
Potential Gonado-Protective Effect of Cichorium endivia and Its Major Phenolic Acids against Methotrexate-Induced Testicular Injury in Mice

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Instruments and Chemicals
2.2. Plant Material Collection and Extraction
2.3. Determination of Total Phenolic Content “(TPC) Assay”
2.4. Evaluation of Antioxidant Activity
2.4.1. DPPH Radical-Scavenging Activity
2.4.2. Ferric Reducing Antioxidant Power (FRAP) Assay
2.4.3. Total Antioxidant Capacity (TAC) Assay
2.5. Fractionation of C. endivia L. Extract and Isolation of Phytochemical Constituents
2.6. Preparation of the Phenolic Fraction of C. endivia L. Extract
2.7. In Vivo Study
2.7.1. Animals
2.7.2. Induction of Testicular Injury
2.7.3. Study Design
2.7.4. Collection of the Samples
2.7.5. Histopathological Examination
2.7.6. Determination of Biochemical Parameters
Determination of Testosterone
Determination of Oxidative Stress Biomarkers
Determination of Inflammatory Markers and Apoptosis-Related Proteins
Determination of Testicular Levels of NF-κB, TNF-α, p53, and miR-29a by Quantitative Real-Time PCR
2.7.7. Statistical Analysis
3. Results
3.1. Total Phenolic Content (TPC)
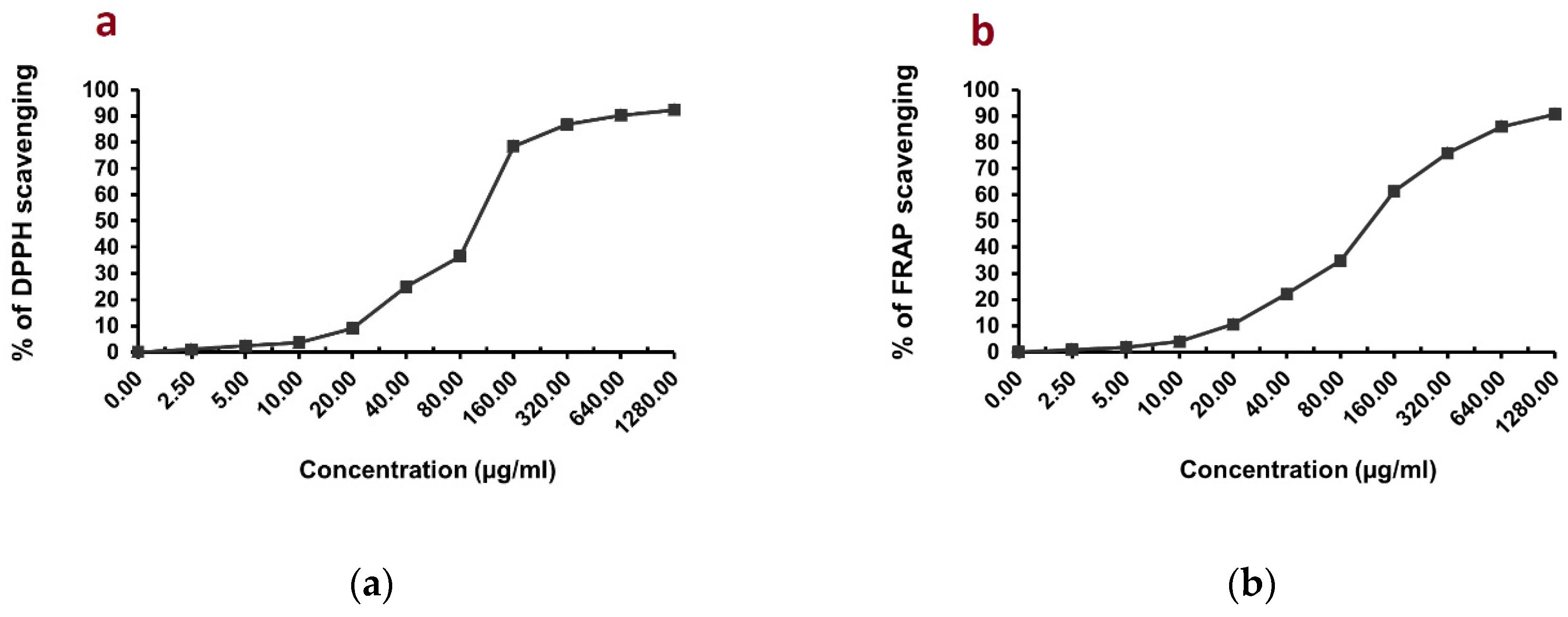
3.2. Evaluation of In Vitro Antioxidant Activity of C. endivia L. Extract
3.3. Chemical Identification of the Isolated Compounds
3.4. In Vivo Study
3.4.1. Histopathological Finding
3.4.2. Effect on Biochemical Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amer, A.M. Antimicrobial effects of Egyptian local chicory, Cichorium endivia subsp. pumilum. Int. J. Microbiol. 2018, 2018, 6475072. [Google Scholar] [PubMed]
- Masoud, M.; Zayed, M.A.; Gad, D.; Elhaak, M.A. Effect of gamma irradiation on some active constituents and metabolites of Cichorium pumilum Jacq. Egypt. J. Exp. Biol. 2018, 14, 153–159. [Google Scholar]
- Koudela, M.; Petříková, K. Nutritional composition and yield of endive cultivars–Cichorium endivia L. Hortic. Sci. 2007, 34, 6–10. [Google Scholar] [CrossRef]
- Abou-Zeid, N.A. Utilization of the weed Cichorium pumilum Jacq. as vegetable protease enzymes for whey protein hydrolysate production. Int. J. Res. Stud. Biosci. 2015, 3, 44–53. [Google Scholar]
- Azaizeh, H.; Saad, B.; Khalil, K.; Said, O. The state of the art of traditional Arab herbal medicine in the Eastern region of the Mediterranean: A review. Evid. Based Complement. Altern. 2006, 3, 229–235. [Google Scholar] [CrossRef]
- Al Khateeb, W.; Hussein, E.; Qouta, L.; Alu’datt, M.; Al-Shara, B.; Abu-Zaiton, A. In vitro propagation and characterization of phenolic content along with antioxidant and antimicrobial activities of Cichorium pumilum Jacq. Plant Cell Tissue Organ Cult. 2012, 110, 103–110. [Google Scholar] [CrossRef]
- Seto, M.; Miyase, T.; Umehara, K.; Ueno, A.; Hirano, Y.; Otani, N. Sesquiterpene lactones from Cichorium endivia L. and C. intybus L. and cytotoxic activity. Chem. Pharm. Bull. 1988, 36, 2423–2429. [Google Scholar] [CrossRef]
- Kisiel, W.; Michalska, K. Root constituents of Cichorium pumilum and rearrangements of some lactucin-like guaianolides. Z. Naturforsch. C 2003, 58, 789–792. [Google Scholar] [CrossRef]
- Kisiel, W.; Michalska, K. Sesquiterpenoids and phenolics from roots of Cichorium endivia var. crispum. Fitoterapia 2006, 77, 354–357. [Google Scholar] [CrossRef]
- Warashina, T.; Miyase, T. Sesquiterpenes from the Roots of Cichorium endivia. Chem. Pharm. Bull. 2008, 56, 1445–1451. [Google Scholar] [CrossRef]
- Saleh, M.R.; Metwally, A.M.; Amer, M.M. Isolation of a flavonoidal substance from Cichorium pumilum jacq. Pharmazie 1975, 30, 404. [Google Scholar] [PubMed]
- Chen, C.J.; Deng, A.J.; Liu, C.; Shi, R.; Qin, H.L.; Wang, A.P. Hepatoprotective activity of Cichorium endivia L. extract and its chemical constituents. Molecules 2011, 16, 9049–9066. [Google Scholar] [CrossRef] [PubMed]
- El-Shafey, N.M.; AbdElgawad, H. Luteolin, a bioactive flavone compound extracted from Cichorium endivia L. subsp. divaricatum alleviates the harmful effect of salinity on maize. Acta Physiol. Plant. 2012, 34, 2165–2177. [Google Scholar] [CrossRef]
- Mascherpa, D.; Carazzone, C.; Marrubini, G.; Gazzani, G.; Papetti, A. Identification of phenolic constituents in Cichorium endivia var. crispum and var. latifolium salads by high-performance liquid chromatography with diode array detection and electrospray ioniziation tandem mass spectrometry. J. Agric. Food Chem. 2012, 60, 12142–12150. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, A.; Ezzat, S.; Qasem, I.; Ali-Shtayeh, M.; Basalah, M.; Ali, H.; Hatamleh, A. Diversity of active constituents in Cichorium endivia and Cynara cornigera extracts. Acta Biol. Hung. 2015, 66, 103–118. [Google Scholar] [CrossRef]
- Khalil, H.E.; Kamel, M.S. Phytochemical and biological studies of Cichorium endivia L. leaves. J. Pharm. Sci. 2015, 7, 509–513. [Google Scholar]
- Papetti, A.; Daglia, M.; Aceti, C.; Sordelli, B.; Spini, V.; Carazzone, C.; Gazzani, G. Hydroxycinnamic acid derivatives occurring in Cichorium endivia vegetables. J. Pharm. Biomed. 2008, 48, 472–476. [Google Scholar] [CrossRef]
- Singab, A.B.; Ayoub, N.A.; Noaman, E.; Ayoub, I.M. Phytochemical constituents, antioxidant and hepatoprotective activities of Egyptian Cichorium pumilum Jacq. Bull. Fac. Pharm. Cairo Univ. 2010, 48, 13. [Google Scholar]
- Mikropoulou, E.V.; Vougogiannopoulou, K.; Kalpoutzakis, E.; Sklirou, A.D.; Skaperda, Z.; Houriet, J.; Wolfender, J.L.; Trougakos, I.P.; Kouretas, D.; Halabalaki, M.; et al. Phytochemical composition of the decoctions of Greek edible greens (chórta) and evaluation of antioxidant and cytotoxic properties. Molecules 2018, 23, 1541. [Google Scholar] [CrossRef]
- Wang, F.X.; Deng, A.J.; Wei, J.F.; Qin, H.L.; Wang, A.P. 1H and 13C NMR Assignments of Cytotoxic 3S-1,2,3,4-Tetrahydro-β-carboline-3-carboxylic Acid from the Leaves of Cichorium endivia. J. Anal. Methods Chem. 2012, 2012, 254391. [Google Scholar] [CrossRef]
- Aisa, H.A.; Xin, X.L.; Tang, D. Chemical constituents and their pharmacological activities of plants from Cichorium genus. Chin. Herb. Med. 2020, 12, 224–236. [Google Scholar] [CrossRef]
- Han, C.; Wu, X.; Zou, N.; Zhang, Y.; Yuan, J.; Gao, Y.; Chen, W.; Yao, J.; Li, C.; Hou, J.; et al. Cichorium pumilum Jacq Extract Inhibits LPS-Induced Inflammation via MAPK Signaling Pathway and Protects Rats from Hepatic Fibrosis Caused by Abnormalities in the Gut-Liver Axis. Front. Pharmacol. 2021, 12, 683613. [Google Scholar] [CrossRef] [PubMed]
- Kamel, Z.H.; Daw, I.; Marzouk, M. Effect of Cichorium endivia leaves on some biochemical parameters in streptozotocin-induced diabetic rats. Aust. J. Basic Appl. Sci. 2011, 5, 387–396. [Google Scholar]
- Wang, F.X.; Deng, A.J.; Li, M.; Wei, J.F.; Qin, H.L.; Wang, A.P. (3S)-1,2,3,4-Tetrahydro-β-carboline-3-carboxylic acid from Cichorium endivia. L induces apoptosis of human colorectal cancer HCT-8 cells. Molecules 2012, 18, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Alshehri, A.; Elsayed, H.E. Molecular and biochemical evaluation of anti-proliferative effect of (Cichorium endivia, L.) phenolic extracts on breast cancer cell line: MCF7. E3 J. Biotechnol. Pharm. Res. 2012, 3, 74–82. [Google Scholar]
- Al-Akhras, M.A.H.; Aljarrah, K.; Al-Khateeb, H.; Jaradat, A.; Al-Omari, A.; Al-Nasser, A.; Masadeh, M.M.; Amin, A.; Hamza, A.; Mohammed, K.; et al. Introducing Cichorium pumilum as a potential therapeutical agent against drug-induced benign breast tumor in rats. Electromagn. Biol. Med. 2012, 31, 299–309. [Google Scholar] [CrossRef]
- Enk, C.D.; Hochberg, M.; Torres, A.; Lev, O.; Dor, I.; Srebnik, M.; Dembitsky, V.M. Photoprotection by Cichorium endivia extracts: Prevention of UVB-induced erythema, pyrimidine dimer formation and IL-6 expression. Skin Pharmacol. Physiol. 2004, 17, 42–48. [Google Scholar] [CrossRef]
- Fu, J.; Liu, Y.; Wang, C.; Zhang, H.; Yu, B.; Wang, Y.; Zhu, H. Persistent follicular granulosa cell senescence and apoptosis induced by methotrexate leading to oocyte dysfunction and aberrant embryo development. Clin. Transl. Sci. 2021, 14, 2043–2054. [Google Scholar] [CrossRef]
- Sherif, I.O.; Al-Mutabagani, L.A.; Sarhan, O.M. Ginkgo biloba extract attenuates methotrexate-induced testicular injury in rats: Cross-talk between oxidative stress, inflammation, apoptosis, and miRNA-29a expression. Integr. Cancer Ther. 2020, 19, 1534735420969814. [Google Scholar] [CrossRef]
- Elhady, S.S.; Abdelhameed, R.F.A.; Mehanna, E.T.; Wahba, A.S.; Elfaky, M.A.; Koshak, A.E.; Noor, A.O.; Bogari, H.A.; Malatani, R.T.; Goda, M.S. Metabolic Profiling, Chemical Composition, Antioxidant Capacity, and In Vivo Hepato- and Nephroprotective Effects of Sonchus cornutus in Mice Exposed to Cisplatin. Antioxidants 2022, 11, 819. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin–Ciocalteau reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Yen, G.C.; Duh, P.D. Scavenging effect of methanolic extracts of peanut hulls on free radical and active oxygen species. J. Agric. Food Chem. 1994, 42, 629. [Google Scholar] [CrossRef]
- Gomha, S.M.; Muhammad, Z.A.; Abdel-Aziz, M.R.; Abdel-Aziz, H.M.; Gaber, H.M.; Elaasser, M.M. One-pot synthesis of new thiadiazolyl-pyridines as anticancer and antioxidant agents. J. Heterocycl. Chem. 2018, 55, 530–536. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reaction: Antioxidative activities of products of browning reaction prepared from glucosamine. Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Ferreira, I.C.F.R.; Baptista, P.; Vilas-Boas, M.; Barros, L. Free-radical scavenging capacity and reducing power of wild edible mushrooms from northeast Portugal: Individual cap and stipe activity. Food Chem. 2007, 100, 1511–1516. [Google Scholar] [CrossRef]
- Canabady-Rochelle, L.L.; Harscoat-Schiavo, C.; Kessler, V.; Aymes, A.; Fournier, F.; Girardet, J.M. Determination of reducing power and metal chelating ability of antioxidant peptides: Revisited methods. Food Chem. 2015, 183, 129–135. [Google Scholar] [CrossRef]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef]
- Vijayalakshmi, A.; Kumar, P.R.; Priyadarsini, S.S.; Meenaxshi, C. In vitro antioxidant and anticancer activity of flavonoid fraction from the aerial parts of Cissus quadrangularis Linn. against human breast carcinoma cell lines. J. Chem. 2013, 2013, 150675. [Google Scholar] [CrossRef]
- Johnsen, S.G. Testicular biopsy score count–a method for registration of spermatogenesis in human testes: Normal values and results in 335 hypogonadal males. Horm. Res. Paediatr. 1970, 1, 2–25. [Google Scholar] [CrossRef]
- Cosme, P.; Rodríguez, A.B.; Espino, J.; Garrido, M. Plant phenolics: Bioavailability as a key determinant of their potential health-promoting applications. Antioxidants 2020, 9, 1263. [Google Scholar] [CrossRef]
- Ghosh, T.; Maity, T.K.; Singh, J. Evaluation of antitumor activity of stigmasterol, a constituent isolated from Bacopa monnieri Linn aerial parts against Ehrlich Ascites Carcinoma in mice. Orient. Pharm. Exp. Med. 2011, 11, 41–49. [Google Scholar] [CrossRef]
- Parvez, M.K.; Alam, P.; Arbab, A.H.; Al-Dosari, M.S.; Alhowiriny, T.A.; Alqasoumi, S.I. Analysis of antioxidative and antiviral biomarkers β-amyrin, β-sitosterol, lupeol, ursolic acid in Guiera senegalensis leaves extract by validated HPTLC methods. Saudi Pharm. J. 2018, 26, 685–693. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Guo, Y.J.; Gao, Y.Y. Studies on chemical constituents of root of Cichorium intybus. China J. Chin. Mater. Med. 2002, 27, 209–210. [Google Scholar]
- Chang, S.W.; Kim, K.H.; Lee, I.K.; Choi, S.U.; Ryu, S.Y.; Lee, K.R. Phytochemical constituents of Bistorta manshuriensis. Nat. Prod. Sci. 2009, 15, 234–240. [Google Scholar]
- Kisiel, W.; Michalska, K.; Szneler, E. Norisoprenoids from aerial parts of Cichorium pumilum. Biochem. Syst. Ecol. 2004, 32, 343–346. [Google Scholar] [CrossRef]
- Hammami, S.; Jannet, H.B.; Bergaoui, A.; Ciavatta, L.; Cimino, G.; Mighri, Z. Isolation and structure elucidation of a flavanone, a flavanone glycoside and vomifoliol from Echiochilon fruticosum growing in Tunisia. Molecules 2004, 9, 602–608. [Google Scholar] [CrossRef]
- Huynh, N.V.; Nguyen, T.H.T.; Nguyen, K.P.P.; Hansen, P.E. Structural studies of the chemical constituents of Tithonia tagetiflora Desv. (Asteraceae). Magn. Reson. Chem. 2013, 51, 439–443. [Google Scholar] [CrossRef]
- Al-Madhagy, S.; Mostafa, N.M.; Youssef, F.S.; Awad, G.; Eldahshan, O.A.; Singab, A.N.B. Isolation and structure elucidation of compounds from Coccinia grandis leaves extract. Egypt. J. Chem. 2019, 62, 1869–1877. [Google Scholar] [CrossRef]
- Altemimi, A.B.; Mohammed, M.J.; Yi-Chen, L.; Watson, D.G.; Lakhssassi, N.; Cacciola, F.; Ibrahim, S.A. Optimization of Ultrasonicated Kaempferol Extraction from Ocimum basilicum Using a Box–Behnken Design and Its Densitometric Validation. Foods 2020, 9, 1379. [Google Scholar] [CrossRef]
- Jesionek, W.; Majer-Dziedzic, B.; Choma, I.M. Separation, identification, and investigation of antioxidant ability of plant extract components using TLC, LC–MS, and TLC–DPPH•. J. Liq. Chromatogr. Relat. Technol. 2015, 38, 1147–1153. [Google Scholar] [CrossRef]
- Berker, K.I.; Olgun, F.A.O.; Ozyurt, D.; Demirata, B.; Apak, R. Modified Folin–Ciocalteu antioxidant capacity assay for measuring lipophilic antioxidants. J. Agric. Food Chem. 2013, 61, 4783–4791. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, I.G.; Apetrei, C. Analytical Methods Used in Determining Antioxidant Activity: A Review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef] [PubMed]
- Bibi Sadeer, N.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The Versatility of Antioxidant Assays in Food Science and Safety—Chemistry, Applications, Strengths, and Limitations. Antioxidants 2020, 9, 709. [Google Scholar] [CrossRef] [PubMed]
- Malik, B.; Pirzadah, T.B.; Tahir, I.; Rehman, R.U. Chemo-profiling, Antioxidant potential and Ionomic analysis of Cichorium intybus L. Pharmacogn. J. 2017, 9, 917–928. [Google Scholar] [CrossRef]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Eltamany, E.E.; Elhady, S.S.; Ahmed, H.A.; Badr, J.M.; Noor, A.O.; Ahmed, S.A.; Nafie, M.S. Chemical Profiling, Antioxidant, Cytotoxic Activities and Molecular Docking Simulation of Carrichtera annua DC. (Cruciferae). Antioxidants 2020, 9, 1286. [Google Scholar] [CrossRef]
- Elhusseiny, S.M.; El-Mahdy, T.S.; Awad, M.F.; Elleboudy, N.S.; Farag, M.M.S.; Yassein, M.A.; Aboshanab, K.M. Proteome Analysis and In Vitro Antiviral, Anticancer and Antioxidant Capacities of the Aqueous Extracts of Lentinula edodes and Pleurotus ostreatus Edible Mushrooms. Molecules 2021, 26, 4623. [Google Scholar] [CrossRef]
- Apak, R.; Özyürek, M.; Güçlü, K.; Çapanoğlu, E. Antioxidant Activity/Capacity Measurement. 2. Hydrogen Atom Transfer (HAT)-Based, Mixed-Mode (Electron Transfer (ET)/HAT), and Lipid Peroxidation Assays. J. Agric. Food Chem. 2016, 64, 1028–1045. [Google Scholar] [CrossRef]
- Uddin, N.; Mitra, K.; Haque, Z. Comparative Bio-Active Compounds Determination and In Vitro Antioxidant Properties of Newly Developed Soy Mixed Wheat Flour and Traditional Wheat Flour. Int. J. Food Prop. 2015, 19, 2113–2126. [Google Scholar] [CrossRef]
- Diwan, R.; Shinde, A.; Malpathak, N. Phytochemical Composition and Antioxidant Potential of Ruta graveolens L. In Vitro Culture Lines. J. Bot. 2012, 2012, 685427. [Google Scholar] [CrossRef]
- Marzouk, M.; Sayed, A.; Soliman, A. Hepatoprotective and antioxidant effects of Cichorium endivia L. leaves extract against acetaminophen toxicity on rats. J. Med. Med. Sci. 2011, 2, 1273–1279. [Google Scholar]
- Lfitat, A.; Zejli, H.; Bousraf, F.Z.; Bousselham, A.; El Atki, Y.; Gourch, A.; Lyoussi, B.; Abdellaoui, A. Comparative Assessment of Total Phenolics Content and in Vitro Antioxidant Capacity Variations of Macerated Leaf Extracts of Olea Europaea L. and Argania spinosa (L.) Skeels. Mater. Today Proc. 2021, 45, 7271–7277. [Google Scholar] [CrossRef]
- Hinneburg, I.; Dorman, H.J.D.; Hiltunen, R. Antioxidant Activities of Extracts from Selected Culinary Herbs and Spices. Food Chem. 2006, 97, 122–129. [Google Scholar] [CrossRef]
- Rosa, L.D.S.; Jordão, N.A.; Soares, N.D.C.P.; Mesquita, J.F.D.; Monteiro, M.; Teodoro, A.J. Pharmacokinetic, antiproliferative and apoptotic effects of phenolic acids in human colon adenocarcinoma cells using in vitro and in silico approaches. Molecules 2018, 23, 2569. [Google Scholar] [CrossRef] [PubMed]
- Kagambega, W.; Belem, H.; Meda, R.N.T.; Koama, B.K.; Drabo, A.F.; Kabore, J.; Traore, A.; Ouédraogo, G.A.; Benedec, D.; Hanganu, D.; et al. Polyphenolic Profile, Anti-Inflammatory and Anti-Nociceptive Activities of Some African Medicinal Plants. Plants 2022, 11, 1377. [Google Scholar] [CrossRef] [PubMed]
- Shafik, A. Intratunical injection of methotrexate for the treatment of seminoma of the testicle. Anticancer Drugs 1993, 4, 193–195. [Google Scholar] [CrossRef] [PubMed]
- Sussman, A.; Leonard, J.M. Psoriasis, methotrexate, and oligospermia. Arch. Dermatol. 1980, 116, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Van Scott, E.J.; Reinertson, R.P. Morphologic and physiologic effects of chemotherapeutic agents in psoriasis. J. Investig. Dermatol. 1959, 33, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Pınar, N.; Çakırca, G.; Özgür, T.; Kaplan, M. The protective effects of alpha lipoic acid on methotrexate induced testis injury in rats. Biomed. Pharmacother. 2018, 97, 1486–1492. [Google Scholar] [CrossRef]
- Abdelzaher, W.Y.; Khalaf, H.M.; El-Hussieny, M.; Bayoumi, A.M.A.; Shehata, S.; Refaie, M.M.M. Role of nitric oxide donor in methotrexate-induced testicular injury via modulation of pro-inflammatory mediators, eNOS and P-glycoprotein. Hum. Exp. Toxicol. 2020, 39, 1700–1709. [Google Scholar] [CrossRef]
- Bouwmeester, T.; Bauch, A.; Ruffner, H.; Angrand, P.O.; Bergamini, G.; Croughton, K.; Cruciat, C.; Eberhard, D.; Gagneur, J.; Ghidelli, S.; et al. A physical and functional map of the human TNF-α/NF-κB signal transduction pathway. Nat. Cell Biol. 2004, 6, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, T.T.; Zhao, H.Y.; Wang, H. Melatonin protects methotrexate-induced testicular injury in rats. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7517–7525. [Google Scholar] [PubMed]
- Owumi, S.E.; Ochaoga, S.E.; Odunola, O.A.; Farombi, E.O. Protocatechuic acid inhibits testicular and epididymal toxicity associated with methotrexate in rats. Andrologia 2019, 51, e13350. [Google Scholar] [CrossRef] [PubMed]
- Samani, S.M.; Bojnordi, T.E.; Zarghampour, M.; Merat, S.; Fouladi, D.F. Expression of p53, Bcl-2 and Bax in endometrial carcinoma, endometrial hyperplasia and normal endometrium: A histopathological study. J. Obstet. Gynaecol. 2018, 38, 999–1004. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Sun, H.; Wei, W.; Liu, Q.; Wang, Y.; Zhang, Y.; Lian, F.; Liu, F.; Li, C.; Ying, K.; et al. Mitoxantrone triggers immunogenic prostate cancer cell death via p53-dependent PERK expression. Cell. Oncol. 2020, 43, 1099–1116. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zheng, J.; Wang, Y.; Gao, Z. miRNA-29a inhibits colon cancer growth by regulation of the PTEN/Akt/GSK3β and Wnt/β-catenin signaling pathways. Oncol. Lett. 2018, 16, 2638–2644. [Google Scholar] [CrossRef]
- Li, R.; Liu, J.; Li, Q.; Chen, G.; Yu, X. miR-29a suppresses growth and metastasis in papillary thyroid carcinoma by targeting AKT3. Tumor Biol. 2016, 37, 3987–3996. [Google Scholar] [CrossRef]
- Park, S.Y.; Lee, J.H.; Ha, M.; Nam, J.W.; Kim, V.N. miR-29 miRNAs activate p53 by targeting p85α and CDC42. Nat. Struct. Mol. Biol. 2009, 16, 23–29. [Google Scholar] [CrossRef]
- Akinjo, O.O.; Gant, T.W.; Marczylo, E.L. Perturbation of epigenetic processes by doxorubicin in the mouse testis. Toxicol. Res. 2016, 5, 1229–1243. [Google Scholar] [CrossRef]
- Meunier, L.; Siddeek, B.; Vega, A.; Lakhdari, N.; Inoubli, L.; Bellon, R.P.; Lemaire, G.; Mauduit, C.; Benahmed, M. Perinatal programming of adult rat germ cell death after exposure to xenoestrogens: Role of microRNA miR-29 family in the downregulation of DNA methyltransferases and Mcl-1. Endocrinology 2012, 153, 1936–1947. [Google Scholar] [CrossRef]
- Eltamany, E.E.; Elhady, S.S.; Nafie, M.S.; Ahmed, H.A.; Abo-Elmatty, D.M.; Ahmed, S.A.; Badr, J.M.; Abdel-Hamed, A.R. The Antioxidant Carrichtera annua DC. Ethanolic Extract Counteracts Cisplatin Triggered Hepatic and Renal Toxicities. Antioxidants 2021, 10, 825. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kim, K.S.; Son, J.Y.; Kim, H.R.; Park, J.H.; Lee, S.H.; Lee, D.E.; Kim, I.S.; Lee, K.Y.; Lee, B.M.; et al. Protective effects of Dendropanax morbifera against cisplatin-induced nephrotoxicity without altering chemotherapeutic efficacy. Antioxidants 2019, 8, 256. [Google Scholar] [CrossRef] [PubMed]
- Soliman, A.M.; Desouky, S.; Marzouk, M.; Sayed, A.A. Origanum majorana attenuates nephrotoxicity of cisplatin anticancer drug through ameliorating oxidative stress. Nutrients 2016, 8, 264. [Google Scholar] [CrossRef] [PubMed]
- Heidari-Soreshjani, S.; Asadi-Samani, M.; Yang, Q.; Saeedi-Boroujeni, A. Phytotherapy of nephrotoxicity-induced by cancer drugs: An updated review. J. Nephropathol. 2017, 6, 254–263. [Google Scholar] [CrossRef]
- Ilyas, S.; Tabasum, R.; Iftikhar, A.; Nazir, M.; Hussain, A.; Hussain, A.; Ali, M.S.; Saleem, F.; Saleem, U.; Froeyen, M.; et al. Effect of Berberis vulgaris L. root extract on ifosfamide-induced in vivo toxicity and in vitro cytotoxicity. Sci. Rep. 2021, 11, 1708. [Google Scholar] [CrossRef] [PubMed]
- Dorostghoal, M.; Seyyednejad, S.M.; Nejad, M.N.T. Cichorium intybus L. extract ameliorates testicular oxidative stress induced by lead acetate in male rats. Clin. Exp. Reprod. Med. 2020, 47, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Dorostghoal, M.; Seyyednejad, S.M.; Nejad, M.N.T. Beneficial effects of Cichorium intybus L. extract on oxidative status and reproductive parameters in male Wistar rats: An experimental study. Int. J. Reprod. Biomed. 2019, 17, 425–434. [Google Scholar] [CrossRef]
- Nishi, K.; Ramakrishnan, S.; Gunasekaran, V.P.; Parkash, K.; Ramakrishnan, A.; Vijayakumar, N.; Ganeshan, M. Protective effects of p-coumaric acid on ethanol induced male reproductive toxicity. Life Sci. 2018, 209, 1–8. [Google Scholar] [CrossRef]
- Zahra, Z.; Khan, M.R.; Majid, M.; Maryam, S.; Sajid, M. Gonadoprotective ability of Vincetoxicum arnottianum extract against bisphenol A-induced testicular toxicity and hormonal imbalance in male Sprague Dawley rats. Andrologia 2020, 52, e13590. [Google Scholar] [CrossRef]
- Abd El Tawab, A.M.; Shahin, N.N.; AbdelMohsen, M.M. Protective effect of Satureja montana extract on cyclophosphamide-induced testicular injury in rats. Chem. Biol. Interact. 2014, 224, 196–205. [Google Scholar] [CrossRef]
- Hassanein, E.H.; Abdel-Wahab, B.A.; Ali, F.E.; El-Ghafar, A.; Omnia, A.M.; Kozman, M.R.; Sharkawi, S.M. Trans-ferulic acid ameliorates cisplatin-induced testicular damage via suppression of TLR4, P38-MAPK, and ERK1/2 signaling pathways. Environ. Sci. Pollut. Res. 2021, 28, 41948–41964. [Google Scholar] [CrossRef] [PubMed]
- Roychoudhury, S.; Sinha, B.; Choudhury, B.P.; Jha, N.K.; Palit, P.; Kundu, S.; Mandal, S.C.; Kolesarova, A.; Yousef, M.I.; Ruokolainen, J.; et al. Scavenging properties of plant-derived natural biomolecule para-coumaric acid in the prevention of oxidative stress-induced diseases. Antioxidants 2021, 10, 1205. [Google Scholar] [CrossRef] [PubMed]
- Saad, A.B.; Rjeibi, I.; Brahmi, N.; Elaloui, E.; Zouari, N. Nicotine-induced oxidative stress, testis injury, AChE inhibition and brain damage alleviated by Mentha spicata. Inflammopharmacology 2020, 28, 939–948. [Google Scholar] [CrossRef]
- Sindhu, G.; Nishanthi, E.; Sharmila, R. Nephroprotective effect of vanillic acid against cisplatin induced nephrotoxicity in wistar rats: A biochemical and molecular study. Environ. Toxicol. Pharmacol. 2015, 39, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Kassab, R.B.; Lokman, M.S.; Daabo, H.M.; Gaber, D.A.; Habotta, O.A.; Hafez, M.M.; Zhery, A.S.; Moneim, A.E.A.; Fouda, M.S. Ferulic acid influences Nrf2 activation to restore testicular tissue from cadmium-induced oxidative challenge, inflammation, and apoptosis in rats. J. Food Biochem. 2020, 44, e13505. [Google Scholar] [CrossRef]
- Hasanein, P.; Fazeli, F.; Parviz, M.; Roghani, M. Ferulic acid prevents lead-induced testicular oxidative stress and suppressed spermatogenesis in rats. Andrologia 2018, 50, e12798. [Google Scholar] [CrossRef]
- Chandra, K.; Jain, V.; Jabin, A.; Dwivedi, S.; Joshi, S.; Ahmad, S.; Jain, S.K. Effect of Cichorium intybus seeds supplementation on the markers of glycemic control, oxidative stress, inflammation, and lipid profile in type 2 diabetes mellitus: A randomized, double-blind placebo study. Phytother. Res. 2020, 34, 1609–1618. [Google Scholar] [CrossRef]
- Olsen, N.J.; Branch, V.K.; Jonnala, G.; Seskar, M.; Cooper, M. Phase 1, placebo-controlled, dose escalation trial of chicory root extract in patients with osteoarthritis of the hip or knee. BMC Musculoskelet. Disord. 2010, 11, 156. [Google Scholar] [CrossRef]
- Ghamarian, A.; Abdollahi, M.; Su, X.; Amiri, A.; Ahadi, A.; Nowrouzi, A. Effect of chicory seed extract on glucose tolerance test (GTT) and metabolic profile in early and late stage diabetic rats. DARU J. Pharm. Sci. 2012, 20, 56. [Google Scholar] [CrossRef]
- Hsu, C.L.; Fang, S.C.; Yen, G.C. Anti-inflammatory effects of phenolic compounds isolated from the flowers of Nymphaea mexicana Zucc. Food Funct. 2013, 4, 1216–1222. [Google Scholar] [CrossRef]
- Fang, S.C.; Hsu, C.L.; Yen, G.C. Anti-inflammatory effects of phenolic compounds isolated from the fruits of Artocarpus heterophyllus. J. Agric. Food Chem. 2008, 56, 4463–4468. [Google Scholar] [CrossRef] [PubMed]
- Sabitha, R.; Nishi, K.; Gunasekaran, V.P.; Annamalai, G.; Agilan, B.; Ganeshan, M. p-Coumaric acid ameliorates ethanol–induced kidney injury by inhibiting inflammatory cytokine production and NF–κB signaling in rats. Asian Pac. J. Trop. Biomed. 2019, 9, 188–195. [Google Scholar]
- Abdel-Moneim, A.; Yousef, A.I.; El-Twab, A.; Sanaa, M.; Abdel Reheim, E.S.; Ashour, M.B. Gallic acid and p-coumaric acid attenuate type 2 diabetes-induced neurodegeneration in rats. Metab. Brain Dis. 2017, 32, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Wu, S.; Li, B.; Tan, J.; Yan, J.; Wang, Y.; Tang, Z.; Liu, M.; Fu, C.; Zhang, H.; et al. Dietary ferulic acid and vanillic acid on inflammation, gut barrier function and growth performance in lipopolysaccharide-challenged piglets. Anim. Nutr. 2022, 8, 144–152. [Google Scholar] [CrossRef]
- Ziadlou, R.; Barbero, A.; Martin, I.; Wang, X.; Qin, L.; Alini, M.; Grad, S. Anti-inflammatory and chondroprotective effects of vanillic acid and epimedin C in human osteoarthritic chondrocytes. Biomolecules 2020, 10, 932. [Google Scholar] [CrossRef]
- El-Masry, T.; Altwaijry, N.; Alotaibi, B.; Tousson, E.; Alboghdadly, A.; Saleh, A. Chicory (Cichorium intybus L.) extract ameliorates hydroxyapatite nanoparticles induced kidney damage in rats. Pak. J. Pharm. Sci. 2020, 33, 1251–1260. [Google Scholar]
- Erkec, O.E.; Arihan, O.; Colcimen, N.; Kara, M.; Karatas, E.; Demir, H.; Ragbetli, M.C. Effects of Cichorium intybus on serum oxidative stress, liver and kidney volume, and cyclin B1 and Bcl-2 levels in the brains of rats with ethanol induced damage. Cell. Mol. Biol. 2018, 64, 30–35. [Google Scholar] [CrossRef]
- Parvez, M.K.; Al-Dosari, M.S.; Tabish Rehman, M.; Alajmi, M.F.; Alqahtani, A.S.; AlSaid, M.S. New terpenic and phenolic compounds from Suaeda monoica reverse oxidative and apoptotic damages in human endothelial cells. Saudi Pharm. J. 2021, 29, 1102–1111. [Google Scholar] [CrossRef]
- Jazvinšćak Jembrek, M.; Oršolić, N.; Mandić, L.; Sadžak, A.; Šegota, S. Anti-Oxidative, Anti-Inflammatory and Anti-Apoptotic Effects of Flavonols: Targeting Nrf2, NF-κB and p53 Pathways in Neurodegeneration. Antioxidants 2021, 10, 1628. [Google Scholar] [CrossRef]
- Prince, P.; Dhanasekar, K.; Rajakumar, S. Preventive effects of vanillic acid on lipids, bax, bcl-2 and myocardial infarct size on isoproterenol-induced myocardial infarcted rats: A biochemical and in vitro study. Cardiovasc. Toxicol. 2011, 11, 58–66. [Google Scholar] [CrossRef]
- Ding, S.; Liu, D.; Wang, L.; Wang, G.; Zhu, Y. Inhibiting microRNA-29a protects myocardial ischemia-reperfusion injury by targeting SIRT1 and suppressing oxidative stress and NLRP3-mediated pyroptosis pathway. J. Pharmacol. Exp. Ther. 2020, 372, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Li, X.; Ren, Y.; Wang, J.; Xu, D.; Hang, Y.; Zhou, T.; Wang, L. MicroRNA-29a regulates lipopolysaccharide (LPS)-induced inflammatory responses in murine macrophages through the Akt1/NF-κB pathway. Exp. Cell Res. 2017, 360, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Ma, L.; Liu, Z.; Yang, Z.; Wang, J.; Liu, J.; Jiang, G. Targeting microRNAs: A new action mechanism of natural compounds. Oncotarget 2017, 8, 15961–15970. [Google Scholar] [CrossRef] [PubMed]
- Biersack, B. Current state of phenolic and terpenoidal dietary factors and natural products as non-coding RNA/microRNA modulators for improved cancer therapy and prevention. Non-Coding RNA Res. 2016, 1, 12–34. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primer | Reverse Primer | Annealing Temperature | |
|---|---|---|---|
| NF-κb | 5′-CAATGGCTACACAGGACCA-3′ | 5′-CACTGTCACCTGGAACCAGA-3′ | 52 °C |
| TNF-α | 5′-TCTACTGAACTTCGGGGTGATCG-3′ | 5′-TGATCTGAGTGTGAGGGTCTGGG-3′ | 56 °C |
| p53 | 5′-ACCGCCGACCTATCCTTACC-3′ | 5′-TCTTCTGTACGGCGGTCTCTC-3′ | 56 °C |
| β-actin | 5′-ACGGCCAGGTCATCACTATTG-3′ | 5′-CAAGAAGGAAGGCTGGAAAAGA-3′ | 52 °C |
| miR-29a | 5′-GCGCACTGATTTCTTTTGGTGTTCAG-3′ | 5′-GCGAGCACAGAATTAATACGAC-3′ | 51 °C |
| RNU6B | 5′-CTCGCTTCGGCAGCACATA-3′ | 5′-CGCTTCACGAATTTGCGTG-3′ | 53 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eltamany, E.E.; Mosalam, E.M.; Mehanna, E.T.; Awad, B.M.; Mosaad, S.M.; Abdel-Kader, M.S.; Ibrahim, A.K.; Badr, J.M.; Goda, M.S. Potential Gonado-Protective Effect of Cichorium endivia and Its Major Phenolic Acids against Methotrexate-Induced Testicular Injury in Mice. Biomedicines 2022, 10, 1986. https://doi.org/10.3390/biomedicines10081986
Eltamany EE, Mosalam EM, Mehanna ET, Awad BM, Mosaad SM, Abdel-Kader MS, Ibrahim AK, Badr JM, Goda MS. Potential Gonado-Protective Effect of Cichorium endivia and Its Major Phenolic Acids against Methotrexate-Induced Testicular Injury in Mice. Biomedicines. 2022; 10(8):1986. https://doi.org/10.3390/biomedicines10081986
Chicago/Turabian StyleEltamany, Enas E., Esraa M. Mosalam, Eman T. Mehanna, Basma M. Awad, Sarah M. Mosaad, Maged S. Abdel-Kader, Amany K. Ibrahim, Jihan M. Badr, and Marwa S. Goda. 2022. "Potential Gonado-Protective Effect of Cichorium endivia and Its Major Phenolic Acids against Methotrexate-Induced Testicular Injury in Mice" Biomedicines 10, no. 8: 1986. https://doi.org/10.3390/biomedicines10081986