
Murine and Human Gastric Tissue Establishes Organoids after 48 Hours of Cold Ischemia Time during Shipment

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
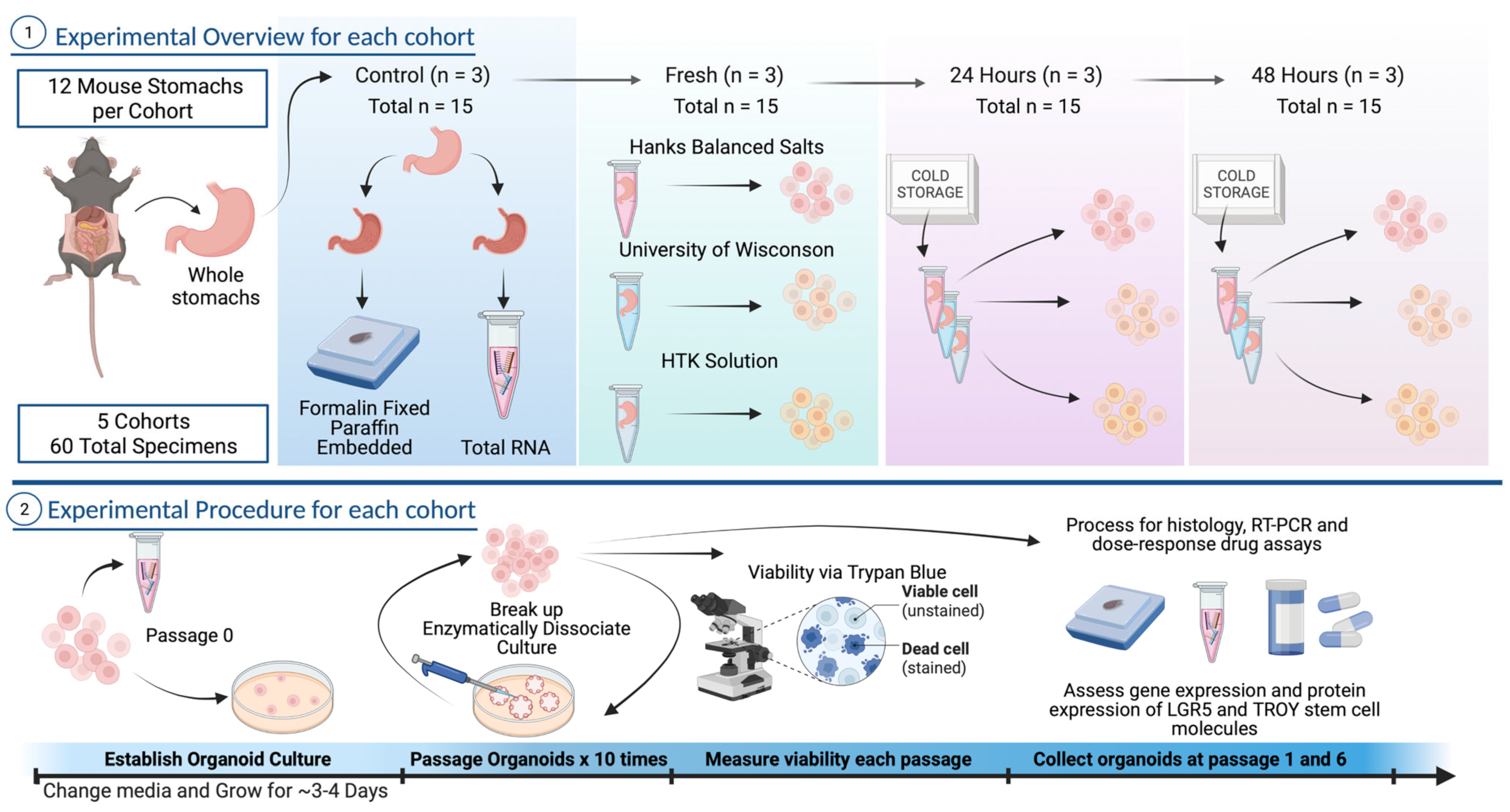
2.1. Study Design
2.2. Mouse and Human Specimens
2.3. Organoid Culture
2.4. Immunofluorescence and Immunohistochemistry
2.5. qRT-PCR
2.6. In Vitro Dose–Response Assay
2.7. Statistical Analysis
3. Results
3.1. Gastric Organoids Recapitulate Murine Gastric Tissue and Morphology
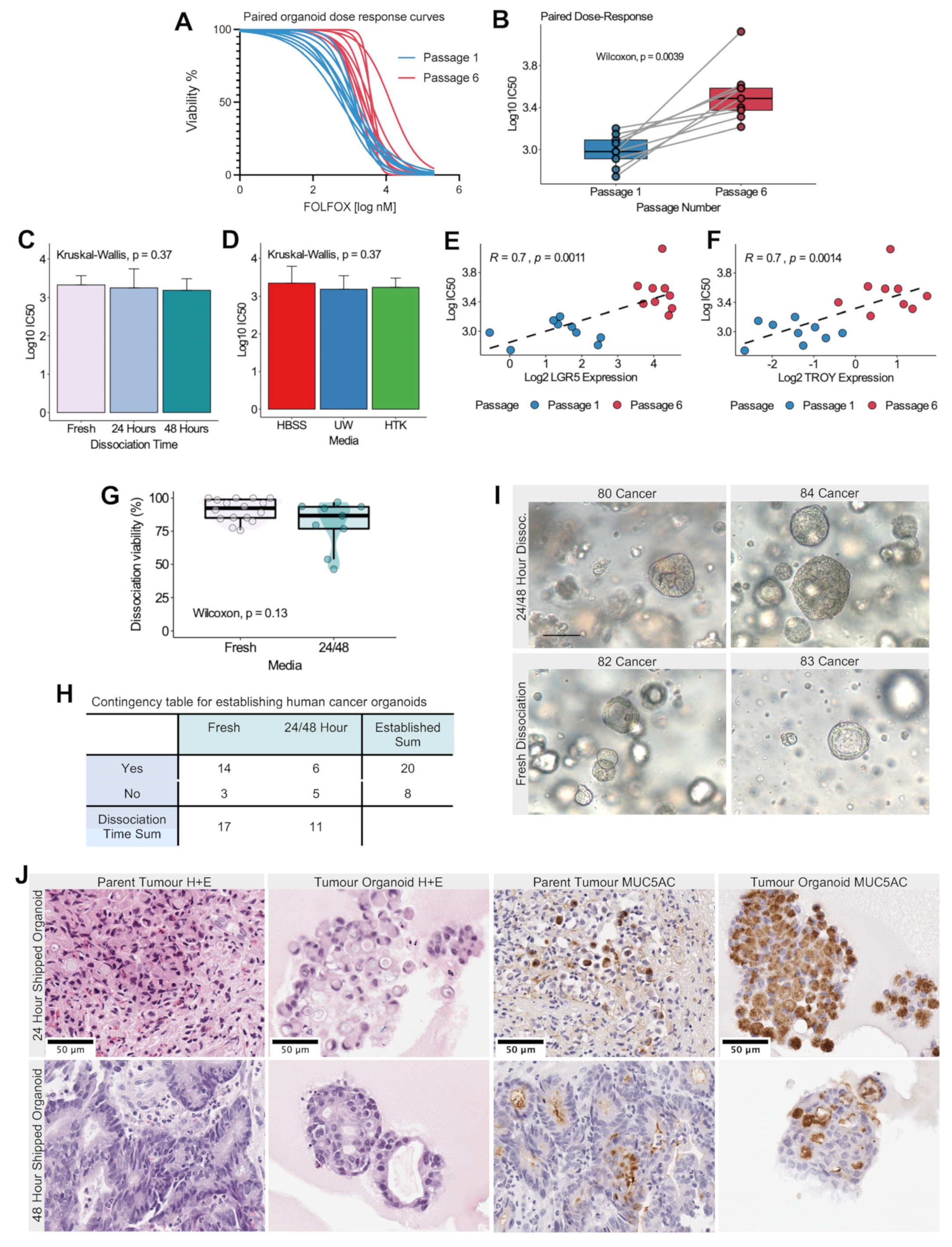
3.2. Dissociation Viability Is Affected by Storage Medium and Is Cohort-Dependent
3.3. Long-Term Organoid Viability Is not Affected by Storage Medium and Dissociation Time
3.4. Cold Storage for 24 or 48 Hours Decreases Organoid Growth Rate
3.5. In Vitro Gene Expression of Gastric Stem Cell Markers LGR5 and TROY Re-Establishes Baseline Endogenous Levels after 6 Passages and Is Unaffected by Cold Storage Conditions
3.6. In Vitro Organoid Dose Response to Cytotoxic Therapy Is Significantly Associated with Passage Number and Stem Cell Gene Expression
3.7. Human Tumor Organoids can Be Established following 24- and 48-Hour Cold Storage in HBSS Solution
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, J.; Koo, B.K.; Knoblich, J.A. Human Organoids: Model Systems for Human Biology and Medicine. Nat. Rev. Mol. Cell Biol. 2020, 21, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Aberle, M.R.; Burkhart, R.A.; Tiriac, H.; Olde Damink, S.W.M.; Dejong, C.H.C.; Tuveson, D.A.; van Dam, R.M. Patient-Derived Organoid Models Help Define Personalized Management of Gastrointestinal Cancer. Br. J. Surg. 2018, 105, e48–e60. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, K.; Wu, C.; O’Rourke, K.P.; Szeglin, B.C.; Zheng, Y.; Sauvé, C.E.G.; Adileh, M.; Wasserman, I.; Marco, M.R.; Kim, A.S.; et al. A Rectal Cancer Organoid Platform to Study Individual Responses to Chemoradiation. Nat. Med. 2019, 25, 1607–1614. [Google Scholar] [CrossRef]
- Foo, M.A.; You, M.; Chan, S.L.; Sethi, G.; Bonney, G.K.; Yong, W.P.; Chow, E.K.H.; Fong, E.L.S.; Wang, L.; Goh, B.C. Clinical Translation of Patient-Derived Tumour Organoids- Bottlenecks and Strategies. Biomark Res. 2022, 10, 10. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Lin, M.; Rao, M.; Thompson, H.; Hirai, K.; Choi, M.; Georgakis, G.V.; Sasson, A.R.; Bucobo, J.C.; Tzimas, D.; et al. Development of Patient-Derived Gastric Cancer Organoids from Endoscopic Biopsies and Surgical Tissues. Ann. Surg. Oncol. 2018, 25, 2767–2775. [Google Scholar] [CrossRef]
- Ooft, S.N.; Weeber, F.; Schipper, L.; Dijkstra, K.K.; McLean, C.M.; Kaing, S.; van de Haar, J.; Prevoo, W.; van Werkhoven, E.; Snaebjornsson, P.; et al. Prospective Experimental Treatment of Colorectal Cancer Patients Based on Organoid Drug Responses. ESMO Open 2021, 6, 100103. [Google Scholar] [CrossRef]
- Fujii, M.; Shimokawa, M.; Date, S.; Takano, A.; Matano, M.; Nanki, K.; Ohta, Y.; Toshimitsu, K.; Nakazato, Y.; Kawasaki, K.; et al. A Colorectal Tumor Organoid Library Demonstrates Progressive Loss of Niche Factor Requirements during Tumorigenesis. Cell Stem Cell 2016, 18, 827–838. [Google Scholar] [CrossRef] [Green Version]
- Puca, L.; Bareja, R.; Prandi, D.; Shaw, R.; Benelli, M.; Karthaus, W.R.; Hess, J.; Sigouros, M.; Donoghue, A.; Kossai, M.; et al. Patient Derived Organoids to Model Rare Prostate Cancer Phenotypes. Nat. Commun. 2018, 9, 2404. [Google Scholar] [CrossRef] [Green Version]
- Mowat, C.; Mosley, S.R.; Namdar, A.; Schiller, D.; Baker, K. Anti-Tumor Immunity in Mismatch Repair-Deficient Colorectal Cancers Requires Type I IFN–Driven CCL5 and CXCL10. J. Exp. Med. 2021, 218, e20210108. [Google Scholar] [CrossRef]
- Bartfeld, S.; Bayram, T.; van de Wetering, M.; Huch, M.; Begthel, H.; Kujala, P.; Vries, R.; Peters, P.J.; Clevers, H. In Vitro Expansion of Human Gastric Epithelial Stem Cells and Their Responses to Bacterial Infection. Gastroenterology 2015, 148, 126.e6–136.e6. [Google Scholar] [CrossRef]
- Miyoshi, H.; Stappenbeck, T.S. In Vitro Expansion and Genetic Modification of Gastrointestinal Stem Cells in Spheroid Culture. Nat. Protoc. 2013, 8, 2471–2482. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Biosystems, A. Applied Biosystems Guide to Performing Relative Quantitation of Gene Expression Using Real-Time Quantitative PCR. Appl. Biosyst. 2008, 2009, 1–60. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. In R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Corfield, A.P.; Myerscough, N.; Longman, R.; Sylvester, P.; Arul, S.; Pignatelli, M. Mucins and Mucosal Protection in the Gastrointestinal Tract: New Prospects for Mucins in the Pathology of Gastrointestinal Disease. Gut 2000, 47, 589 LP–594 LP. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, J.-Y.; Brezden-Masley, C.; Streutker, C.J. Lgr5 and Stem/Progenitor Gene Expression in Gastric/Gastroesophageal Junction Carcinoma—Significance of Potentially Retained Stemness. BMC Cancer 2020, 20, 860. [Google Scholar] [CrossRef] [PubMed]
- Evelina, S.; Wen, Z.; Linn, F.; Max, K.; Nicholas, M.; Csaba, A.; Per, O.; Fredrik, E.; Agnieszka, L.; Feria, H.; et al. An Atlas of the Protein-Coding Genes in the Human, Pig, and Mouse Brain. Science (1979) 2020, 367, eaay5947. [Google Scholar] [CrossRef]
- Pauli, C.; Hopkins, B.D.; Prandi, D.; Shaw, R.; Fedrizzi, T.; Sboner, A.; Sailer, V.; Augello, M.; Puca, L.; Rosati, R.; et al. Personalized in vitro and in vivo Cancer Models to Guide Precision Medicine. Cancer Discov. 2017, 7, 462–477. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.H.N.; Siu, H.C.; Law, S.; Ho, S.L.; Yue, S.S.K.; Tsui, W.Y.; Chan, D.; Chan, A.S.; Ma, S.; Lam, K.O.; et al. A Comprehensive Human Gastric Cancer Organoid Biobank Captures Tumor Subtype Heterogeneity and Enables Therapeutic Screening. Cell Stem Cell 2018, 23, 882.e11–897.e11. [Google Scholar] [CrossRef] [Green Version]
- van de Wetering, M.; Francies, H.E.; Francis, J.M.; Bounova, G.; Iorio, F.; Pronk, A.; Van Houdt, W.; Van Gorp, J.; Taylor-Weiner, A.; Kester, L.; et al. Prospective Derivation of a Living Organoid Biobank of Colorectal Cancer Patients. Cell 2015, 161, 933–945. [Google Scholar] [CrossRef] [Green Version]
- Tiriac, H.; Belleau, P.; Engle, D.D.; Plenker, D.; Deschênes, A.; Somerville, T.D.D.; Froeling, F.E.M.; Burkhart, R.A.; Denroche, R.E.; Jang, G.H.; et al. Organoid Profiling Identifies Common Responders to Chemotherapy in Pancreatic Cancer. Cancer Discov. 2018, 8, 1112–1129. [Google Scholar] [CrossRef] [Green Version]
- Vlachogiannis, G.; Hedayat, S.; Vatsiou, A.; Jamin, Y.; Fernández-Mateos, J.; Khan, K.; Lampis, A.; Eason, K.; Huntingford, I.; Burke, R.; et al. Patient-Derived Organoids Model Treatment Response of Metastatic Gastrointestinal Cancers. Science (1979) 2018, 359, 920–926. [Google Scholar] [CrossRef]
- Walsh, A.J.; Cook, R.S.; Sanders, M.E.; Arteaga, C.L.; Skala, M.C. Drug Response in Organoids Generated from Frozen Primary Tumor Tissues. Sci. Rep. 2016, 6, 18889. [Google Scholar] [CrossRef] [Green Version]
- Georgiou, M.; Chichagova, V.; Hilgen, G.; Dorgau, B.; Sernagor, E.; Armstrong, L.; Lako, M. Room Temperature Shipment Does Not Affect the Biological Activity of Pluripotent Stem Cell-Derived Retinal Organoids. PLoSOne 2020, 15, e0233860. [Google Scholar] [CrossRef]
- Singh, R.K.; Winkler, P.; Binette, F.; Glickman, R.D.; Seiler, M.; Petersen-Jones, S.M.; Nasonkin, I.O. Development of a Protocol for Maintaining Viability While Shipping Organoid-Derived Retinal Tissue. J. Tissue Eng. Regen. Med. 2020, 14, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Lin, W.; Wen, L.; Li, G. Lgr5 in Cancer Biology: Functional Identification of Lgr5 in Cancer Progression and Potential Opportunities for Novel Therapy. Stem Cell Res. Ther. 2019, 10, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hata, M.; Hayakawa, Y.; Koike, K. Gastric Stem Cell and Cellular Origin of Cancer. Biomedicines 2018, 6, 100. [Google Scholar] [CrossRef] [Green Version]
- Xi, H.-Q.; Cui, J.-X.; Shen, W.-S.; Wu, X.-S.; Bian, S.-B.; Li, J.-Y.; Song, Z.; Wei, B.; Chen, L. Increased Expression of Lgr5 Is Associated with Chemotherapy Resistance in Human Gastric Cancer. Oncol. Rep. 2014, 32, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Phipson, B.; Er, P.X.; Combes, A.N.; Forbes, T.A.; Howden, S.E.; Zappia, L.; Yen, H.J.; Lawlor, K.T.; Hale, L.J.; Sun, J.; et al. Evaluation of Variability in Human Kidney Organoids. Nat. Methods 2019, 16, 79–87. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Covariate | Beta | 95% CI 1 | p-Value |
|---|---|---|---|
| Cohort | |||
| 1 | — | — | |
| 2 | −11 | −24, 1.2 | 0.075 |
| 3 | −14 | −32, 4.4 | 0.13 |
| 4 | −10 | −19, −0.72 | 0.036 |
| 5 | −16 | −26, −5.9 | 0.003 |
| Age of Mouse | −0.18 | −0.45, 0.10 | 0.2 |
| Medium | |||
| HBSS | — | — | |
| UW | −2.8 | −9.8, 4.2 | 0.4 |
| HTK | −14 | −22, −6.8 | <0.001 |
| Dissociation Time | |||
| Fresh | — | — | |
| 24 Hours | −4.7 | −12, 2.5 | 0.2 |
| 48 Hours | −6.8 | −14, 0.18 | 0.056 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skubleny, D.; Garg, S.; Wickware, J.; Purich, K.; Ghosh, S.; Spratlin, J.; Schiller, D.E.; Rayat, G.R. Murine and Human Gastric Tissue Establishes Organoids after 48 Hours of Cold Ischemia Time during Shipment. Biomedicines 2023, 11, 151. https://doi.org/10.3390/biomedicines11010151
Skubleny D, Garg S, Wickware J, Purich K, Ghosh S, Spratlin J, Schiller DE, Rayat GR. Murine and Human Gastric Tissue Establishes Organoids after 48 Hours of Cold Ischemia Time during Shipment. Biomedicines. 2023; 11(1):151. https://doi.org/10.3390/biomedicines11010151
Chicago/Turabian StyleSkubleny, Daniel, Saurabh Garg, Jim Wickware, Kieran Purich, Sunita Ghosh, Jennifer Spratlin, Dan E. Schiller, and Gina R. Rayat. 2023. "Murine and Human Gastric Tissue Establishes Organoids after 48 Hours of Cold Ischemia Time during Shipment" Biomedicines 11, no. 1: 151. https://doi.org/10.3390/biomedicines11010151