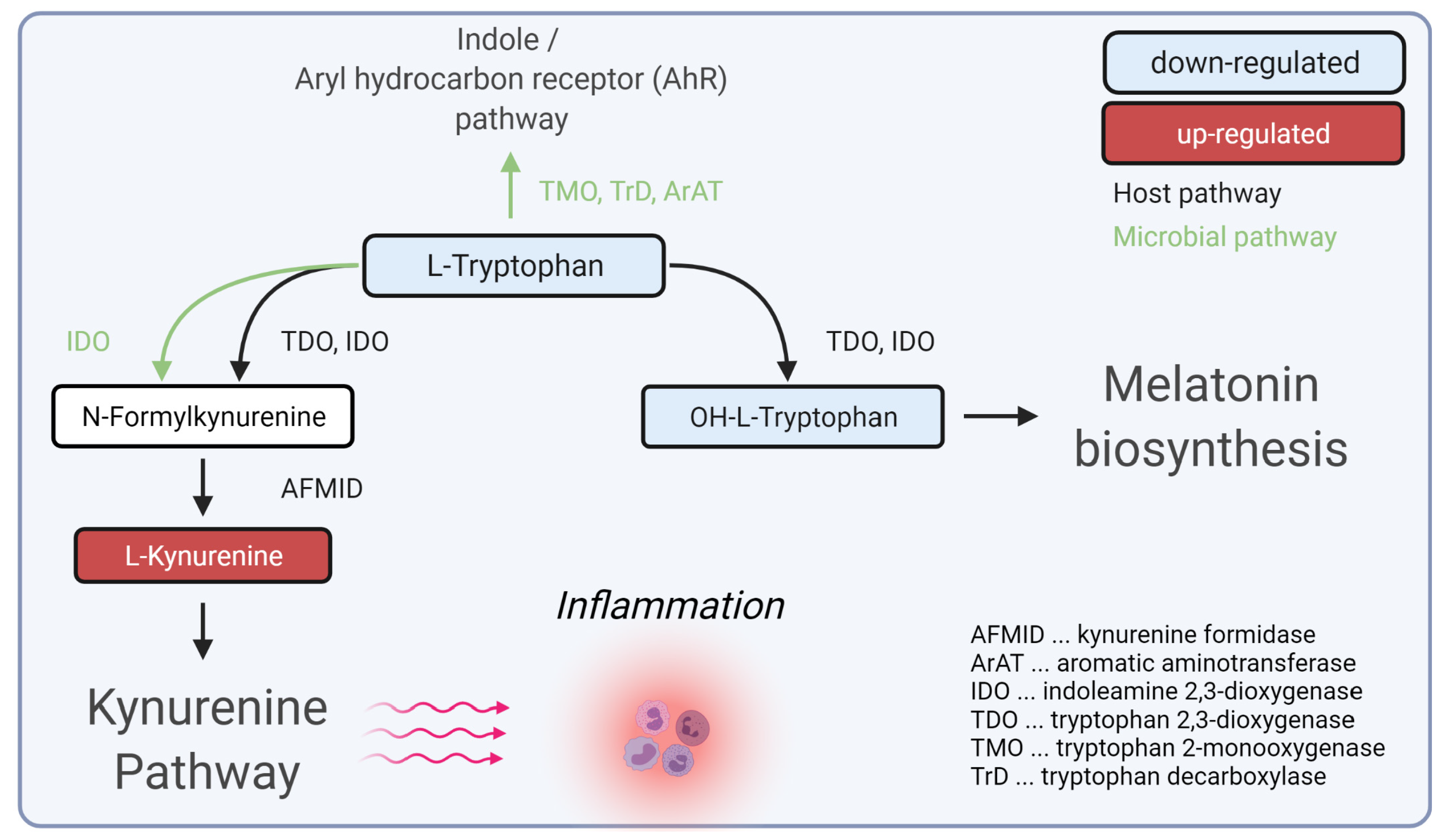
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACR | American College of Rheumatology |
| ADMA | asymmetric dimethylarginine |
| AFMID | kynurenine formidase |
| AhR | Aryl hydrocarbon receptor |
| ANOVA | Analysis of variance |
| ArAT | aromatic aminotransferase |
| ARG | arginase |
| ASL | argininosuccinate lyase |
| ASS | argininosuccinate synthase |
| CCS | collision cross section |
| Cer | ceramide |
| dcSSc | diffuse cutaneous Systemic sclerosis |
| DMA | dimethylarginine |
| FAO | fatty acid oxidation |
| HILIC | hydrophilic interaction chromatography |
| HMDB | Human Metabolome Database |
| HPLC | high-performance liquid chromatography |
| IDO | indoleamine 2,3-dioxygenase |
| IMS-QTOF-MS | ion mobility quadrupole time-of-flight mass spectrometer |
| KP | kynurenine pathway |
| lcSSc | limited cutaneous Systemic sclerosis |
| LDL | low-density lipoproteins |
| LPC | lysophosphatidylcholine |
| m/z | mass to charge ratio |
| MoNA | MassBank of North America |
| mRSS | modified Rodnan Skin Score |
| ncSSc | non-cutaneous systemic sclerosis |
| NOS | nitric oxide synthase |
| NOS | NO-synthase |
| OAT | ornithine aminotransferase |
| ODC | ornithine decarboxylase |
| ORNT1 | ornithine translocase |
| OTC | ornithine transcarbamylase |
| P5C | pyrroline-5-carboxylate |
| PAG | phenylacetylglutamine |
| QQQ-MS | triple quadrupole mass spectrometer |
| RT | retention time |
| SDMA | symmetric dimethylarginine |
| SM | sphingomyelin |
| SSc | systemic sclerosis |
| TDO | tryptophan 2,3-dioxygenase |
| TMO | tryptophan 2-monooxygenase |
| TrD | tryptophan decarboxylase |
References
- Varga, J.; Abraham, D. Systemic sclerosis: A prototypic multisystem fibrotic disorder. J. Clin. Invest. 2007, 117, 557–567. [Google Scholar] [CrossRef]
- Allanore, Y.; Simms, R.; Distler, O.; Trojanowska, M.; Pope, J.; Denton, C.P.; Varga, J. Systemic sclerosis. Nat. Rev. Dis. Primers 2015, 1, 1–21. [Google Scholar] [CrossRef]
- Denton, C.P.; Khanna, D. Systemic sclerosis. Lancet 2017, 390, 1685–1699. [Google Scholar] [CrossRef]
- Van Den Hoogen, F.; Khanna, D.; Fransen, J.; Johnson, S.R.; Baron, M.; Tyndall, A.; Matucci-Cerinic, M.; Naden, R.P.; Medsger, T.A.; Carreira, P.E.; et al. 2013 classification criteria for systemic sclerosis: An American College of Rheumatology/European League against Rheumatism collaborative initiative. Arthritis. Rheum. 2013, 65, 2737–2747. [Google Scholar] [CrossRef] [Green Version]
- Gabrielli, A.; Avvedimento, E.V.; Krieg, T. Scleroderma. N. Engl. J. Med. 2009, 360, 1989–2003. [Google Scholar] [CrossRef]
- Khanna, D.; Furst, D.E.; Clements, P.J.; Allanore, Y.; Baron, M.; Czirjak, L.; Distler, O.; Foeldvari, I.; Kuwana, M.; Matucci-Cerinic, M.; et al. Standardization of the modified Rodnan skin score for use in clinical trials of systemic sclerosis. Scleroderma Relat. Disord. 2017, 2, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Hinchcliff, M.; O’Reilly, S. Current and Potential New Targets in Systemic Sclerosis Therapy: A New Hope. Curr. Rheumatol. Rep. 2020, 22, 1–7. [Google Scholar] [CrossRef]
- Misra, D.P.; Ahmed, S.; Agarwal, V. Is biological therapy in systemic sclerosis the answer? Rheumatol Int. 2020, 40, 679–694. [Google Scholar] [CrossRef]
- Sierra-Sepúlveda, A.; Esquinca-González, A.; Benavides-Suárez, S.A.; Sordo-Lima, D.E.; Caballero-Islas, A.E.; Cabral-Castañeda, A.R.; Rodríguez-Reyna, T.S. Systemic Sclerosis Pathogenesis and Emerging Therapies, beyond the Fibroblast. Biomed. Res. Int. 2019, 2019, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zanatta, E.; Codullo, V.; Avouac, J.; Allanore, Y. Systemic sclerosis: Recent insight in clinical management. Jt. Bone Spine 2020, 87, 293–299. [Google Scholar] [CrossRef]
- Distler, O.; Highland, K.B.; Gahlemann, M.; Azuma, A.; Fischer, A.; Mayes, M.D.; Raghu, G.; Sauter, W.; Girard, M.; Alves, M.; et al. Nintedanib for Systemic Sclerosis-Associated Interstitial Lung Disease. N. Engl. J. Med. 2019, 380, 2518–2528. [Google Scholar] [CrossRef] [PubMed]
- Yoon, N.; Jang, A.-K.; Seo, Y.; Jung, B.H. Metabolomics in Autoimmune Diseases: Focus on Rheumatoid Arthritis, Systemic Lupus Erythematous, and Multiple Sclerosis. Metabolites 2021, 11, 812. [Google Scholar] [CrossRef] [PubMed]
- Sandlers, Y. Amino Acids Profiling for the Diagnosis of Metabolic Disorders. Biochem. Test. 2019, 12, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S. Metabolomics for Investigating Physiological and Pathophysiological Processes. Physiol. Rev. 2019, 99, 1819–1875. [Google Scholar] [CrossRef] [PubMed]
- Murgia, F.; Svegliati, S.; Poddighe, S.; Lussu, M.; Manzin, A.; Spadoni, T.; Fischetti, C.; Gabrielli, A.; Atzori, L. Metabolomic profile of systemic sclerosis patients. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Bellocchi, C.; Fernández-Ochoa, Á.; Montanelli, G.; Vigone, B.; Santaniello, A.; Milani, C.; Quirantes-Piné, R.; Borrás-Linares, I.; Ventura, M.; Segura-Carrettero, A.; et al. Microbial and metabolic multi-omic correlations in systemic sclerosis patients. Ann. N. Y. Acad. Sci. 2018, 1421, 97–109. [Google Scholar] [CrossRef]
- Fernández-Ochoa, Á.; Quirantes-Piné, R.; Borrás-Linares, I.; Gemperline, D.; Alarcón Riquelme, M.E.; Beretta, L.; Segura-Carretero, A. Urinary and plasma metabolite differences detected by HPLC-ESI-QTOF-MS in systemic sclerosis patients. J. Pharm. Biomed. Anal. 2019, 162, 82–90. [Google Scholar] [CrossRef]
- Ottria, A.; Hoekstra, A.T.; Zimmermann, M.; Van Der Kroef, M.; Vazirpanah, N.; Cossu, M.; Chouri, E.; Rossato, M.; Beretta, L.; Tieland, R.G.; et al. Fatty Acid and Carnitine Metabolism Are Dysregulated in Systemic Sclerosis Patients. Front. Immunol. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Smolenska, Z.; Zabielska-Kaczorowska, M.; Wojteczek, A.; Kutryb-Zajac, B.; Zdrojewski, Z. Metabolic Pattern of Systemic Sclerosis: Association of Changes in Plasma Concentrations of Amino Acid-Related Compounds With Disease Presentation. Front. Mol. Biosci. 2020, 7, 1–7. [Google Scholar] [CrossRef]
- Deidda, M.; Piras, C.; Cadeddu Dessalvi, C.; Locci, E.; Barberini, L.; Orofino, S.; Musu, M.; Mura, M.N.; Manconi, P.E.; Finco, G.; et al. Distinctive metabolomic fingerprint in scleroderma patients with pulmonary arterial hypertension. Int. J. Cardiol. 2017, 241, 401–406. [Google Scholar] [CrossRef]
- Meier, C.; Freiburghaus, K.; Bovet, C.; Schniering, J.; Allanore, Y.; Distler, O.; Nakas, C.; Maurer, B. Serum metabolites as biomarkers in systemic sclerosis-associated interstitial lung disease. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Luo, M.; Chen, X.; Yin, Y.; Xiong, X.; Wang, R.; Zhu, Z.-J. Ion mobility collision cross-section atlas for known and unknown metabolite annotation in untargeted metabolomics. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, 1–128. [Google Scholar] [CrossRef]
- Shi, B.; Wang, W.; Korman, B.; Kai, L.; Wang, Q.; Wei, J.; Bale, S.; Marangoni, R.G.; Bhattacharyya, S.; Miller, S.; et al. Targeting CD38-dependent NAD+ metabolism to mitigate multiple organ fibrosis. iScience 2021, 24, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, D.; Song, P.; Zou, M.H. Tryptophan-kynurenine pathway is dysregulated in inflammation, and immune activation. Front. Biosci. 2015, 1116–1143. [Google Scholar]
- Mellor, A.L.; Munn, D.H. IDO expression by dendritic cells: Tolerance and tryptophan catabolism. Nat. Rev. Immunol. 2004, 4, 762–774. [Google Scholar] [CrossRef] [PubMed]
- Mándi, Y.; Vécsei, L. The kynurenine system and immunoregulation. J. Neural. Transm. 2012, 119, 197–209. [Google Scholar] [CrossRef]
- Fallarino, F.; Grohmann, U.; Vacca, C.; Bianchi, R.; Orabona, C.; Spreca, A.; Fioretti, M.C.; Puccetti, P. T cell apoptosis by tryptophan catabolism. Cell. Death. Differ. 2002, 9, 1069–1077. [Google Scholar] [CrossRef]
- Puccetti, P.; Grohmann, U. IDO and regulatory T cells: A role for reverse signalling and non-canonical NF-κB activation. Nat. Rev. Immunol. 2007, 7, 817–823. [Google Scholar] [CrossRef]
- Biernacki, T.; Sandi, D.; Bencsik, K.; Vécsei, L. Kynurenines in the Pathogenesis of Multiple Sclerosis: Therapeutic Perspectives. Cells 2020, 9, 1564. [Google Scholar] [CrossRef]
- Sgonc, R.; Gruschwitz, M.S.; Boeck, G.; Sepp, N.; Gruber, J.; Wick, G. Endothelial cell apoptosis in systemic sclerosis is induced by antibody-dependent cell-mediated cytotoxicity via CD95. Arthritis. Rheum. 2000, 43, 2550–2562. [Google Scholar] [CrossRef]
- Agus, A.; Planchais, J.; Sokol, H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell. Host. Microbe. 2018, 23, 716–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patrone, V.; Puglisi, E.; Cardinali, M.; Schnitzler, T.S.; Svegliati, S.; Festa, A.; Gabrielli, A.; Morelli, L. Gut microbiota profile in systemic sclerosis patients with and without clinical evidence of gastrointestinal involvement. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Andréasson, K.; Alrawi, Z.; Persson, A.; Jönsson, G.; Marsal, J. Intestinal dysbiosis is common in systemic sclerosis and associated with gastrointestinal and extraintestinal features of disease. Arthritis. Res. Ther. 2016, 18, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Caldwell, R.W.; Rodriguez, P.C.; Toque, H.A.; Narayanan, S.P.; Caldwell, R.B. Arginase: A Multifaceted Enzyme Important in Health and Disease. Physiol. Rev. 2018, 98, 641–665. [Google Scholar] [CrossRef] [Green Version]
- Watford, M. The Urea Cycle: Teaching intermediary metabolism in a physiological setting. Biochem. Mol. Biol. Educ. 2003, 31, 289–297. [Google Scholar] [CrossRef]
- Bruch-Gerharz, D.; Schnorr, O.; Suschek, C.; Beck, K.-F.; Pfeilschifter, J.; Ruzicka, T.; Kolb-Bachofen, V. Arginase 1 Overexpression in Psoriasis. Am. J. Clin. Pathol. 2003, 162, 203–211. [Google Scholar] [CrossRef]
- Brooks, W.H. Autoimmune diseases and polyamines. Clin. Rev. Allergy. Immunol. 2012, 42, 58–70. [Google Scholar] [CrossRef]
- Bandyopadhyay, M.; Larregina, A.T. Keratinocyte-Polyamines and Dendritic Cells: A Bad Duet for Psoriasis. Immunity 2020, 53, 16–18. [Google Scholar] [CrossRef]
- Vona, R.; Giovannetti, A.; Gambardella, L.; Malorni, W.; Pietraforte, D.; Straface, E. Oxidative stress in the pathogenesis of systemic scleroderma: An overview. J. Cell. Mol. Med. 2018, 22, 3308–3314. [Google Scholar] [CrossRef]
- Zhu, H.; Chen, W.; Liu, D.; Luo, H. The role of metabolism in the pathogenesis of systemic sclerosis. Metabolism 2019, 93, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Napoli, C.; De Nigris, F.; Williams-Ignarro, S.; Pignalosa, O.; Sica, V.; Ignarro, L.J. Nitric oxide and atherosclerosis: An update. Nitric Oxide 2006, 15, 265–279. [Google Scholar] [CrossRef] [PubMed]
- D’Aniello, C.; Patriarca, E.J.; Phang, J.M.; Minchiotti, G. Proline Metabolism in Tumor Growth and Metastatic Progression. Front. Oncol. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zhu, W.; Chen, C.; Yan, B.; Zhu, L.; Chen, X.; Peng, C. The mechanisms of lysophosphatidylcholine in the development of diseases. Life Sci. 2020, 247, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J. Prix Fixe: Efferocytosis as a Four-Course Meal. Curr. Top. Microbiol. Immunol. 2017, 403, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Grossmayer, G.E.; Keppeler, H.; Boeltz, S.; Janko, C.; Rech, J.; Herrmann, M.; Lauber, K.; Muñoz, L.E. Elevated Serum Lysophosphatidylcholine in Patients with Systemic Lupus Erythematosus Impairs Phagocytosis of Necrotic Cells In Vitro. Front. Immunol. 2017, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Shea, B.S.; Tager, A.M. Sphingolipid regulation of tissue fibrosis. Open Rheumatol. J. 2012, 6, 123–129. [Google Scholar] [CrossRef]
- Huwiler, A.; Pfeilschifter, J. Sphingolipid signaling in renal fibrosis. Matrix Biol. 2018, 68–69, 230–247. [Google Scholar] [CrossRef]
- Geroldinger-Simić, M.; Bögl, T.; Himmelsbach, M.; Sepp, N.; Buchberger, W. Changes in Plasma Phospholipid Metabolism Are Associated with Clinical Manifestations of Systemic Sclerosis. Diagnostics (Basel) 2021, 11, 2116. [Google Scholar] [CrossRef]
- He, J.; Chan, T.; Hong, X.; Zheng, F.; Zhu, C.; Yin, L.; Dai, W.; Tang, D.; Liu, D.; Dai, Y. Microbiome and Metabolome Analyses Reveal the Disruption of Lipid Metabolism in Systemic Lupus Erythematosus. Front. Immunol. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- McKleroy, W.; Lee, T.-H.; Atabai, K. Always cleave up your mess: Targeting collagen degradation to treat tissue fibrosis. AJP 2013, 304, L709–L721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, H.M.; Ahn, S.H.; Choi, P.; Ko, Y.-A.; Han, S.H.; Chinga, F.; Park, A.S.D.; Tao, J.; Sharma, K.; Pullman, J.; et al. Defective fatty acid oxidation in renal tubular epithelial cells has a key role in kidney fibrosis development. Nat. Med. 2015, 21, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.L.; O’Neill, L.A. Reprogramming mitochondrial metabolism in macrophages as an anti-inflammatory signal. Eur. J. Immunol. 2016, 46, 13–21. [Google Scholar] [CrossRef]
- Zhao, X.; Kwan, J.Y.Y.; Yip, K.; Liu, P.P.; Liu, F.-F. Targeting metabolic dysregulation for fibrosis therapy. Nat. Rev. Drug Discov. 2020, 19, 57–75. [Google Scholar] [CrossRef]
- Volkmann, E.R.; Hoffmann-Vold, A.-M.; Chang, Y.-L.; Jacobs, J.P.; Tillisch, K.; Mayer, E.A.; Clements, P.J.; Hov, J.R.; Kummen, M.; Midtvedt, Ø.; et al. Systemic sclerosis is associated with specific alterations in gastrointestinal microbiota in two independent cohorts. BMJ Open Gastroenterol. 2017, 4, 1–9. [Google Scholar] [CrossRef]
- Visconti, A.; Le Roy, C.I.; Rosa, F.; Rossi, N.; Martin, T.C.; Mohney, R.P.; Li, W.; De Rinaldis, E.; Bell, J.T.; Venter, J.C.; et al. Interplay between the human gut microbiome and host metabolism. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bellocchi, C.; Fernández-Ochoa, Á.; Montanelli, G.; Vigone, B.; Santaniello, A.; Quirantes-Piné, R.; Borrás-Linares, I.; Gerosa, M.; Artusi, C.; Gualtierotti, R.; et al. Identification of a Shared Microbiomic and Metabolomic Profile in Systemic Autoimmune Diseases. J. Clin. Med. 2019, 8, 1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.C.; Brown, J.; Gong, M.; Ge, Y.; Zadeh, M.; Li, W.; Croker, B.P.; Michailidis, G.; Garrett, T.J.; Mohamadzadeh, M.; et al. Mansour Mohamadzadeh and Laurence Morel. Gut microbiota dysbiosis and altered tryptophan catabolism contribute to autoimmunity in lupus-susceptible mice. Sci. Transl. Med. 2020, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Singh, J.; Rattan, S.; DiMarino, A.J.; Cohen, S.; Jimenez, S.A. Review article: Pathogenesis and clinical manifestations of gastrointestinal involvement in systemic sclerosis. Aliment. Pharmacol. Ther. 2017, 45, 883–898. [Google Scholar] [CrossRef]
- He, Z.; Liu, Z.; Gong, L. Biomarker identification and pathway analysis of rheumatoid arthritis based on metabolomics in combination with ingenuity pathway analysis. Proteomics 2021, 21, 1–12. [Google Scholar] [CrossRef]
- Sanchez-Lopez, E.; Cheng, A.; Guma, M. Can Metabolic Pathways Be Therapeutic Targets in Rheumatoid Arthritis? J. Clin. Med. 2019, 8, 753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boros, F.; Vécsei, L. Progress in the development of kynurenine and quinoline-3-carboxamide-derived drugs. Expert. Opin. Investig. Drugs. 2020, 29, 1223–1247. [Google Scholar] [CrossRef]
- Hesselstrand, R.; Distler, J.H.W.; Riemekasten, G.; Wuttge, D.M.; Törngren, M.; Nyhlén, H.C.; Andersson, F.; Eriksson, H.; Sparre, B.; Tuvesson, H.; et al. An open-label study to evaluate biomarkers and safety in systemic sclerosis patients treated with paquinimod. Arthritis. Res. Ther. 2021, 23, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mondanelli, G.; Iacono, A.; Carvalho, A.; Orabona, C.; Volpi, C.; Pallotta, M.T.; Matino, D.; Esposito, S.; Grohmann, U. Amino acid metabolism as drug target in autoimmune diseases. Autoimmun. Rev. 2019, 18, 334–348. [Google Scholar] [CrossRef] [PubMed]
- Khanna, D.; Lin, C.J.F.; Furst, D.E.; Goldin, J.; Kim, G.; Kuwana, M.; Allanore, Y.; Matucci-Cerinic, M.; Distler, O.; Shima, Y.; et al. Tocilizumab in systemic sclerosis: A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet Respir. Med. 2020, 8, 963–974. [Google Scholar] [CrossRef]
- Azuma, A.; Nukiwa, T.; Tsuboi, E.; Suga, M.; Abe, S.; Nakata, K.; Taguchi, Y.; Nagai, S.; Itoh, H.; Ohi, M.; et al. Double-blind, placebo-controlled trial of pirfenidone in patients with idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care. Med. 2005, 171, 1040–1047. [Google Scholar] [CrossRef]
- Hua-Huy, T.; Le-Dong, N.-N.; Duong-Quy, S.; Bei, Y.; Rivière, S.; Tiev, K.-P.; Nicco, C.; Chéreau, C.; Batteux, F.; Dinh-Xuan, A.T. Increased exhaled nitric oxide precedes lung fibrosis in two murine models of systemic sclerosis. J. Breath. Res. 2015, 9, 1–15. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
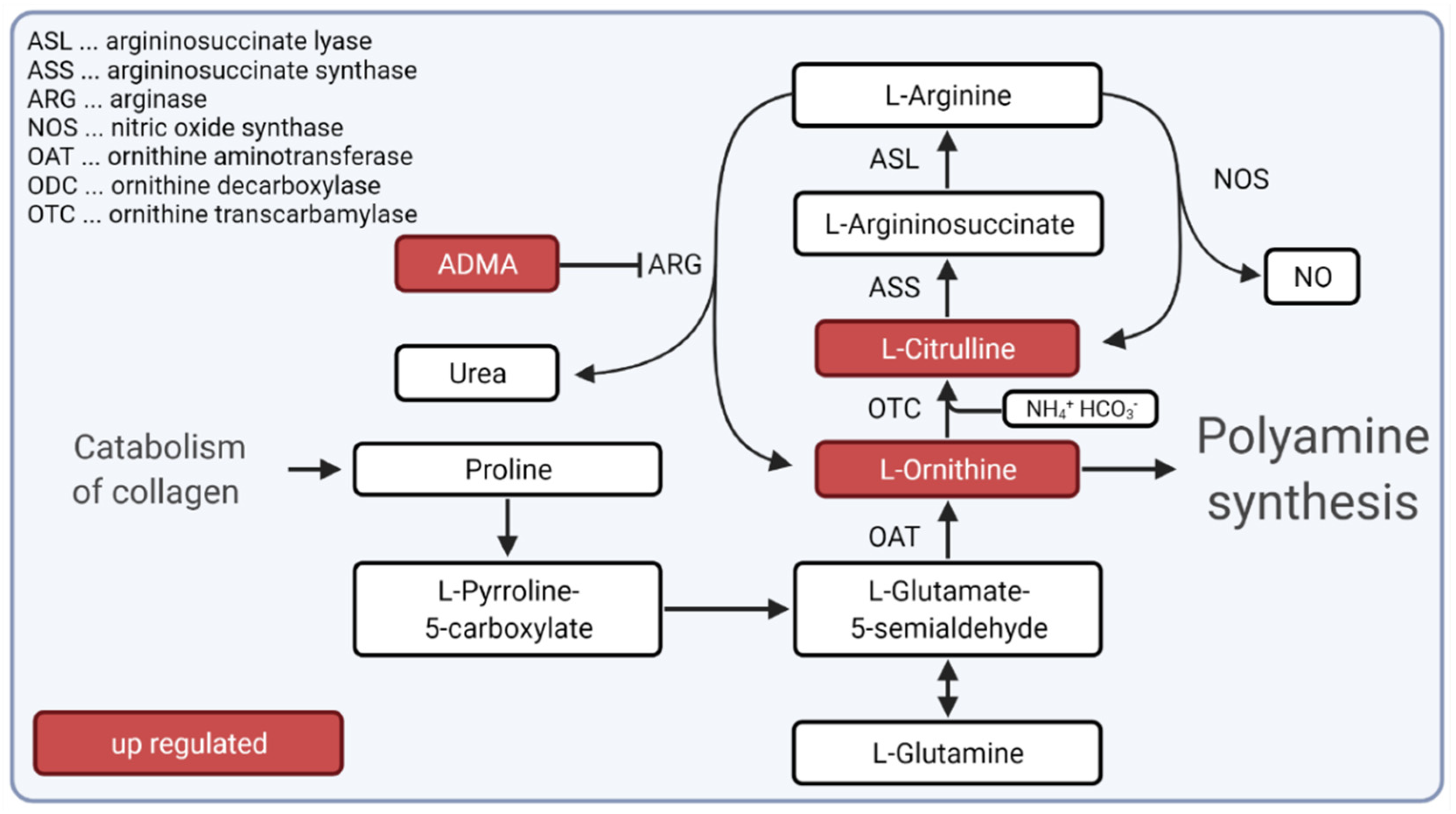{kind=link}
| Exact Mass (m/z) | RT/ (min) | Name | p-Value | Fold Change (SSc/Control) |
|---|---|---|---|---|
| 205.0971 | 14.7 | Tryptophan | <0.0001 | 0.3117 |
| 90.0549 | 18.2 | Alanine | <0.0001 | 0.3444 |
| 203.1503 | 23.2 | Dimethylarginine | <0.0001 | 2.6918 |
| 209.0921 | 14.5 | Kynurenine | 0.0004 | 2.1049 |
| 176.1030 | 22.1 | Citrulline | 0.0016 | 2.3760 |
| 133.0972 | 23.9 | Ornithine | 0.0026 | 2.3604 |
| Feature (RT_mz_DTCCSN2) | Metabolite ID | Name | CCSpred | m/zcalc | p-Value | Fold Change (SSc/Control) |
|---|---|---|---|---|---|---|
| 13.89_572.3679_236.76 | HMDB0010401 | LPC a 22:4 a (+H) | 243.3 | 572.3711 | <0.0001 | 0.4771 |
| 4.283_265.1168_160.02 | HMDB06344 | PAG (+H) | 159.3 | 265.1183 | <0.0001 | 2.1906 |
| 13.511_572.3676_238.65 | HMDB0010401 | LPC a 22:4 b (+H) | 243.3 | 572.3711 | 0.0002 | 0.4958 |
| 13.619_673.5254_274.79 | HMDB0240612 | SM 32:2 (+H) | 273.9 | 673.5279 | 0.0020 | 0.7053 |
| 15.977_221.0915_150.56 | HMDB0000472 | OH-tryptophan (+H) | 149.7 | 221.0926 | 0.0055 | 0.6392 |
| 13.987_548.3692_234.01 | HMDB0010392 | LPC a 20:2 (+H) | 240.7 | 548.3711 | 0.0079 | 0.5156 |
| 13.472_685.5613_286.32 | HMDB0013464 | SM 34:1 (−H2O +H) | 283.5 | 685.5649 | 0.0101 | 0.9840 |
| 13.203_783.6341_296.53 | HMDB0240670 | SM 40:3 (+H) | 291.2 | 783.6375 | 0.0152 | 0.7605 |
| 12.479_248.1485_156.00 | HMDB0013127 | OH-butyrylcarnitine (+H) | 158.4 | 248.1498 | 0.0219 | 1.2434 |
| 7.364_332.2419_193.87 | HMDB0061636 | OH-decanoylcarnitine (+H) | 189.3 | 332.2437 | 0.0396 | 1.6815 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bögl, T.; Mlynek, F.; Himmelsbach, M.; Sepp, N.; Buchberger, W.; Geroldinger-Simić, M. Plasma Metabolomic Profiling Reveals Four Possibly Disrupted Mechanisms in Systemic Sclerosis. Biomedicines 2022, 10, 607. https://doi.org/10.3390/biomedicines10030607
Bögl T, Mlynek F, Himmelsbach M, Sepp N, Buchberger W, Geroldinger-Simić M. Plasma Metabolomic Profiling Reveals Four Possibly Disrupted Mechanisms in Systemic Sclerosis. Biomedicines. 2022; 10(3):607. https://doi.org/10.3390/biomedicines10030607
Chicago/Turabian StyleBögl, Thomas, Franz Mlynek, Markus Himmelsbach, Norbert Sepp, Wolfgang Buchberger, and Marija Geroldinger-Simić. 2022. "Plasma Metabolomic Profiling Reveals Four Possibly Disrupted Mechanisms in Systemic Sclerosis" Biomedicines 10, no. 3: 607. https://doi.org/10.3390/biomedicines10030607
APA StyleBögl, T., Mlynek, F., Himmelsbach, M., Sepp, N., Buchberger, W., & Geroldinger-Simić, M. (2022). Plasma Metabolomic Profiling Reveals Four Possibly Disrupted Mechanisms in Systemic Sclerosis. Biomedicines, 10(3), 607. https://doi.org/10.3390/biomedicines10030607