
GM1 Oligosaccharide Efficacy in Parkinson’s Disease: Protection against MPTP

 ,
,  , , ,
, , ,  , , , ,
, , , ,  , and
, and 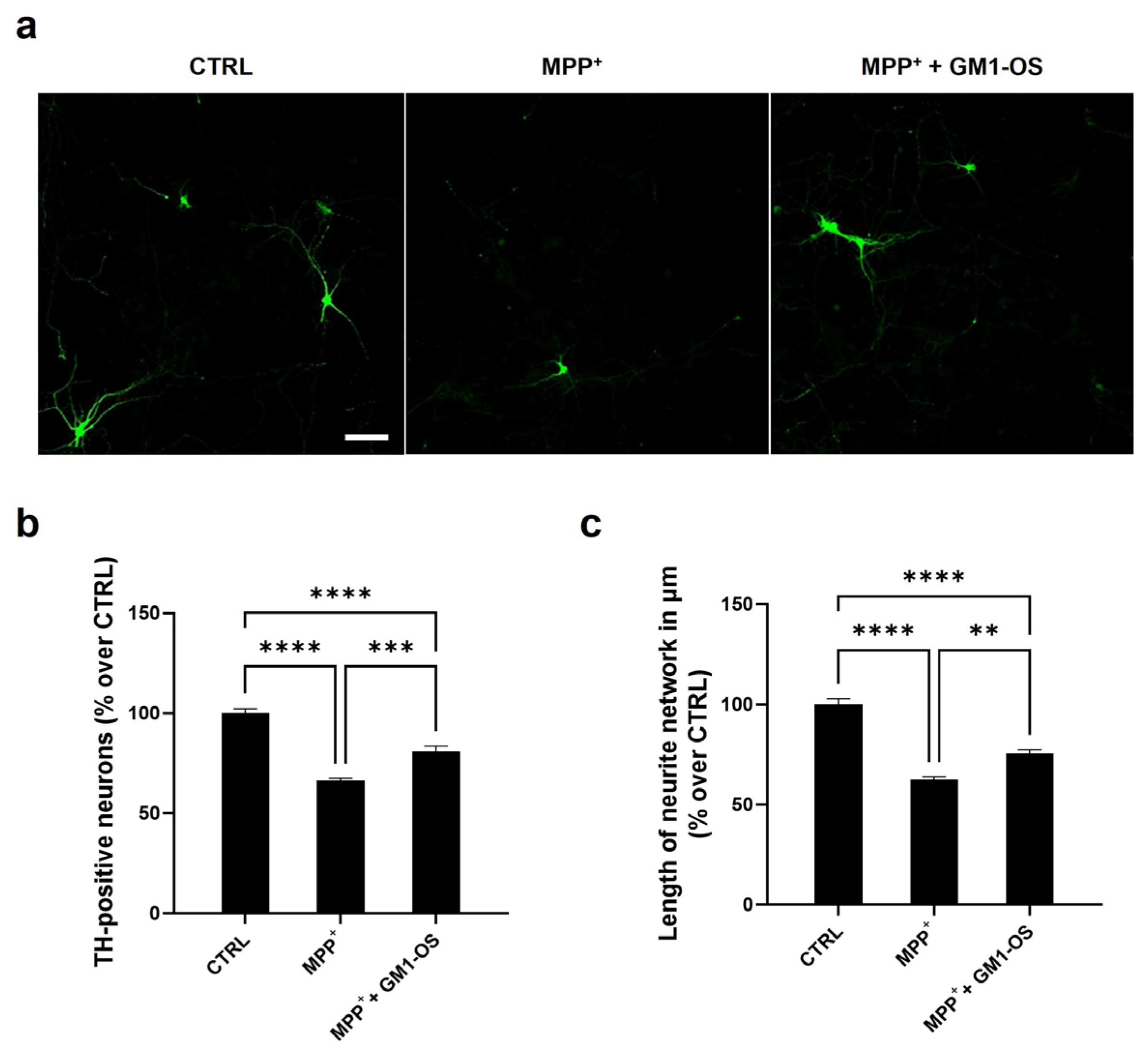{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
Antibodies
2.2. Primary Cultures
2.2.1. Primary Culture of DA Neurons
2.2.2. Primary Culture of CGNs
2.3. Cell Treatments
2.4. Cell Immunostaining
2.5. MitoSOX Red Staining
2.6. Morphological Analysis
2.7. MTT Assay
2.8. LDH Assay
2.9. WB Analysis
2.10. GM1-OS Preparation
2.11. Protein Determination
2.12. Statistical Analysis
3. Results
3.1. GM1-OS Protection from MPTP in DA Neurons
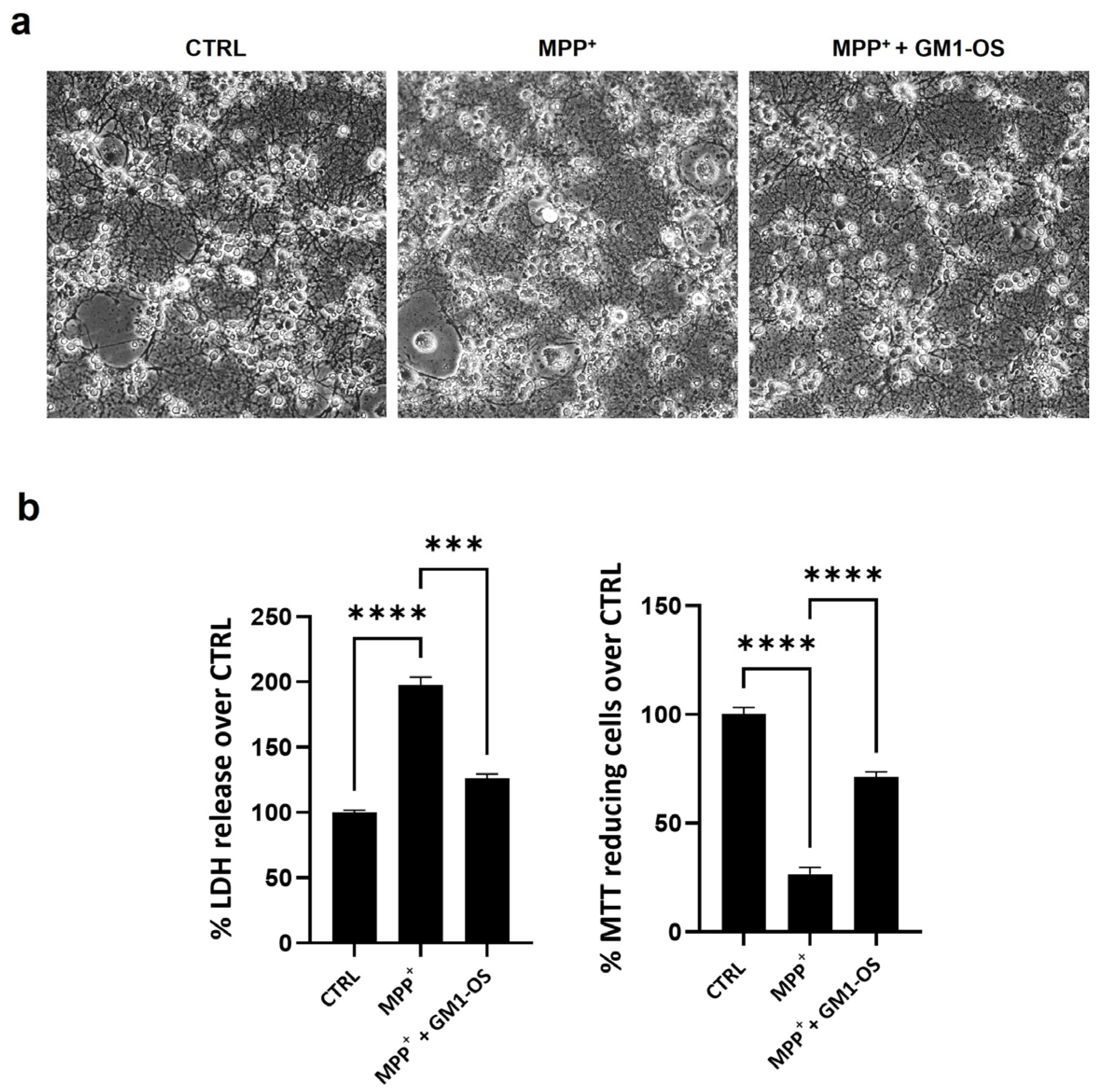
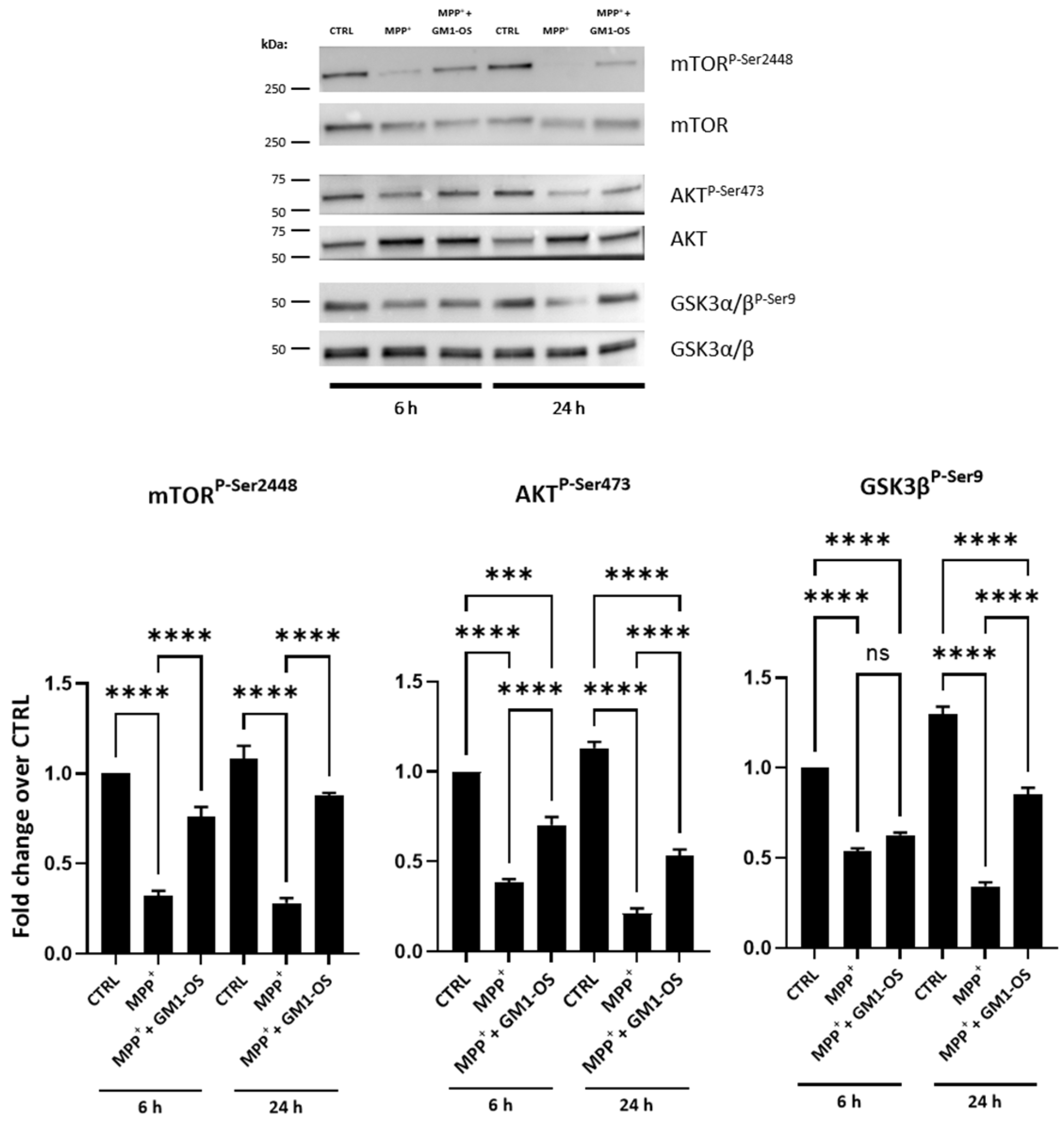
3.2. GM1-OS Protection from MPTP in CGNs
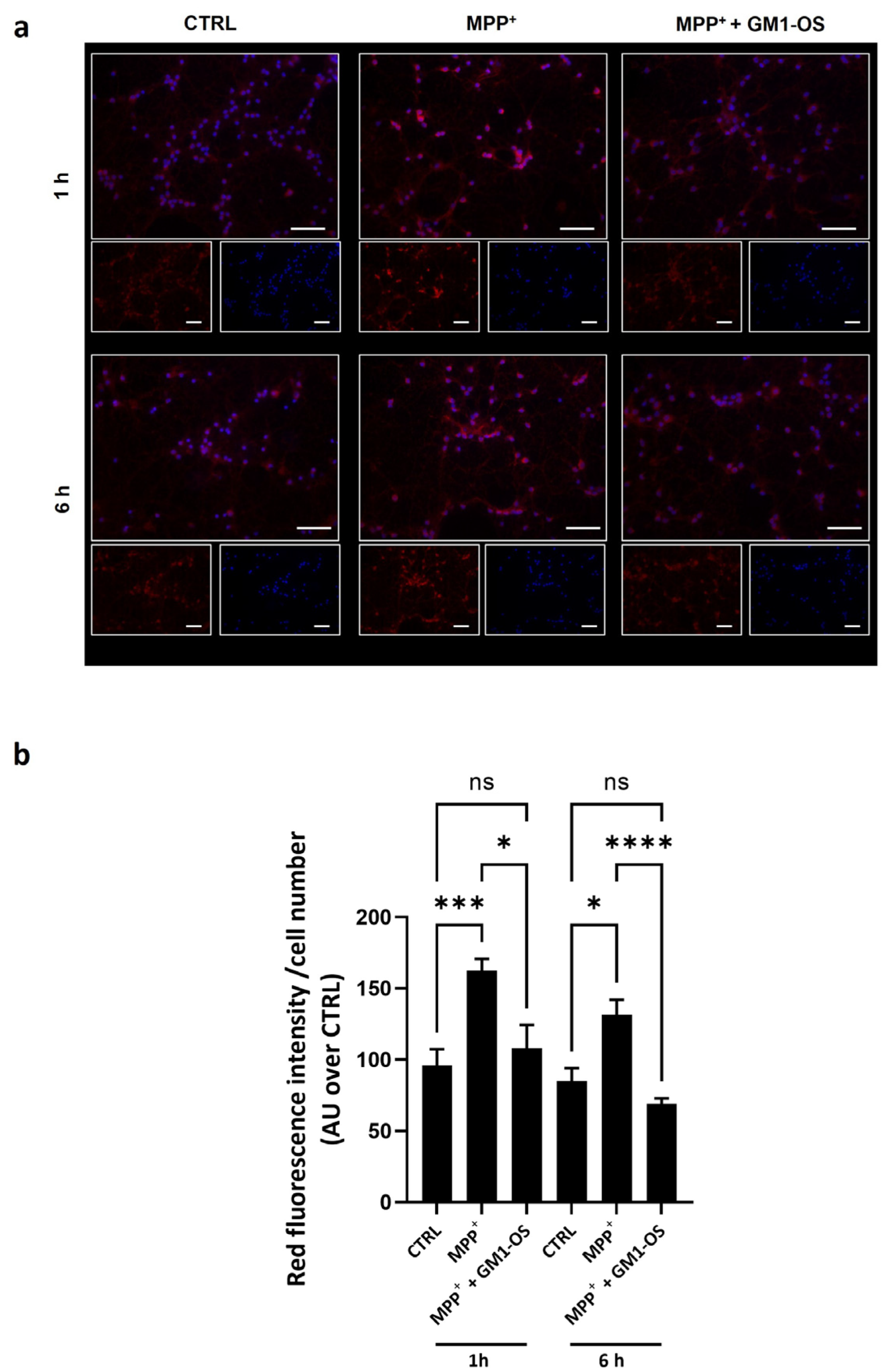
3.3. GM1-OS Lowered the Excess of Mitochondrial Superoxide (O2•−) Induced by MPTP
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Ganglioside nomenclature is in accordance with IUPAC-IUBB recommendations [58] | |
| αS | alpha-synuclein |
| Akt | RAC-alpha serine/threonine-protein kinase |
| BBB | Blood–brain barrier |
| BSA | bovine serum albumin |
| BDNF | brain-derived neurotrophic factor |
| CGN | cerebellar granule neurons |
| CMF-PBS | calcium magnesium free-PBS |
| CNS | central nervous system |
| CTRL | control |
| DA | dopamine |
| DAPI | 4′,5-diamina-2-phenylindole |
| DAT | DA transporter |
| DMEM | Dulbecco’s modified Eagles’ medium |
| EDTA | ethylenediaminetetraacetic acid |
| FBS | fetal bovine serum |
| FCS | fetal calf serum |
| GDNF | glial-derived neurotrophic factor |
| GM1 | II3Neu5Ac-Gg4Cer, β-Gal-(1-3)-β-GalNAc-(1-4)-[α-Neu5Ac-(2-3)]-β-Gal-(1-4)-β-Glc-Cer |
| GM1-OS | GM1-oligosaccharide, II3Neu5Ac-Gg4, β-Gal-(1-3)-β-GalNAc-(1-4)-[α-Neu5Ac-(2-3)]-β-Gal-(1-4)-Glc |
| GSK-3β | glycogen synthase kinase-3β |
| HBSS | Hanks’ balanced salt solution |
| HPR | horseradish peroxidase |
| IHC | immunohistochemistry |
| IL-6 | interleukin 6 |
| IP | Protease inhibitor cocktail |
| LDH | lactate dehydrogenase |
| MAPK | mitogen-activated protein kinase |
| MPTP | 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine hydrocholoride |
| MPP+ | 1-methyl-4-phenylpyridinium |
| mTORs | mammalian target of rapamycin |
| MTT | 3-(4,5,-dimethylthiazole-2yl)-2,5-diphenyltetrazolium bromide |
| Na3VO4 | sodium orthovanadate |
| O2•- | superoxide |
| p-AKT | phospho-AKT |
| p-GSK-3β | phospho-GSK-3β |
| p-mTOR | phospho-mTOR |
| PBS | phosphate-buffered saline |
| PFA | paraformaldehyde |
| PMSF | phenylmethanesulfonyl fluoride |
| PD | Parkinson’s disease |
| P/S | penicillin and streptomycin |
| PVDF | polyvinylidene difluoride |
| ROS | reactive oxygen species |
| RRID | research resource identifier |
| SDS | sodium dodecyl sulfate |
| SEM | standard error of the mean |
| Ser | Serine residue |
| SNpc | substantia nigra pars compacta |
| TH | tyrosine hydroxylase |
| TrkA | neurotrophin tyrosin kinase receptor |
| WB | western blot |
References
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Primers 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed]
- Del Rey, N.L.; Quiroga-Varela, A.; Garbayo, E.; Carballo-Carbajal, I.; Fernandez-Santiago, R.; Monje, M.H.G.; Trigo-Damas, I.; Blanco-Prieto, M.J.; Blesa, J. Advances in Parkinson’s Disease: 200 Years Later. Front. Neuroanat. 2018, 12, 113. [Google Scholar] [CrossRef] [PubMed]
- Toomey, C.E.; Heywood, W.E.; Evans, J.R.; Lachica, J.; Pressey, S.N.; Foti, S.C.; Al Shahrani, M.; D’Sa, K.; Hargreaves, I.P.; Heales, S.; et al. Mitochondrial dysfunction is a key pathological driver of early stage Parkinson’s. Acta Neuropathol. Commun. 2022, 10, 134. [Google Scholar] [CrossRef]
- Gonzalez-Rodriguez, P.; Zampese, E.; Stout, K.A.; Guzman, J.N.; Ilijic, E.; Yang, B.; Tkatch, T.; Stavarache, M.A.; Wokosin, D.L.; Gao, L.; et al. Disruption of mitochondrial complex I induces progressive parkinsonism. Nature 2021, 599, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Lang, M.; Pramstaller, P.P.; Pichler, I. Crosstalk of organelles in Parkinson’s disease - MiT family transcription factors as central players in signaling pathways connecting mitochondria and lysosomes. Mol. Neurodegener. 2022, 17, 50. [Google Scholar] [CrossRef]
- Fanning, S.; Selkoe, D.; Dettmer, U. Parkinson’s disease: Proteinopathy or lipidopathy? NPJ Parkinsons. Dis. 2020, 6, 3. [Google Scholar] [CrossRef]
- Ledeen, R.W.; Wu, G. The multi-tasked life of GM1 ganglioside, a true factotum of nature. Trends Biochem. Sci. 2015, 40, 407–418. [Google Scholar] [CrossRef]
- Chowdhury, S.; Wu, G.; Lu, Z.H.; Kumar, R.; Ledeen, R. Age-Related Decline in Gangliosides GM1 and GD1a in Non-CNS Tissues of Normal Mice: Implications for Peripheral Symptoms of Parkinson’s Disease. Biomedicines 2023, 11, 209. [Google Scholar] [CrossRef]
- Ledeen, R.W.; Wu, G. Gangliosides, alpha-Synuclein, and Parkinson’s Disease. Prog. Mol. Biol. Transl. Sci. 2018, 156, 435–454. [Google Scholar] [CrossRef]
- Ledeen, R.; Wu, G. Gangliosides of the Nervous System. Methods Mol. Biol. 2018, 1804, 19–55. [Google Scholar] [CrossRef]
- Ledeen, R.; Chowdhury, S.; Lu, Z.H.; Chakraborty, M.; Wu, G. Systemic deficiency of GM1 ganglioside in Parkinson’s disease tissues and its relation to the disease etiology. Glycoconj. J. 2022, 39, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Lu, Z.H.; Kulkarni, N.; Ledeen, R.W. Deficiency of ganglioside GM1 correlates with Parkinson’s disease in mice and humans. J. Neurosci. Res. 2012, 90, 1997–2008. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.S. Altered expression of genes involved in ganglioside biosynthesis in substantia nigra neurons in Parkinson’s disease. PLoS ONE 2018, 13, e0199189. [Google Scholar] [CrossRef] [PubMed]
- Hadaczek, P.; Wu, G.; Sharma, N.; Ciesielska, A.; Bankiewicz, K.; Davidow, A.L.; Lu, Z.H.; Forsayeth, J.; Ledeen, R.W. GDNF signaling implemented by GM1 ganglioside; failure in Parkinson’s disease and GM1-deficient murine model. Exp. Neurol. 2015, 263, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Lu, Z.H.; Kulkarni, N.; Amin, R.; Ledeen, R.W. Mice lacking major brain gangliosides develop parkinsonism. Neurochem. Res. 2011, 36, 1706–1714. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Lu, Z.H.; Seo, J.H.; Alselehdar, S.K.; DeFrees, S.; Ledeen, R.W. Mice deficient in GM1 manifest both motor and non-motor symptoms of Parkinson’s disease; successful treatment with synthetic GM1 ganglioside. Exp. Neurol. 2020, 329, 113284. [Google Scholar] [CrossRef]
- Fazzari, M.; Lunghi, G.; Chiricozzi, E.; Mauri, L.; Sonnino, S. Gangliosides and the Treatment of Neurodegenerative Diseases: A Long Italian Tradition. Biomedicines 2022, 10, 363. [Google Scholar] [CrossRef]
- Chiricozzi, E.; Lunghi, G.; Di Biase, E.; Fazzari, M.; Sonnino, S.; Mauri, L. GM1 Ganglioside Is A Key Factor in Maintaining the Mammalian Neuronal Functions Avoiding Neurodegeneration. Int. J. Mol. Sci. 2020, 21, 868. [Google Scholar] [CrossRef]
- Magistretti, P.J.; Geisler, F.H.; Schneider, J.S.; Li, P.A.; Fiumelli, H.; Sipione, S. Gangliosides: Treatment Avenues in Neurodegenerative Disease. Front. Neurol. 2019, 10, 859. [Google Scholar] [CrossRef]
- Chiricozzi, E.; Di Biase, E.; Lunghi, G.; Fazzari, M.; Loberto, N.; Aureli, M.; Mauri, L.; Sonnino, S. Turning the spotlight on the oligosaccharide chain of GM1 ganglioside. Glycoconj. J. 2021, 38, 101–117. [Google Scholar] [CrossRef]
- Chiricozzi, E.; Mauri, L.; Lunghi, G.; Di Biase, E.; Fazzari, M.; Maggioni, M.; Valsecchi, M.; Prioni, S.; Loberto, N.; Pome, D.Y.; et al. Parkinson’s disease recovery by GM1 oligosaccharide treatment in the B4galnt1(+/−) mouse model. Sci. Rep. 2019, 9, 19330. [Google Scholar] [CrossRef] [PubMed]
- Nonnekes, J.; Post, B.; Tetrud, J.W.; Langston, J.W.; Bloem, B.R. MPTP-induced parkinsonism: An historical case series. Lancet Neurol. 2018, 17, 300–301. [Google Scholar] [CrossRef] [PubMed]
- Mustapha, M.; Mat Taib, C.N. MPTP-induced mouse model of Parkinson’s disease: A promising direction of therapeutic strategies. Bosn. J. Basic Med. Sci. 2021, 21, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Callizot, N.; Combes, M.; Henriques, A.; Poindron, P. Necrosis, apoptosis, necroptosis, three modes of action of dopaminergic neuron neurotoxins. PLoS ONE 2019, 14, e0215277. [Google Scholar] [CrossRef]
- Visanji, N.P.; Orsi, A.; Johnston, T.H.; Howson, P.A.; Dixon, K.; Callizot, N.; Brotchie, J.M.; Rees, D.D. PYM50028, a novel, orally active, nonpeptide neurotrophic factor inducer, prevents and reverses neuronal damage induced by MPP+ in mesencephalic neurons and by MPTP in a mouse model of Parkinson’s disease. FASEB J. 2008, 22, 2488–2497. [Google Scholar] [CrossRef]
- Langston, J.W.; Irwin, I.; Langston, E.B.; Forno, L.S. 1-Methyl-4-phenylpyridinium ion (MPP+): Identification of a metabolite of MPTP, a toxin selective to the substantia nigra. Neurosci. Lett. 1984, 48, 87–92. [Google Scholar] [CrossRef]
- Perry, T.L.; Yong, V.W.; Jones, K.; Wall, R.A.; Clavier, R.M.; Foulks, J.G.; Wright, J.M. Effects of N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine and its metabolite, N-methyl-4-phenylpyridinium ion, on dopaminergic nigrostriatal neurons in the mouse. Neurosci. Lett. 1985, 58, 321–326. [Google Scholar] [CrossRef]
- Zaltieri, M.; Longhena, F.; Pizzi, M.; Missale, C.; Spano, P.; Bellucci, A. Mitochondrial Dysfunction and alpha-Synuclein Synaptic Pathology in Parkinson’s Disease: Who’s on First? Parkinsons. Dis. 2015, 2015, 108029. [Google Scholar] [CrossRef]
- Plun-Favreau, H.; Klupsch, K.; Moisoi, N.; Gandhi, S.; Kjaer, S.; Frith, D.; Harvey, K.; Deas, E.; Harvey, R.J.; McDonald, N.; et al. The mitochondrial protease HtrA2 is regulated by Parkinson’s disease-associated kinase PINK1. Nat. Cell Biol. 2007, 9, 1243–1252. [Google Scholar] [CrossRef]
- Meredith, G.E.; Rademacher, D.J. MPTP mouse models of Parkinson’s disease: An update. J. Parkinsons. Dis. 2011, 1, 19–33. [Google Scholar] [CrossRef]
- Schneider, J.S.; Seyfried, T.N.; Choi, H.S.; Kidd, S.K. Intraventricular Sialidase Administration Enhances GM1 Ganglioside Expression and Is Partially Neuroprotective in a Mouse Model of Parkinson’s Disease. PLoS ONE 2015, 10, e0143351. [Google Scholar] [CrossRef]
- Chiricozzi, E.; Maggioni, M.; di Biase, E.; Lunghi, G.; Fazzari, M.; Loberto, N.; Elisa, M.; Scalvini, F.G.; Tedeschi, G.; Sonnino, S. The Neuroprotective Role of the GM1 Oligosaccharide, II(3)Neu5Ac-Gg4, in Neuroblastoma Cells. Mol. Neurobiol. 2019, 56, 6673–6702. [Google Scholar] [CrossRef] [PubMed]
- Saulino, M.F.; Schengrund, C.L. Effects of specific gangliosides on the in vitro proliferation of MPTP-susceptible cells. J. Neurochem. 1993, 61, 1277–1283. [Google Scholar] [CrossRef] [PubMed]
- De Girolamo, L.A.; Hargreaves, A.J.; Billett, E.E. Protection from MPTP-induced neurotoxicity in differentiating mouse N2a neuroblastoma cells. J. Neurochem. 2001, 76, 650–660. [Google Scholar] [CrossRef]
- Fazzari, M.; Audano, M.; Lunghi, G.; Di Biase, E.; Loberto, N.; Mauri, L.; Mitro, N.; Sonnino, S.; Chiricozzi, E. The oligosaccharide portion of ganglioside GM1 regulates mitochondrial function in neuroblastoma cells. Glycoconj. J. 2020, 37, 293–306. [Google Scholar] [CrossRef]
- Di Biase, E.; Lunghi, G.; Fazzari, M.; Maggioni, M.; Pome, D.Y.; Valsecchi, M.; Samarani, M.; Fato, P.; Ciampa, M.G.; Prioni, S.; et al. Gangliosides in the differentiation process of primary neurons: The specific role of GM1-oligosaccharide. Glycoconj. J. 2020, 37, 329–343. [Google Scholar] [CrossRef]
- Hu, S.; Mak, S.; Zuo, X.; Li, H.; Wang, Y.; Han, Y. Neuroprotection Against MPP(+)-Induced Cytotoxicity Through the Activation of PI3-K/Akt/GSK3beta/MEF2D Signaling Pathway by Rhynchophylline, the Major Tetracyclic Oxindole Alkaloid Isolated from Uncaria rhynchophylla. Front. Pharmacol. 2018, 9, 768. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Lunghi, G.; Fazzari, M.; Di Biase, E.; Mauri, L.; Sonnino, S.; Chiricozzi, E. Modulation of calcium signaling depends on the oligosaccharide of GM1 in Neuro2a mouse neuroblastoma cells. Glycoconj. J. 2020, 37, 713–720. [Google Scholar] [CrossRef]
- Spath, M.; Aichert, I.; Timmann, D.; Ceballos-Baumann, A.O.; Wagner-Sonntag, E.; Ziegler, W. The role of the basal ganglia and cerebellum in adaptation to others’ speech rate and rhythm: A study of patients with Parkinson’s disease and cerebellar degeneration. Cortex 2022, 157, 81–98. [Google Scholar] [CrossRef] [PubMed]
- van der Zee, S.; Kanel, P.; Muller, M.; van Laar, T.; Bohnen, N.I. Identification of cholinergic centro-cingulate topography as main contributor to cognitive functioning in Parkinson’s disease: Results from a data-driven approach. Front. Aging Neurosci. 2022, 14, 1006567. [Google Scholar] [CrossRef] [PubMed]
- Rusholt, E.H.L.; Salvesen, L.; Brudek, T.; Tesfay, B.; Pakkenberg, B.; Olesen, M.V. Pathological changes in the cerebellum of patients with multiple system atrophy and Parkinson’s disease-a stereological study. Brain Pathol. 2020, 30, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Selvakumar, T.; Kilpatrick, D.L. Culturing mouse cerebellar granule neurons. Methods Mol. Biol. 2013, 1018, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Consalez, G.G.; Goldowitz, D.; Casoni, F.; Hawkes, R. Origins, Development, and Compartmentation of the Granule Cells of the Cerebellum. Front. Neural Circuits 2020, 14, 611841. [Google Scholar] [CrossRef]
- Guo, J.N.; Tian, L.Y.; Liu, W.Y.; Mu, J.; Zhou, D. Activation of the Akt/mTOR signaling pathway: A potential response to long-term neuronal loss in the hippocampus after sepsis. Neural Regen. Res. 2017, 12, 1832–1842. [Google Scholar] [CrossRef]
- Babichev, Y.; Kabaroff, L.; Datti, A.; Uehling, D.; Isaac, M.; Al-Awar, R.; Prakesch, M.; Sun, R.X.; Boutros, P.C.; Venier, R.; et al. PI3K/AKT/mTOR inhibition in combination with doxorubicin is an effective therapy for leiomyosarcoma. J. Transl. Med. 2016, 14, 67. [Google Scholar] [CrossRef]
- Gu, C.; Zhang, Y.; Hu, Q.; Wu, J.; Ren, H.; Liu, C.F.; Wang, G. P7C3 inhibits GSK3beta activation to protect dopaminergic neurons against neurotoxin-induced cell death in vitro and in vivo. Cell Death Dis. 2017, 8, e2858. [Google Scholar] [CrossRef]
- Golpich, M.; Amini, E.; Hemmati, F.; Ibrahim, N.M.; Rahmani, B.; Mohamed, Z.; Raymond, A.A.; Dargahi, L.; Ghasemi, R.; Ahmadiani, A. Glycogen synthase kinase-3 beta (GSK-3beta) signaling: Implications for Parkinson’s disease. Pharmacol. Res. 2015, 97, 16–26. [Google Scholar] [CrossRef]
- Dalia, A.; Neff, N.H.; Hadjiconstantinou, M. GM1 ganglioside improves dopaminergic markers of rat mesencephalic cultures treated with MPP+. J. Neurosci. 1993, 13, 3104–3111. [Google Scholar] [CrossRef]
- Stull, N.D.; Schneider, J.S.; Iacovitti, L. GM1 ganglioside partially rescues cultured dopaminergic neurons from MPP(+)-induced damage: Dependence on initial damage and time of treatment. Brain Res. 1994, 640, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Fikry, H.; Saleh, L.A.; Abdel Gawad, S. Neuroprotective effects of curcumin on the cerebellum in a rotenone-induced Parkinson’s Disease Model. CNS Neurosci. Ther. 2022, 28, 732–748. [Google Scholar] [CrossRef] [PubMed]
- Gatt, A.P.; Duncan, O.F.; Attems, J.; Francis, P.T.; Ballard, C.G.; Bateman, J.M. Dementia in Parkinson’s disease is associated with enhanced mitochondrial complex I deficiency. Mov. Disord. 2016, 31, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.J.; Li, K.S.; Shen, Y.Q. Activated immune cells in Parkinson’s disease. J. Neuroimmune Pharmacol. 2011, 6, 323–329. [Google Scholar] [CrossRef]
- Mullin, S.; Schapira, A.H. Pathogenic mechanisms of neurodegeneration in Parkinson disease. Neurol Clin. 2015, 33, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Di Biase, E.; Lunghi, G.; Maggioni, M.; Fazzari, M.; Pome, D.Y.; Loberto, N.; Ciampa, M.G.; Fato, P.; Mauri, L.; Sevin, E.; et al. GM1 Oligosaccharide Crosses the Human Blood-Brain Barrier In Vitro by a Paracellular Route. Int. J. Mol. Sci. 2020, 21, 2858. [Google Scholar] [CrossRef]
- Svennerholm, L.; Brane, G.; Karlsson, I.; Lekman, A.; Ramstrom, I.; Wikkelso, C. Alzheimer disease—Effect of continuous intracerebroventricular treatment with GM1 ganglioside and a systematic activation programme. Dement. Geriatr. Cogn. Disord. 2002, 14, 128–136. [Google Scholar] [CrossRef]
- Chester, M.A. IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN). Nomenclature of glycolipids--recommendations 1997. Eur. J. Biochem. 1998, 257, 293–298. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fazzari, M.; Lunghi, G.; Henriques, A.; Callizot, N.; Ciampa, M.G.; Mauri, L.; Prioni, S.; Carsana, E.V.; Loberto, N.; Aureli, M.; et al. GM1 Oligosaccharide Efficacy in Parkinson’s Disease: Protection against MPTP. Biomedicines 2023, 11, 1305. https://doi.org/10.3390/biomedicines11051305
Fazzari M, Lunghi G, Henriques A, Callizot N, Ciampa MG, Mauri L, Prioni S, Carsana EV, Loberto N, Aureli M, et al. GM1 Oligosaccharide Efficacy in Parkinson’s Disease: Protection against MPTP. Biomedicines. 2023; 11(5):1305. https://doi.org/10.3390/biomedicines11051305
Chicago/Turabian StyleFazzari, Maria, Giulia Lunghi, Alexandre Henriques, Noëlle Callizot, Maria Grazia Ciampa, Laura Mauri, Simona Prioni, Emma Veronica Carsana, Nicoletta Loberto, Massimo Aureli, and et al. 2023. "GM1 Oligosaccharide Efficacy in Parkinson’s Disease: Protection against MPTP" Biomedicines 11, no. 5: 1305. https://doi.org/10.3390/biomedicines11051305
APA StyleFazzari, M., Lunghi, G., Henriques, A., Callizot, N., Ciampa, M. G., Mauri, L., Prioni, S., Carsana, E. V., Loberto, N., Aureli, M., Mari, L., Sonnino, S., Chiricozzi, E., & Di Biase, E. (2023). GM1 Oligosaccharide Efficacy in Parkinson’s Disease: Protection against MPTP. Biomedicines, 11(5), 1305. https://doi.org/10.3390/biomedicines11051305