
Aging in Ocular Blood Vessels: Molecular Insights and the Role of Oxidative Stress



Abstract
1. Introduction
2. The Concepts of Aging and Vascular Aging
2.1. General Insights into Aging
2.2. Insights into Vascular Aging
3. Vascular Aging in Ocular Blood Vessels
3.1. The Choroid
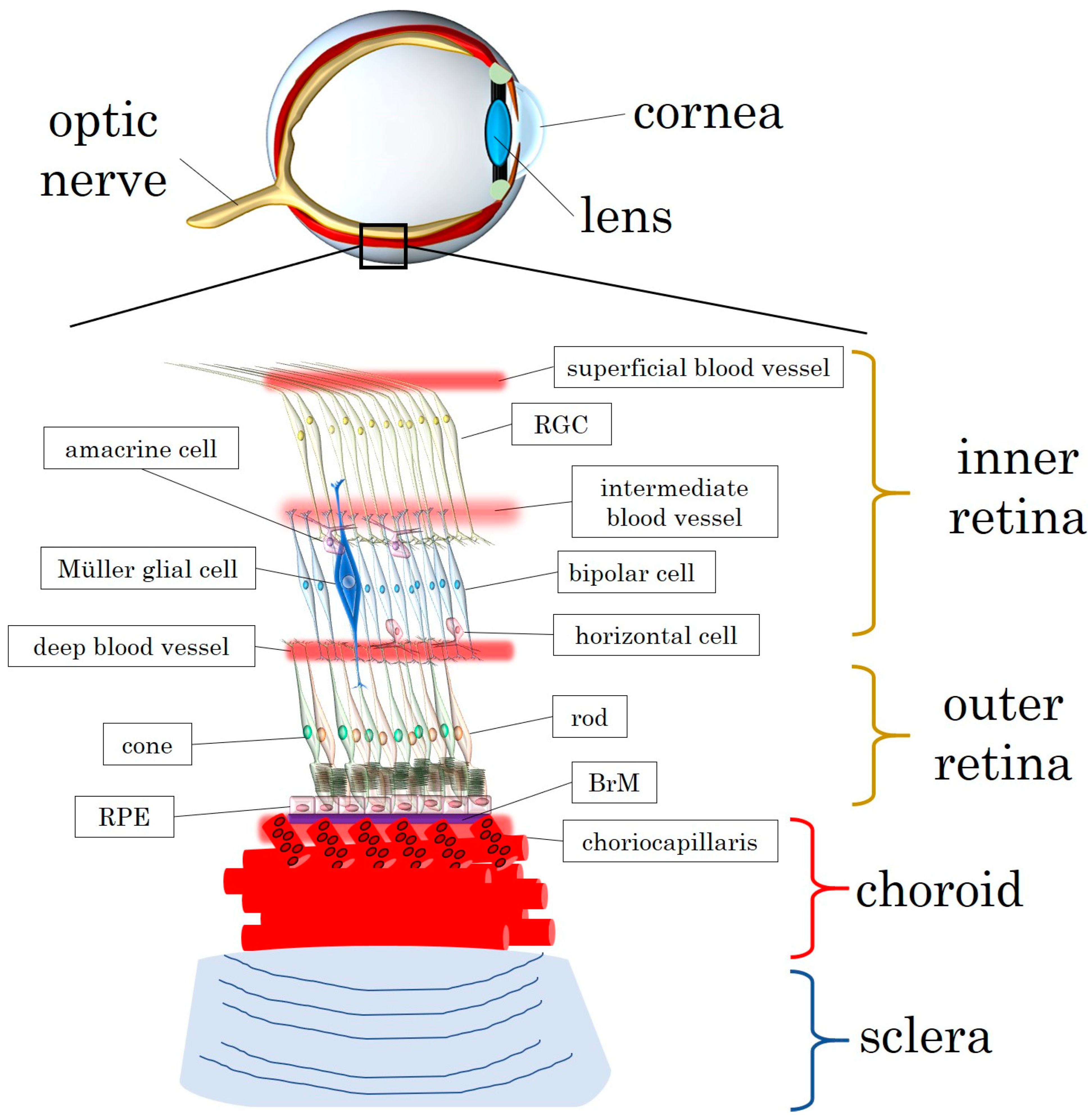
3.1.1. Choroidal Vascular Circulation: Physiological Significance
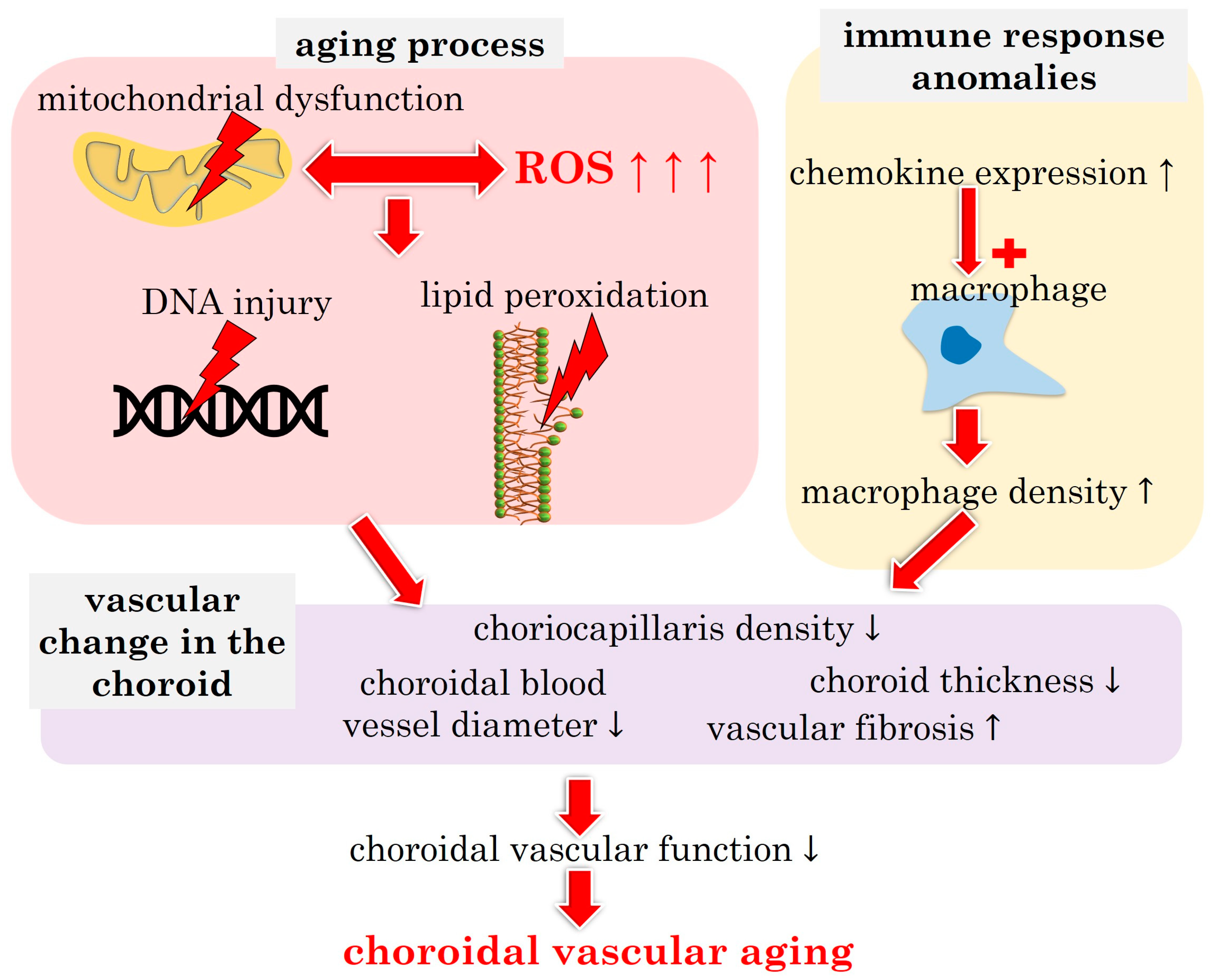
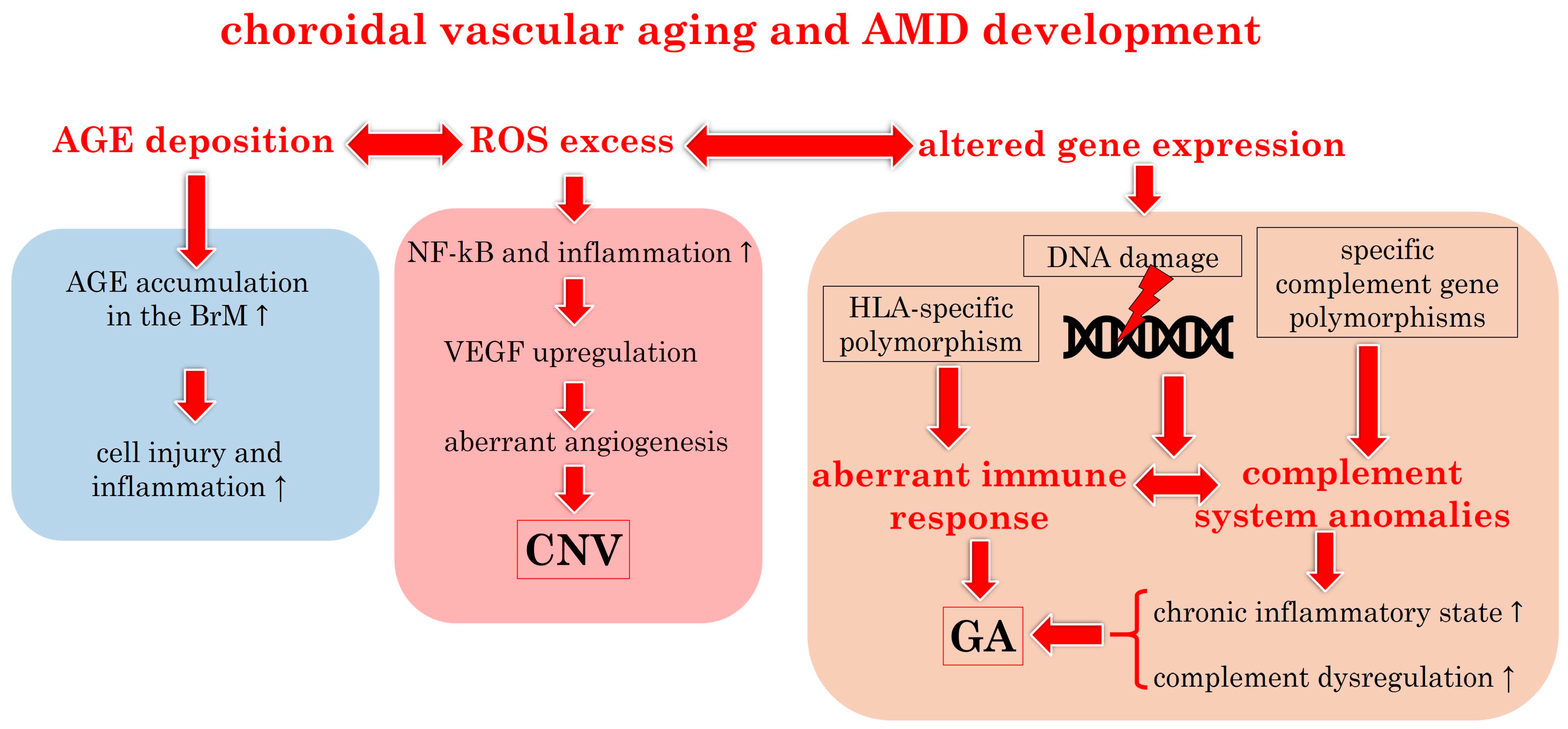
3.1.2. Choroidal Vascular Aging: Pathophysiological Insights
3.1.3. Molecular Insights into Choroidal Vascular Aging
3.2. Retina
3.2.1. Retinal Vascular Circulation: Physiological Significance
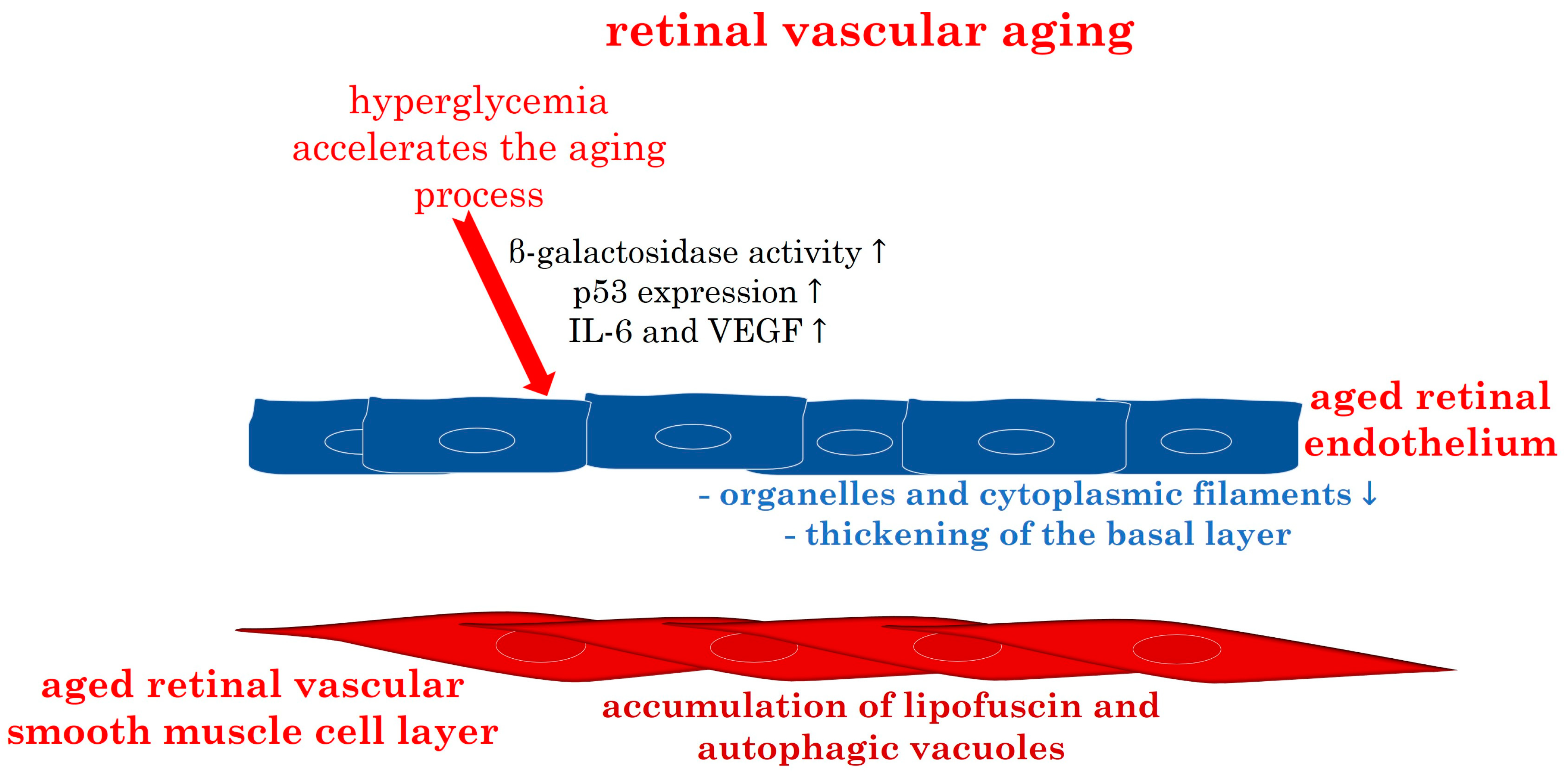
3.2.2. Retinal Vascular Aging: Pathophysiology
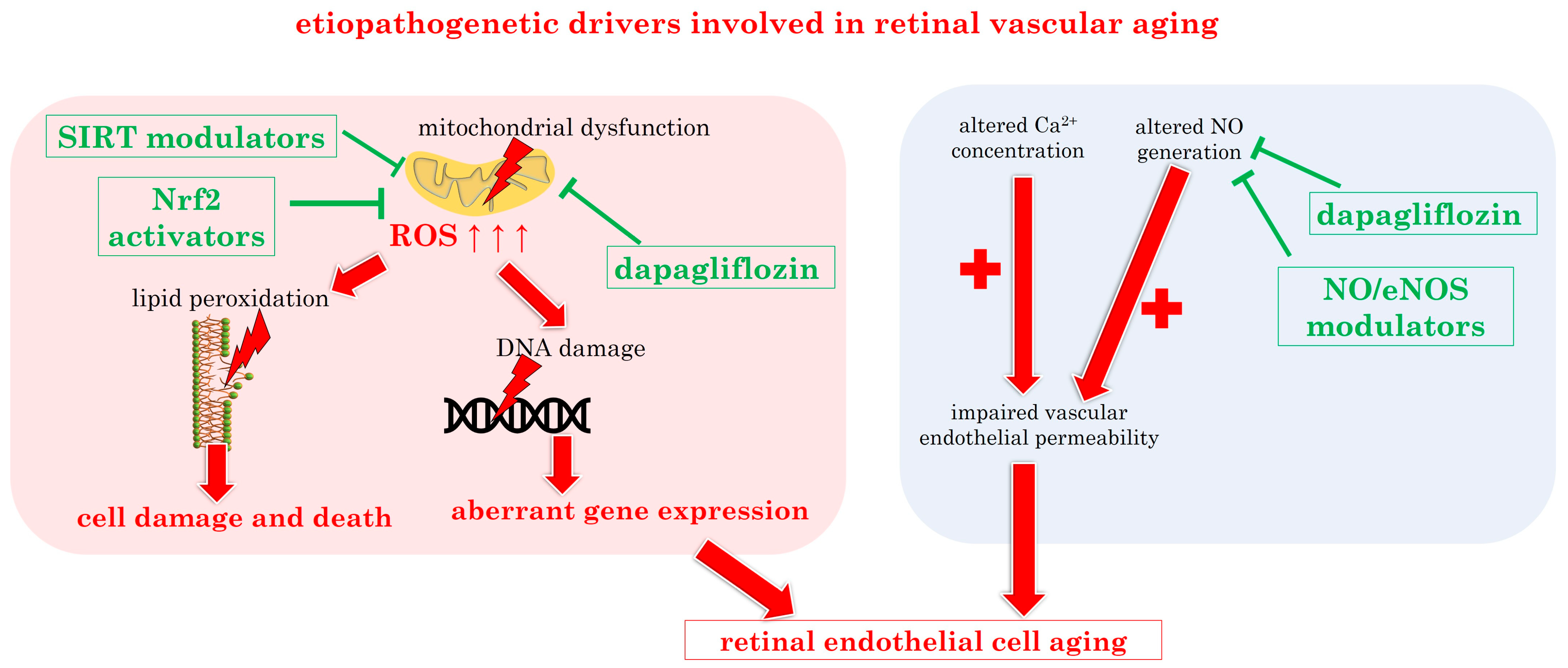
3.2.3. Molecular Insights into Retinal Vascular Aging: The Pivotal Role of Oxidative Stress
4. Emerging Therapeutic Strategies and Novel Targeting Pathways
4.1. Modulation of the eNOS/NO Pathway
4.2. Nrf2 Activators and Mitochondria-Targeting Molecules
4.3. SIRT Activators, Telomerase Enhancers, and Senolytics
4.4. Antidiabetics as Anti-Aging Drugs
4.5. Complement Inhibitors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bektas, A.; Schurman, S.H.; Sen, R.; Ferrucci, L. Aging, Inflammation and the Environment. Exp. Gerontol. 2018, 105, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zheng, Y.; Goronzy, J.J.; Weyand, C.M. T Cell Aging as a Risk Factor for Autoimmunity. J. Autoimmun. 2023, 137, 102947. [Google Scholar] [CrossRef] [PubMed]
- Gloor, A.D.; Berry, G.J.; Goronzy, J.J.; Weyand, C.M. Age as a Risk Factor in Vasculitis. Semin. Immunopathol. 2022, 44, 281–301. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.; Třískala, Z.; Honců, P.; Krejčí, M.; Kajzar, J.; Bičíková, M.; Ondřejíková, L.; Jandová, D.; Sterzl, I. Aging, Hormones and Receptors. Physiol. Res. 2020, 69, S255–S272. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Kobayashi, T.; Shiba, T.; Hayashi, N. Effects of Aging and Exercise Habits on Blood Flow Profile of the Ocular Circulation. PLoS ONE 2022, 17, e0266684. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; Tang, X.; Hu, A.; Fan, M.; Wu, P.; Lu, X.; Lin, J.; Yang, F.; Zhao, X.; Huang, J.; et al. Therapeutic Benefit of Melatonin in Choroidal Neovascularization during Aging through the Regulation of Senescent Macrophage/Microglia Polarization. Investig. Ophthalmol. Vis. Sci. 2023, 64, 19. [Google Scholar] [CrossRef] [PubMed]
- Hayflick, L. The Limited In Vitro Lifetime of Human Diploid Cell Strains. Exp. Cell Res. 1965, 37, 614–636. [Google Scholar] [CrossRef]
- Hayflick, L.; Moorhead, P.S. The Serial Cultivation of Human Diploid Cell Strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef] [PubMed]
- Regulski, M.J. Cellular Senescence: What, Why, and How. Wounds 2017, 29, 168–174. [Google Scholar]
- Kudlova, N.; De Sanctis, J.B.; Hajduch, M. Cellular Senescence: Molecular Targets, Biomarkers, and Senolytic Drugs. Int. J. Mol. Sci. 2022, 23, 4168. [Google Scholar] [CrossRef]
- Kritsilis, M.; V Rizou, S.; Koutsoudaki, P.N.; Evangelou, K.; Gorgoulis, V.G.; Papadopoulos, D. Ageing, Cellular Senescence and Neurodegenerative Disease. Int. J. Mol. Sci. 2018, 19, 2937. [Google Scholar] [CrossRef] [PubMed]
- Childs, B.G.; Baker, D.J.; Wijshake, T.; Conover, C.A.; Campisi, J.; van Deursen, J.M. Senescent Intimal Foam Cells Are Deleterious at All Stages of Atherosclerosis. Science 2016, 354, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.J.; Childs, B.G.; Durik, M.; Wijers, M.E.; Sieben, C.J.; Zhong, J.; Saltness, R.A.; Jeganathan, K.B.; Verzosa, G.C.; Pezeshki, A.; et al. Naturally Occurring P16(Ink4a)-Positive Cells Shorten Healthy Lifespan. Nature 2016, 530, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.J.; Wijshake, T.; Tchkonia, T.; LeBrasseur, N.K.; Childs, B.G.; van de Sluis, B.; Kirkland, J.L.; van Deursen, J.M. Clearance of p16Ink4a-Positive Senescent Cells Delays Ageing-Associated Disorders. Nature 2011, 479, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Coppé, J.-P.; Patil, C.K.; Rodier, F.; Sun, Y.; Muñoz, D.P.; Goldstein, J.; Nelson, P.S.; Desprez, P.-Y.; Campisi, J. Senescence-Associated Secretory Phenotypes Reveal Cell-Nonautonomous Functions of Oncogenic RAS and the P53 Tumor Suppressor. PLoS Biol. 2008, 6, 2853–2868. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of Aging: An Expanding Universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-Aging. An Evolutionary Perspective on Immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.J.; Wang, E.K.; Benayoun, B.A. Functional genomics of inflamm-aging and immunosenescence. Brief Funct Genomics. 2022, 21, 43–55. [Google Scholar] [CrossRef]
- Xia, S.; Zhang, X.; Zheng, S.; Khanabdali, R.; Kalionis, B.; Wu, J.; Wan, W.; Tai, X. An Update on Inflamm-Aging: Mechanisms, Prevention, and Treatment. J. Immunol. Res. 2016, 2016, 8426874. [Google Scholar] [CrossRef]
- Fraser, H.C.; Kuan, V.; Johnen, R.; Zwierzyna, M.; Hingorani, A.D.; Beyer, A.; Partridge, L. Biological Mechanisms of Aging Predict Age-Related Disease Co-Occurrence in Patients. Aging Cell 2022, 21, e13524. [Google Scholar] [CrossRef] [PubMed]
- M Nilsson, P. Early Vascular Ageing—A Concept in Development. Eur. Endocrinol. 2015, 11, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.J.-E.; Bodary, P.F. How Old Are Your Arteries? Exercise-Mediated Protection from Age-Associated Vascular Stiffness. J. Am. Heart Assoc. 2014, 3, e000941. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.C.; Cunha, P.M.G.M.; Vitorino, P.V.O.; Souza, A.L.L.; Deus, G.D.; Feitosa, A.; Barbosa, E.C.D.; Gomes, M.M.; Jardim, P.C.B.V.; Barroso, W.K.S. Vascular Aging and Arterial Stiffness. Arq. Bras. Cardiol. 2022, 119, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Rizzoni, D.; Rizzoni, M.; Nardin, M.; Chiarini, G.; Agabiti-Rosei, C.; Aggiusti, C.; Paini, A.; Salvetti, M.; Muiesan, M.L. Vascular Aging and Disease of the Small Vessels. High Blood Press. Cardiovasc. Prev. 2019, 26, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Ungvari, Z.; Tarantini, S.; Kiss, T.; Wren, J.D.; Giles, C.B.; Griffin, C.T.; Murfee, W.L.; Pacher, P.; Csiszar, A. Endothelial dysfunction and angiogenesis impairment in the ageing vasculature. Nat. Rev. Cardiol. 2018, 15, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Ungvari, Z.; Tarantini, S.; Donato, A.J.; Galvan, V.; Csiszar, A. Mechanisms of Vascular Aging. Circ. Res. 2018, 123, 849–867. [Google Scholar] [CrossRef] [PubMed]
- Maloberti, A.; Vallerio, P.; Triglione, N.; Occhi, L.; Panzeri, F.; Bassi, I.; Pansera, F.; Piccinelli, E.; Peretti, A.; Garatti, L.; et al. Vascular Aging and Disease of the Large Vessels: Role of Inflammation. High Blood Press. Cardiovasc. Prev. 2019, 26, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Cortes-Canteli, M.; Iadecola, C. Alzheimer’s Disease and Vascular Aging: JACC Focus Seminar. J. Am. Coll. Cardiol. 2020, 75, 942–951. [Google Scholar] [CrossRef]
- Culley, M.K.; Chan, S.Y. Endothelial Senescence: A New Age in Pulmonary Hypertension. Circ. Res. 2022, 130, 928–941. [Google Scholar] [CrossRef]
- De Silva, T.M.; Faraci, F.M. Contributions of Aging to Cerebral Small Vessel Disease. Annu. Rev. Physiol. 2020, 82, 275–295. [Google Scholar] [CrossRef]
- Sayin, N.; Kara, N.; Pekel, G.; Altinkaynak, H. Choroidal Thickness Changes after Dynamic Exercise as Measured by Spectral-Domain Optical Coherence Tomography. Indian. J. Ophthalmol. 2015, 63, 445–450. [Google Scholar] [CrossRef]
- Nickla, D.L.; Wallman, J. The Multifunctional Choroid. Prog. Retin. Eye Res. 2010, 29, 144–168. [Google Scholar] [CrossRef]
- Fields, M.A.; Del Priore, L.V.; Adelman, R.A.; Rizzolo, L.J. Interactions of the Choroid, Bruch’s Membrane, Retinal Pigment Epithelium, and Neurosensory Retina Collaborate to Form the Outer Blood-Retinal-Barrier. Prog. Retin. Eye Res. 2020, 76, 100803. [Google Scholar] [CrossRef] [PubMed]
- Kur, J.; Newman, E.A.; Chan-Ling, T. Cellular and Physiological Mechanisms Underlying Blood Flow Regulation in the Retina and Choroid in Health and Disease. Prog. Retin. Eye Res. 2012, 31, 377–406. [Google Scholar] [CrossRef] [PubMed]
- Hayreh, S.S.; Hayreh, S.B. Uveal Vascular Bed in Health and Disease: Uveal Vascular Bed Anatomy. Paper 1 of 2. Eye 2023, 37, 2590–2616. [Google Scholar] [CrossRef] [PubMed]
- Spaide, R.F. CHOROIDAL BLOOD FLOW: Review and Potential Explanation for the Choroidal Venous Anatomy Including the Vortex Vein System. Retina 2020, 40, 1851–1864. [Google Scholar] [CrossRef]
- Weiter, J.J.; Ernest, J.T. Anatomy of the Choroidal Vasculature. Am. J. Ophthalmol. 1974, 78, 583–590. [Google Scholar] [CrossRef]
- Reiner, A.; Fitzgerald, M.E.C.; Del Mar, N.; Li, C. Neural Control of Choroidal Blood Flow. Prog. Retin. Eye Res. 2018, 64, 96–130. [Google Scholar] [CrossRef]
- Ostrin, L.A.; Harb, E.; Nickla, D.L.; Read, S.A.; Alonso-Caneiro, D.; Schroedl, F.; Kaser-Eichberger, A.; Zhou, X.; Wildsoet, C.F. IMI-The Dynamic Choroid: New Insights, Challenges, and Potential Significance for Human Myopia. Invest. Ophthalmol. Vis. Sci. 2023, 64, 4. [Google Scholar] [CrossRef]
- Wright, W.S.; Eshaq, R.S.; Lee, M.; Kaur, G.; Harris, N.R. Retinal Physiology and Circulation: Effect of Diabetes. Compr. Physiol. 2020, 10, 933–974. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, L.; Xu, Y.; Pang, Z.; Mu, G. The Influence of the Choroid on the Onset and Development of Myopia: From Perspectives of Choroidal Thickness and Blood Flow. Acta Ophthalmol. 2021, 99, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Behar-Cohen, F.; Gelizé, E.; Jonet, L.; Lassiaz, P. [Anatomy of the retina]. Med. Sci. 2020, 36, 594–599. [Google Scholar] [CrossRef]
- Barteselli, G.; Chhablani, J.; El-Emam, S.; Wang, H.; Chuang, J.; Kozak, I.; Cheng, L.; Bartsch, D.-U.; Freeman, W.R. Choroidal Volume Variations with Age, Axial Length, and Sex in Healthy Subjects: A Three-Dimensional Analysis. Ophthalmology 2012, 119, 2572–2578. [Google Scholar] [CrossRef] [PubMed]
- Grossniklaus, H.E.; Nickerson, J.M.; Edelhauser, H.F.; Bergman, L.A.M.K.; Berglin, L. Anatomic Alterations in Aging and Age-Related Diseases of the Eye. Invest. Ophthalmol. Vis. Sci. 2013, 54, ORSF23–ORSF27. [Google Scholar] [CrossRef] [PubMed]
- Ungvari, Z.; Tarantini, S.; Sorond, F.; Merkely, B.; Csiszar, A. Mechanisms of Vascular Aging, A Geroscience Perspective: JACC Focus Seminar. J. Am. Coll. Cardiol. 2020, 75, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Lipecz, A.; Miller, L.; Kovacs, I.; Czakó, C.; Csipo, T.; Baffi, J.; Csiszar, A.; Tarantini, S.; Ungvari, Z.; Yabluchanskiy, A.; et al. Microvascular Contributions to Age-Related Macular Degeneration (AMD): From Mechanisms of Choriocapillaris Aging to Novel Interventions. Geroscience 2019, 41, 813–845. [Google Scholar] [CrossRef] [PubMed]
- Seddon, J.M.; McLeod, D.S.; Bhutto, I.A.; Villalonga, M.B.; Silver, R.E.; Wenick, A.S.; Edwards, M.M.; Lutty, G.A. Histopathological Insights into Choroidal Vascular Loss in Clinically Documented Cases of Age-Related Macular Degeneration. JAMA Ophthalmol. 2016, 134, 1272–1280. [Google Scholar] [CrossRef] [PubMed]
- Pache, M.; Meyer, P.; Prünte, C.; Orgül, S.; Nuttli, I.; Flammer, J. Sildenafil Induces Retinal Vasodilatation in Healthy Subjects. Br. J. Ophthalmol. 2002, 86, 156–158. [Google Scholar] [CrossRef][Green Version]
- Yiu, G.; Vuong, V.S.; Tran, S.; Migacz, J.; Cunefare, D.; Farsiu, S.; Khandelwal, N.; Agrawal, R.; Cheung, C.M.G. Vascular Response to Sildenafil Citrate in Aging and Age-Related Macular Degeneration. Sci. Rep. 2019, 9, 5049. [Google Scholar] [CrossRef]
- Zhao, B.; Zhu, L.; Ye, M.; Lou, X.; Mou, Q.; Hu, Y.; Zhang, H.; Zhao, Y. Oxidative stress and epigenetics in ocular vascular aging: An updated review. Mol. Med. 2023, 29, 28. [Google Scholar] [CrossRef] [PubMed]
- Hidayat, A.A.; Fine, B.S. Diabetic choroidopathy. Light and electron microscopic observations of seven cases. Ophthalmology. 1985, 92, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Shiragami, C.; Shiraga, F.; Matsuo, T.; Tsuchida, Y.; Ohtsuki, H. Risk factors for diabetic choroidopathy in patients with diabetic retinopathy. Graefes Arch Clin. Exp. Ophthalmol. 2002, 240, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Hamadneh, T.; Aftab, S.; Sherali, N.; Vetrivel Suresh, R.; Tsouklidis, N.; An, M. Choroidal Changes in Diabetic Patients with Different Stages of Diabetic Retinopathy. Cureus 2020, 12, e10871. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.; Liu, J.; Deng, W.; Luo, C.; Qi, J.; Chen, M.; Xu, H. Old Age Promotes Retinal Fibrosis in Choroidal Neovascularization through Circulating Fibrocytes and Profibrotic Macrophages. J. Neuroinflamm. 2023, 20, 45. [Google Scholar] [CrossRef] [PubMed]
- Nivison-Smith, L.; Khandelwal, N.; Tong, J.; Mahajan, S.; Kalloniatis, M.; Agrawal, R. Normal Aging Changes in the Choroidal Angioarchitecture of the Macula. Sci. Rep. 2020, 10, 10810. [Google Scholar] [CrossRef]
- Kumar, A.; Zhao, L.; Fariss, R.N.; McMenamin, P.G.; Wong, W.T. Vascular Associations and Dynamic Process Motility in Perivascular Myeloid Cells of the Mouse Choroid: Implications for Function and Senescent Change. Invest. Ophthalmol. Vis. Sci. 2014, 55, 1787–1796. [Google Scholar] [CrossRef]
- Green, W.R.; Key, S.N. Senile Macular Degeneration: A Histopathologic Study. Trans. Am. Ophthalmol. Soc. 1977, 75, 180–254. [Google Scholar] [CrossRef]
- McLeod, D.S.; Lutty, G.A. High-Resolution Histologic Analysis of the Human Choroidal Vasculature. Invest. Ophthalmol. Vis. Sci. 1994, 35, 3799–3811. [Google Scholar] [PubMed]
- Fragiotta, S.; Scuderi, L.; Iodice, C.M.; Rullo, D.; Di Pippo, M.; Maugliani, E.; Abdolrahimzadeh, S. Choroidal Vasculature Changes in Age-Related Macular Degeneration: From a Molecular to a Clinical Perspective. Int. J. Mol. Sci. 2022, 23, 12010. [Google Scholar] [CrossRef]
- Grunwald, J.E.; Hariprasad, S.M.; DuPont, J. Effect of Aging on Foveolar Choroidal Circulation. Arch. Ophthalmol. 1998, 116, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.; Gupta, P.; Tan, K.-A.; Cheung, C.M.G.; Wong, T.-Y.; Cheng, C.-Y. Choroidal Vascularity Index as a Measure of Vascular Status of the Choroid: Measurements in Healthy Eyes from a Population-Based Study. Sci. Rep. 2016, 6, 21090. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Liu, Y.; Huang, L.; Tan, H. Advances in epigenetic regulation of vascular aging. Rev. Cardiovasc. Med. 2019, 20, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The Impact of Oxidative Stress and Inflammation on RPE Degeneration in Non-Neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Buonfiglio, F.; Korb, C.A.; Stoffelns, B.; Pfeiffer, N.; Gericke, A. Recent Advances in Our Understanding of Age-Related Macular Degeneration: Mitochondrial Dysfunction, Redox Signaling, and the Complement System. Aging Dis. 2024. [Google Scholar] [CrossRef]
- Goverdhan, S.V.; Howell, M.W.; Mullins, R.F.; Osmond, C.; Hodgkins, P.R.; Self, J.; Avery, K.; Lotery, A.J. Association of HLA Class I and Class II Polymorphisms with Age-Related Macular Degeneration. Invest. Ophthalmol. Vis. Sci. 2005, 46, 1726–1734. [Google Scholar] [CrossRef] [PubMed]
- Bhutto, I.; Lutty, G. Understanding age-related macular degeneration (AMD): Relationships between the photoreceptor/retinal pigment epithelium/Bruch’s membrane/choriocapillaris complex. Mol. Asp. Med. 2012, 33, 295–317. [Google Scholar] [CrossRef] [PubMed]
- Chirco, K.R.; Sohn, E.H.; Stone, E.M.; Tucker, B.A.; Mullins, R.F. Structural and Molecular Changes in the Aging Choroid: Implications for Age-Related Macular Degeneration. Eye 2017, 31, 10–25. [Google Scholar] [CrossRef]
- Steinle, J.J.; Sharma, S.; Chin, V.C. Normal Aging Involves Altered Expression of Growth Factors in the Rat Choroid. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 135–140. [Google Scholar] [CrossRef][Green Version]
- Chen, M.; Glenn, J.V.; Dasari, S.; McVicar, C.; Ward, M.; Colhoun, L.; Quinn, M.; Bierhaus, A.; Xu, H.; Stitt, A.W. RAGE Regulates Immune Cell Infiltration and Angiogenesis in Choroidal Neovascularization. PLoS ONE 2014, 9, e89548. [Google Scholar] [CrossRef]
- Bhutto, I.A.; Baba, T.; Merges, C.; Juriasinghani, V.; McLeod, D.S.; Lutty, G.A. C-Reactive Protein and Complement Factor H in Aged Human Eyes and Eyes with Age-Related Macular Degeneration. Br. J. Ophthalmol. 2011, 95, 1323–1330. [Google Scholar] [CrossRef]
- Robbie, S.J.; Georgiadis, A.; Barker, S.E.; Duran, Y.; Smith, A.J.; Ali, R.R.; Luhmann, U.F.O.; Bainbridge, J.W. Enhanced Ccl2-Ccr2 Signaling Drives More Severe Choroidal Neovascularization with Aging. Neurobiol. Aging 2016, 40, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.H.; Chowers, I.; Lotery, A.J. Beyond the Complement Cascade: Insights into Systemic Immunosenescence and Inflammaging in Age-Related Macular Degeneration and Current Barriers to Treatment. Cells 2023, 12, 1708. [Google Scholar] [CrossRef]
- Addabbo, F.; Ratliff, B.; Park, H.-C.; Kuo, M.-C.; Ungvari, Z.; Csiszar, A.; Krasnikov, B.; Sodhi, K.; Zhang, F.; Nasjletti, A.; et al. The Krebs Cycle and Mitochondrial Mass Are Early Victims of Endothelial Dysfunction: Proteomic Approach. Am. J. Pathol. 2009, 174, 34–43. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Springo, Z.; Tarantini, S.; Toth, P.; Tucsek, Z.; Koller, A.; Sonntag, W.E.; Csiszar, A.; Ungvari, Z. Aging Exacerbates Pressure-Induced Mitochondrial Oxidative Stress in Mouse Cerebral Arteries. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 70, 1355–1359. [Google Scholar] [CrossRef]
- Tarantini, S.; Valcarcel-Ares, N.M.; Yabluchanskiy, A.; Fulop, G.A.; Hertelendy, P.; Gautam, T.; Farkas, E.; Perz, A.; Rabinovitch, P.S.; Sonntag, W.E.; et al. Treatment with the Mitochondrial-Targeted Antioxidant Peptide SS-31 Rescues Neurovascular Coupling Responses and Cerebrovascular Endothelial Function and Improves Cognition in Aged Mice. Aging Cell 2018, 17, e12731. [Google Scholar] [CrossRef] [PubMed]
- Country, M.W. Retinal metabolism: A comparative look at energetics in the retina. Brain Res. 2017, 1672, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Pournaras, C.J.; Rungger-Brändle, E.; Riva, C.E.; Hardarson, S.H.; Stefansson, E. Regulation of Retinal Blood Flow in Health and Disease. Prog. Retin. Eye Res. 2008, 27, 284–330. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Balne, P.K.; Meissner, K.E.; Barathi, V.A.; Schmetterer, L.; Agrawal, R. Assessment of Flow Dynamics in Retinal and Choroidal Microcirculation. Surv. Ophthalmol. 2018, 63, 646–664. [Google Scholar] [CrossRef]
- Yu, D.-Y.; Cringle, S.J.; Yu, P.K.; Balaratnasingam, C.; Mehnert, A.; Sarunic, M.V.; An, D.; Su, E.-N. Retinal Capillary Perfusion: Spatial and Temporal Heterogeneity. Prog. Retin. Eye Res. 2019, 70, 23–54. [Google Scholar] [CrossRef]
- Funk, R.H. Blood Supply of the Retina. Ophthalmic Res. 1997, 29, 320–325. [Google Scholar] [CrossRef]
- Wei, Y.; Jiang, H.; Shi, Y.; Qu, D.; Gregori, G.; Zheng, F.; Rundek, T.; Wang, J. Age-Related Alterations in the Retinal Microvasculature, Microcirculation, and Microstructure. Invest. Ophthalmol. Vis. Sci. 2017, 58, 3804–3817. [Google Scholar] [CrossRef] [PubMed]
- Teberik, K.; Kaya, M. Retinal and Choroidal Thickness in Patients with High Myopia without Maculopathy. Pak. J. Med. Sci. 2017, 33, 1438–1443. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, F.; Campbell, M. The Blood-Retina Barrier in Health and Disease. FEBS J. 2023, 290, 878–891. [Google Scholar] [CrossRef] [PubMed]
- Leley, S.P.; Ciulla, T.A.; Bhatwadekar, A.D. Diabetic Retinopathy in the Aging Population: A Perspective of Pathogenesis and Treatment. Clin. Interv. Aging 2021, 16, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Binet, F.; Cagnone, G.; Crespo-Garcia, S.; Hata, M.; Neault, M.; Dejda, A.; Wilson, A.M.; Buscarlet, M.; Mawambo, G.T.; Howard, J.P.; et al. Neutrophil Extracellular Traps Target Senescent Vasculature for Tissue Remodeling in Retinopathy. Science 2020, 369, eaay5356. [Google Scholar] [CrossRef] [PubMed]
- Beckman, J.A.; Creager, M.A. Vascular Complications of Diabetes. Circ. Res. 2016, 118, 1771–1785. [Google Scholar] [CrossRef] [PubMed]
- Gericke, A.; Suminska-Jasińska, K.; Bręborowicz, A. Sulodexide Reduces Glucose Induced Senescence in Human Retinal Endothelial Cells. Sci. Rep. 2021, 11, 11532. [Google Scholar] [CrossRef] [PubMed]
- Yue, T.; Shi, Y.; Luo, S.; Weng, J.; Wu, Y.; Zheng, X. The Role of Inflammation in Immune System of Diabetic Retinopathy: Molecular Mechanisms, Pathogenetic Role and Therapeutic Implications. Front. Immunol. 2022, 13, 1055087. [Google Scholar] [CrossRef]
- Nowotny, K.; Jung, T.; Höhn, A.; Weber, D.; Grune, T. Advanced glycation end products and oxidative stress in type 2 diabetes mellitus. Biomolecules 2015, 5, 194–222. [Google Scholar] [CrossRef]
- Terao, R.; Ahmed, T.; Suzumura, A.; Terasaki, H. Oxidative Stress-Induced Cellular Senescence in Aging Retina and Age-Related Macular Degeneration. Antioxidants 2022, 11, 2189. [Google Scholar] [CrossRef] [PubMed]
- Nag, T.C.; Maurya, M.; Roy, T.S. Age-Related Changes of the Human Retinal Vessels: Possible Involvement of Lipid Peroxidation. Ann. Anat. 2019, 226, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Chen, Y.; Wang, W.; Wang, Y.; Hu, W.; Shang, X.; Liao, H.; Shi, D.; Huang, Y.; Ha, J.; et al. Association of Retinal Age Gap with Arterial Stiffness and Incident Cardiovascular Disease. Stroke 2022, 53, 3320–3328. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Garcia, S.; Tsuruda, P.R.; Dejda, A.; Ryan, R.D.; Fournier, F.; Chaney, S.Y.; Pilon, F.; Dogan, T.; Cagnone, G.; Patel, P.; et al. Pathological Angiogenesis in Retinopathy Engages Cellular Senescence and Is Amenable to Therapeutic Elimination via BCL-xL Inhibition. Cell Metab. 2021, 33, 818–832.e7. [Google Scholar] [CrossRef] [PubMed]
- Habibi-Kavashkohie, M.R.; Scorza, T.; Oubaha, M. Senescent Cells: Dual Implications on the Retinal Vascular System. Cells 2023, 12, 2341. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Dietenbeck, T.; Giron, A.; Paques, M.; Kachenoura, N.; Girerd, X. Non-Invasive Evaluation of Retinal Vascular Remodeling and Hypertrophy in Humans: Intricate Effect of Ageing, Blood Pressure and Glycaemia. Clin. Res. Cardiol. 2021, 110, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Versari, D.; Daghini, E.; Virdis, A.; Ghiadoni, L.; Taddei, S. Endothelial Dysfunction as a Target for Prevention of Cardiovascular Disease. Diabetes Care 2009, 32 (Suppl. S2), S314–S321. [Google Scholar] [CrossRef] [PubMed]
- Khemais-Benkhiat, S.; Belcastro, E.; Idris-Khodja, N.; Park, S.-H.; Amoura, L.; Abbas, M.; Auger, C.; Kessler, L.; Mayoux, E.; Toti, F.; et al. Angiotensin II-Induced Redox-Sensitive SGLT1 and 2 Expression Promotes High Glucose-Induced Endothelial Cell Senescence. J. Cell Mol. Med. 2020, 24, 2109–2122. [Google Scholar] [CrossRef]
- van der Loo, B.; Fenton, M.J.; Erusalimsky, J.D. Cytochemical Detection of a Senescence-Associated Beta-Galactosidase in Endothelial and Smooth Muscle Cells from Human and Rabbit Blood Vessels. Exp. Cell Res. 1998, 241, 309–315. [Google Scholar] [CrossRef]
- Harvey, A.; Montezano, A.C.; Touyz, R.M. Vascular Biology of Ageing-Implications in Hypertension. J. Mol. Cell Cardiol. 2015, 83, 112–121. [Google Scholar] [CrossRef]
- Shah, A.V.; Bennett, M.R. DNA Damage-Dependent Mechanisms of Ageing and Disease in the Macro- and Microvasculature. Eur. J. Pharmacol. 2017, 816, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, S.; Biswas, S.; Nagrani, N.; Chu, Y.; Chakrabarti, S.; Feng, B. Glucose-Induced Oxidative Stress and Accelerated Aging in Endothelial Cells Are Mediated by the Depletion of Mitochondrial SIRTs. Physiol. Rep. 2020, 8, e14331. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Aroor, A.R.; Jia, C.; Sowers, J.R. Endothelial Cell Senescence in Aging-Related Vascular Dysfunction. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 1802–1809. [Google Scholar] [CrossRef]
- Sapieha, P.; Mallette, F.A. Cellular Senescence in Postmitotic Cells: Beyond Growth Arrest. Trends Cell Biol. 2018, 28, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.-L.; Li, Y. Endothelial Cell Senescence and Age-Related Vascular Diseases. J. Genet. Genom. 2014, 41, 485–495. [Google Scholar] [CrossRef]
- Venkatesh, D.; Fredette, N.; Rostama, B.; Tang, Y.; Vary, C.P.; Liaw, L.; Urs, S. RhoA-mediated signaling in Notch-induced senescence-like growth arrest and endothelial barrier dysfunction. Arter. Thromb. Vasc. Biol. 2011, 31, 876–882. [Google Scholar] [CrossRef] [PubMed]
- Maksin-Matveev, A.; Kanfi, Y.; Hochhauser, E.; Isak, A.; Cohen, H.Y.; Shainberg, A. Sirtuin 6 protects the heart from hypoxic damage. Exp. Cell Res. 2015, 330, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.J.; Kim, N.; Herman, A.B.; Gorospe, M.; Lee, J.-S. Factors and Pathways Modulating Endothelial Cell Senescence in Vascular Aging. Int. J. Mol. Sci. 2022, 23, 10135. [Google Scholar] [CrossRef] [PubMed]
- Fulco, M.; Sartorelli, V. Comparing and Contrasting the Roles of AMPK and SIRT1 in Metabolic Tissues. Cell Cycle 2008, 7, 3669–3679. [Google Scholar] [CrossRef]
- Balaiya, S.; Abu-Amero, K.K.; Kondkar, A.A.; Chalam, K.V. Sirtuins Expression and Their Role in Retinal Diseases. Oxid. Med. Cell Longev. 2017, 2017, 3187594. [Google Scholar] [CrossRef]
- Behringer, E.J. Impact of Aging on Vascular Ion Channels: Perspectives and Knowledge Gaps across Major Organ Systems. Am. J. Physiol. Heart Circ. Physiol. 2023, 325, H1012–H1038. [Google Scholar] [CrossRef] [PubMed]
- Harraz, O.F.; Jensen, L.J. Aging, calcium channel signaling and vascular tone. Mech. Ageing Dev. 2020, 191, 111336. [Google Scholar] [CrossRef]
- DuPont, J.J.; Kim, S.K.; Kenney, R.M.; Jaffe, I.Z. Sex Differences in the Time Course and Mechanisms of Vascular and Cardiac Aging in Mice: Role of the Smooth Muscle Cell Mineralocorticoid Receptor. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H169–H180. [Google Scholar] [CrossRef]
- Ketchem, J.M.; Bowman, E.J.; Isales, C.M. Male sex hormones, aging, and inflammation. Biogerontology 2023, 24. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Zeng, R.; Wan, Q.; Xie, Y.; Liu, X.; An, S.; Jiang, J.; Yang, J.; Zhou, Y.; Shen, X. Aging-Related Increases in Serum Sex Hormone-Binding Globulin Levels in Men Might Be Related to Increased Synthesis. Exp. Gerontol. 2023, 179, 112249. [Google Scholar] [CrossRef] [PubMed]
- Hahad, O.; Frenis, K.; Kuntic, M.; Daiber, A.; Münzel, T. Accelerated Aging and Age-Related Diseases (CVD and Neurological) Due to Air Pollution and Traffic Noise Exposure. Int. J. Mol. Sci. 2021, 22, 2419. [Google Scholar] [CrossRef] [PubMed]
- Kuntic, M.; Kuntic, I.; Hahad, O.; Lelieveld, J.; Münzel, T.; Daiber, A. Impact of Air Pollution on Cardiovascular Aging. Mech. Ageing Dev. 2023, 214, 111857. [Google Scholar] [CrossRef]
- Ansary, T.M.; Hossain, M.R.; Kamiya, K.; Komine, M.; Ohtsuki, M. Inflammatory Molecules Associated with Ultraviolet Radiation-Mediated Skin Aging. Int. J. Mol. Sci. 2021, 22, 3974. [Google Scholar] [CrossRef] [PubMed]
- Panich, U.; Sittithumcharee, G.; Rathviboon, N.; Jirawatnotai, S. Ultraviolet Radiation-Induced Skin Aging: The Role of DNA Damage and Oxidative Stress in Epidermal Stem Cell Damage Mediated Skin Aging. Stem Cells Int. 2016, 2016, 7370642. [Google Scholar] [CrossRef]
- Singhvi, G.R.; Chordiya, A.V. Prednisolone Purpura. J. Assoc. Physicians India 1989, 37, 541–542. [Google Scholar]
- Mahendra, C.K.; Tan, L.T.H.; Pusparajah, P.; Htar, T.T.; Chuah, L.-H.; Lee, V.S.; Low, L.E.; Tang, S.Y.; Chan, K.-G.; Goh, B.H. Detrimental Effects of UVB on Retinal Pigment Epithelial Cells and Its Role in Age-Related Macular Degeneration. Oxid. Med. Cell Longev. 2020, 2020, 1904178. [Google Scholar] [CrossRef] [PubMed]
- Hiramoto, K.; Yamate, Y.; Sato, E.F. Long-Term Ultraviolet A Eye Irradiation Causes Retina Denaturation in Mice. Biomed. Hub. 2017, 2, 1–10. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, J.; Jiang, H.; Yang, X.; Feng, L.; Hu, L.; Wang, L.; Lu, F.; Shen, M. Retinal Microvasculature Alteration in High Myopia. Invest. Ophthalmol. Vis. Sci. 2016, 57, 6020–6030. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.R.; Toychiev, A.; Ablordeppey, R.K.; Srinivas, M.; Benavente-Perez, A. Age Exacerbates the Effect of Myopia on Retinal Capillaries and String Vessels. Front. Med. 2023, 10, 1112396. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Deng, X.; Ye, Y.; Liu, H.; Fang, Y.; Zhang, Z.; Chen, N.; Sun, M.; Shen, L. Morphological Characteristics of Retinal Vessels in Eyes with High Myopia: Ultra-Wide Field Images Analyzed by Artificial Intelligence Using a Transfer Learning System. Front. Med. 2022, 9, 956179. [Google Scholar] [CrossRef]
- Emeterio Nateras, O.S.; Harrison, J.M.; Muir, E.R.; Zhang, Y.; Peng, Q.; Chalfin, S.; Gutierrez, J.E.; Johnson, D.A.; Kiel, J.W.; Duong, T.Q. Choroidal Blood Flow Decreases with Age: An MRI Study. Curr. Eye Res. 2014, 39, 1059–1067. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lin, Y.; Jiang, H.; Liu, Y.; Rosa Gameiro, G.; Gregori, G.; Dong, C.; Rundek, T.; Wang, J. Age-Related Alterations in Retinal Tissue Perfusion and Volumetric Vessel Density. Invest. Ophthalmol. Vis. Sci. 2019, 60, 685–693. [Google Scholar] [CrossRef]
- Benavente-Perez, A. Evidence of Vascular Involvement in Myopia: A Review. Front. Med. 2023, 10, 1112996. [Google Scholar] [CrossRef] [PubMed]
- Tyrrell, D.J.; Blin, M.G.; Song, J.; Wood, S.C.; Zhang, M.; Beard, D.A.; Goldstein, D.R. Age-Associated Mitochondrial Dysfunction Accelerates Atherogenesis. Circ. Res. 2020, 126, 298–314. [Google Scholar] [CrossRef]
- Tyrrell, D.J.; Blin, M.G.; Song, J.; Wood, S.C.; Goldstein, D.R. Aging Impairs Mitochondrial Function and Mitophagy and Elevates Interleukin 6 within the Cerebral Vasculature. J. Am. Heart Assoc. 2020, 9, e017820. [Google Scholar] [CrossRef]
- Akpek, E.K.; Smith, R.A. Overview of Age-Related Ocular Conditions. Am. J. Manag. Care 2013, 19, S67–S75. [Google Scholar]
- Choi, Y.K. An Altered Neurovascular System in Aging-Related Eye Diseases. Int. J. Mol. Sci. 2022, 23, 14104. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, S.; Xie, B.; Zhong, Y. Aging, Cellular Senescence, and Glaucoma. Aging Dis. 2024, 15, 546–564. [Google Scholar] [CrossRef]
- Izzotti, A.; Bagnis, A.; Saccà, S.C. The role of oxidative stress in glaucoma. Mutat. Res. 2006, 612, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Türksever, C.; Todorova, M.G. Peripapillary Oxygenation and Retinal Vascular Responsiveness to Flicker Light in Primary Open Angle Glaucoma. Metabolites 2022, 12, 597. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhao, Q.; Li, M.; Duan, Q.; Zhao, Y.; Zhang, H. The Effects of Endothelium-Specific CYP2J2 Overexpression on the Attenuation of Retinal Ganglion Cell Apoptosis in a Glaucoma Rat Model. FASEB J. 2019, 33, 11194–11209. [Google Scholar] [CrossRef]
- Romano, F.; Lamanna, F.; Gabrielle, P.H.; Teo, K.Y.C.; Battaglia Parodi, M.; Iacono, P.; Fraser-Bell, S.; Cornish, E.E.; Nassisi, M.; Viola, F.; et al. Update on Retinal Vein Occlusion. Asia Pac. J. Ophthalmol. 2023, 12, 196–210. [Google Scholar] [CrossRef] [PubMed]
- Greaves, M. Aging and the Pathogenesis of Retinal Vein Thrombosis. Br. J. Ophthalmol. 1997, 81, 810–811. [Google Scholar] [CrossRef][Green Version]
- Jung, S.H.; Kim, K.-A.; Sohn, S.W.; Yang, S.J. Association of Aqueous Humor Cytokines with the Development of Retinal Ischemia and Recurrent Macular Edema in Retinal Vein Occlusion. Invest. Ophthalmol. Vis. Sci. 2014, 55, 2290–2296. [Google Scholar] [CrossRef]
- Tyrrell, D.J.; Goldstein, D.R. Ageing and Atherosclerosis: Vascular Intrinsic and Extrinsic Factors and Potential Role of IL-6. Nat. Rev. Cardiol. 2021, 18, 58–68. [Google Scholar] [CrossRef]
- Ortiz-Montero, P.; Londoño-Vallejo, A.; Vernot, J.P. Senescence-associated IL-6 and IL-8 cytokines induce a self- and cross-reinforced senescence/inflammatory milieu strengthening tumorigenic capabilities in the MCF-7 breast cancer cell line. Cell Commun. Signal. 2017, 15, 17. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Cheng, Y.; Wang, S.; Wang, Y.; Liu, P.; Wu, H. Review: The Development of Risk Factors and Cytokines in Retinal Vein Occlusion. Front. Med. 2022, 9, 910600. [Google Scholar] [CrossRef] [PubMed]
- Yabluchanskiy, A.; Ma, Y.; Chiao, Y.A.; Lopez, E.F.; Voorhees, A.P.; Toba, H.; Hall, M.E.; Han, H.-C.; Lindsey, M.L.; Jin, Y.-F. Cardiac Aging Is Initiated by Matrix Metalloproteinase-9-Mediated Endothelial Dysfunction. Am. J. Physiol. Heart Circ. Physiol. 2014, 306, H1398–H1407. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Huang, X.; Dou, L.; Yan, M.; Shen, T.; Tang, W.; Li, J. Aging and Aging-Related Diseases: From Molecular Mechanisms to Interventions and Treatments. Signal Transduct. Target. Ther. 2022, 7, 391. [Google Scholar] [CrossRef] [PubMed]
- Buonfiglio, F.; Böhm, E.W.; Pfeiffer, N.; Gericke, A. Oxidative Stress: A Suitable Therapeutic Target for Optic Nerve Diseases? Antioxidants 2023, 12, 1465. [Google Scholar] [CrossRef] [PubMed]
- Cernadas, M.R.; Sánchez de Miguel, L.; García-Durán, M.; González-Fernández, F.; Millás, I.; Montón, M.; Rodrigo, J.; Rico, L.; Fernández, P.; de Frutos, T.; et al. Expression of Constitutive and Inducible Nitric Oxide Synthases in the Vascular Wall of Young and Aging Rats. Circ. Res. 1998, 83, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Wareham, L.K.; Buys, E.S.; Sappington, R.M. The nitric oxide-guanylate cyclase pathway and glaucoma. Nitric Oxide. 2018, 77, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.K.; Kim, Y.-M. Regulation of Endothelial and Vascular Functions by Carbon Monoxide via Crosstalk with Nitric Oxide. Front. Cardiovasc. Med. 2021, 8, 649630. [Google Scholar] [CrossRef]
- Alp, N.J.; Channon, K.M. Regulation of Endothelial Nitric Oxide Synthase by Tetrahydrobiopterin in Vascular Disease. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 413–420. [Google Scholar] [CrossRef]
- Janaszak-Jasiecka, A.; Płoska, A.; Wierońska, J.M.; Dobrucki, L.W.; Kalinowski, L. Endothelial Dysfunction Due to eNOS Uncoupling: Molecular Mechanisms as Potential Therapeutic Targets. Cell Mol. Biol. Lett. 2023, 28, 21. [Google Scholar] [CrossRef]
- Mortensen, A.; Lykkesfeldt, J. Does Vitamin C Enhance Nitric Oxide Bioavailability in a Tetrahydrobiopterin-Dependent Manner? In Vitro, in Vivo and Clinical Studies. Nitric Oxide 2014, 36, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Olsson, A.K.; Dimberg, A.; Kreuger, J.; Claesson-Welsh, L. VEGF receptor signalling—in control of vascular function. Nat. Rev. Mol. Cell Biol. 2006, 7, 359–371. [Google Scholar] [CrossRef]
- Alfaras, I.; Di Germanio, C.; Bernier, M.; Csiszar, A.; Ungvari, Z.; Lakatta, E.G.; de Cabo, R. Pharmacological Strategies to Retard Cardiovascular Aging. Circ. Res. 2016, 118, 1626–1642. [Google Scholar] [CrossRef]
- Costantino, S.; Paneni, F.; Cosentino, F. Ageing, Metabolism and Cardiovascular Disease. J. Physiol. 2016, 594, 2061–2073. [Google Scholar] [CrossRef]
- Haga, M.; Okada, M. Systems Approaches to Investigate the Role of NF-κB Signaling in Aging. Biochem. J. 2022, 479, 161–183. [Google Scholar] [CrossRef]
- Suh, J.H.; Shenvi, S.V.; Dixon, B.M.; Liu, H.; Jaiswal, A.K.; Liu, R.-M.; Hagen, T.M. Decline in Transcriptional Activity of Nrf2 Causes Age-Related Loss of Glutathione Synthesis, Which Is Reversible with Lipoic Acid. Proc. Natl. Acad. Sci. USA 2004, 101, 3381–3386. [Google Scholar] [CrossRef]
- Koriyama, Y.; Nakayama, Y.; Matsugo, S.; Kato, S. Protective Effect of Lipoic Acid against Oxidative Stress Is Mediated by Keap1/Nrf2-Dependent Heme Oxygenase-1 Induction in the RGC-5 Cellline. Brain Res. 2013, 1499, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Inman, D.M.; Lambert, W.S.; Calkins, D.J.; Horner, P.J. α-Lipoic Acid Antioxidant Treatment Limits Glaucoma-Related Retinal Ganglion Cell Death and Dysfunction. PLoS ONE 2013, 8, e65389. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shen, H.; Shi, J.; Tang, L. The Effects of Alpha Lipoic Acid in Preventing Oxidative Stress-Induced Retinal Pigment Epithelial Cell Injury. Can. J. Physiol. Pharmacol. 2014, 92, 765–772. [Google Scholar] [CrossRef]
- Gioscia-Ryan, R.A.; LaRocca, T.J.; Sindler, A.L.; Zigler, M.C.; Murphy, M.P.; Seals, D.R. Mitochondria-Targeted Antioxidant (MitoQ) Ameliorates Age-Related Arterial Endothelial Dysfunction in Mice. J. Physiol. 2014, 592, 2549–2561. [Google Scholar] [CrossRef]
- Pearson, K.J.; Baur, J.A.; Lewis, K.N.; Peshkin, L.; Price, N.L.; Labinskyy, N.; Swindell, W.R.; Kamara, D.; Minor, R.K.; Perez, E.; et al. Resveratrol Delays Age-Related Deterioration and Mimics Transcriptional Aspects of Dietary Restriction without Extending Life Span. Cell Metab. 2008, 8, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Ungvari, Z.; Orosz, Z.; Labinskyy, N.; Rivera, A.; Xiangmin, Z.; Smith, K.; Csiszar, A. Increased Mitochondrial H2O2 Production Promotes Endothelial NF-κB Activation in Aged Rat Arteries. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H37–H47. [Google Scholar] [CrossRef] [PubMed]
- Rice, K.M.; Preston, D.L.; Walker, E.M.; Blough, E.R. Aging Influences Multiple Incidices of Oxidative Stress in the Aortic Media of the Fischer 344/NNiaxBrown Norway/BiNia Rat. Free Radic. Res. 2006, 40, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.; Jang, K.-H.; Koo, T.-S.; Park, C.; Kim, Y.-H.; Lee, J.; Kim, E. Protective Effects of PARP1-Inhibitory Compound in Dry Age-Related Macular Degeneration. Biomed. Pharmacother. 2021, 133, 111041. [Google Scholar] [CrossRef] [PubMed]
- Gano, L.B.; Donato, A.J.; Pasha, H.M.; Hearon, C.M.; Sindler, A.L.; Seals, D.R. The SIRT1 Activator SRT1720 Reverses Vascular Endothelial Dysfunction, Excessive Superoxide Production, and Inflammation with Aging in Mice. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H1754–H1763. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Luo, Z.; Jin, M.; Sheng, W.; Wang, H.-T.; Long, X.; Wu, Y.; Hu, P.; Xu, H.; Zhang, X. Exploration of Age-Related Mitochondrial Dysfunction and the Anti-Aging Effects of Resveratrol in Zebrafish Retina. Aging 2019, 11, 3117–3137. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhou, L.; Yang, Y.; Liu, Y. Cycloastragenol: An Exciting Novel Candidate for Age-Associated Diseases. Exp. Ther. Med. 2018, 16, 2175–2182. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human Telomere Biology: A Contributory and Interactive Factor in Aging, Disease Risks, and Protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Chiang, J.Y.; Choi, J.Y.; Xiong, Z.-M.; Mao, X.; Collins, F.S.; Hodes, R.J.; Cao, K. Transient Induction of Telomerase Expression Mediates Senescence and Reduces Tumorigenesis in Primary Fibroblasts. Proc. Natl. Acad. Sci. USA 2019, 116, 18983–18993. [Google Scholar] [CrossRef]
- Roos, C.M.; Zhang, B.; Palmer, A.K.; Ogrodnik, M.B.; Pirtskhalava, T.; Thalji, N.M.; Hagler, M.; Jurk, D.; Smith, L.A.; Casaclang-Verzosa, G.; et al. Chronic Senolytic Treatment Alleviates Established Vasomotor Dysfunction in Aged or Atherosclerotic Mice. Aging Cell 2016, 15, 973–977. [Google Scholar] [CrossRef]
- Nieto, M.; Könisgberg, M.; Silva-Palacios, A. Quercetin and Dasatinib, Two Powerful Senolytics in Age-Related Cardiovascular Disease. Biogerontology 2024, 25, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Lin, S.; Copland, D.A.; Dick, A.D.; Liu, J. Cellular Senescence in the Aging Retina and Developments of Senotherapies for Age-Related Macular Degeneration. J. Neuroinflamm. 2021, 18, 32. [Google Scholar] [CrossRef] [PubMed]
- Suminska-Jasinska, K.; Polubinska, A.; Ciszewicz, M.; Mikstacki, A.; Antoniewicz, A.; Breborowicz, A. Sulodexide Reduces Senescence-Related Changes in Human Endothelial Cells. Med. Sci. Monit. 2011, 17, CR222–CR226. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.M.; Battson, M.L.; Jarrell, D.K.; Hou, S.; Ecton, K.E.; Weir, T.L.; Gentile, C.L. SGLT2 Inhibition via Dapagliflozin Improves Generalized Vascular Dysfunction and Alters the Gut Microbiota in Type 2 Diabetic Mice. Cardiovasc. Diabetol. 2018, 17, 62. [Google Scholar] [CrossRef] [PubMed]
- Tai, S.; Zhou, Y.; Fu, L.; Ding, H.; Zhou, Y.; Yin, Z.; Yang, R.; Liu, Z.; Zhou, S. Dapagliflozin impedes endothelial cell senescence by activating the SIRT1 signaling pathway in type 2 diabetes. Heliyon 2023, 9, e19152. [Google Scholar] [CrossRef] [PubMed]
- Böhm, E.W.; Buonfiglio, F.; Voigt, A.M.; Bachmann, P.; Safi, T.; Pfeiffer, N.; Gericke, A. Oxidative Stress in the Eye and Its Role in the Pathophysiology of Ocular Diseases. Redox Biol. 2023, 68, 102967. [Google Scholar] [CrossRef] [PubMed]
- Liao, D.S.; Grossi, F.V.; El Mehdi, D.; Gerber, M.R.; Brown, D.M.; Heier, J.S.; Wykoff, C.C.; Singerman, L.J.; Abraham, P.; Grassmann, F.; et al. Complement C3 Inhibitor Pegcetacoplan for Geographic Atrophy Secondary to Age-Related Macular Degeneration: A Randomized Phase 2 Trial. Ophthalmology 2020, 127, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, G.J.; Westby, K.; Csaky, K.G.; Monés, J.; Pearlman, J.A.; Patel, S.S.; Joondeph, B.C.; Randolph, J.; Masonson, H.; Rezaei, K.A. C5 Inhibitor Avacincaptad Pegol for Geographic Atrophy Due to Age-Related Macular Degeneration: A Randomized Pivotal Phase 2/3 Trial. Ophthalmology 2021, 128, 576–586. [Google Scholar] [CrossRef]
- Qin, S.; Dong, N.; Yang, M.; Wang, J.; Feng, X.; Wang, Y. Complement Inhibitors in Age-Related Macular Degeneration: A Potential Therapeutic Option. J. Immunol. Res. 2021, 2021, 9945725. [Google Scholar] [CrossRef]
- Kolosova, N.G.; Muraleva, N.A.; Zhdankina, A.A.; Stefanova, N.A.; Fursova, A.Z.; Blagosklonny, M.V. Prevention of Age-Related Macular Degeneration-like Retinopathy by Rapamycin in Rats. Am. J. Pathol. 2012, 181, 472–477. [Google Scholar] [CrossRef]
- Niu, Z.; Shi, Y.; Li, J.; Qiao, S.; Du, S.; Chen, L.; Tian, H.; Wei, L.; Cao, H.; Wang, J.; et al. Protective Effect of Rapamycin in Models of Retinal Degeneration. Exp. Eye Res. 2021, 210, 108700. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathophysiological Trigger | Pathomechanism | Refs. |
|---|---|---|
| Hyperglycemia | retinal vascular growth and aging, with increased β-galactosidase activity, p53 expression, elevated IL-6 and VEGF | [88,89,90,96] |
| Hypertension | retinal microvascular remodeling | [96] |
| Oxidative stress | pivotal factor in vascular endothelial cell dysfunction and subsequent damage, exacerbating aging process, cell death and DNA damage in several ocular disorders | [51,101] |
| Mitochondrial dysfunction | reduced mitochondrial biogenesis and SIRT activity | [102,103,129] |
| Telomere alteration | telomere shortening and damage to DNA | [17,103] |
| Ca2+ signaling anomalies | remodeling processes induce impairment in Ca2+ signals, impacting vascular function | [112] |
| Angiogenetic dysregulation, endothelial dysfunction | accumulation of genetic damage and epigenetic changes affecting gene expression, leads to vascular impairment | [103,104,105,133,137,138] |
| Vascular obstruction | aging may exacerbate the pro-inflammatory activity of IL-6, IL-8, and MMP-9, all agents related to retinal vascular occlusion | [139,140,141,142,143] |
| Androgen decline | enhanced synthesis of the serum sex hormone-binding globulin may lead to a reduced level of androgens, which physiologically exert an anti-inflammatory effect | [114,115] |
| Traffic noise and air pollution exposure | -Traffic noise exposure: alteration in stress hormone levels and heart rate as well as establishment of neuroinflammation. -Air pollution: ultrafine particles induce immune reactions and ROS formation.collectively, these factors trigger mitochondrial dysfunction, oxidative stress, telomere shortening, and chronic inflammation | [116,117] |
| UV exposure | raised ROS formation, and subsequently elevated VEGF, MMP-2, and MMP-9, causing neoangiogenesis and degradation of ECM | [122] |
| High myopia | age-related decreased tissue perfusion and loss of endothelial cells, aggravating the reduced ocular blood flow in high myopia | [68,82,126,127] |
| Mechanistic Target | Candidate Molecule | Refs. |
|---|---|---|
| eNOS modulation | BH4 | [150,151] |
| Mitochondria | MitoQ | [160] |
| tetrapeptide SS-31 | [76] | |
| Nrf2 activation | α-lipoic acid | [156,157,158,159] |
| SIRT1 activation | PARP-1 inhibitors | [164] |
| resveratrol | [166] | |
| SRT1720 | [165] | |
| dapagliflozin | [174,175] | |
| mTOR inhibition | rapamycin | [180,181] |
| Telomerase activation | cycloastraganol | [169] |
| Senolytic | dasatinib | [170] |
| quercetin | [170] | |
| Bcl-2 inhibitors | [172] | |
| Sulodexide | [88,173] | |
| Complement inhibition | pegcetacoplan | [177] |
| avacincaptad pegol | [178] | |
| NGM621, AAVCAGsCD59, IBI302 | [179] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, X.; Buonfiglio, F.; Pfeiffer, N.; Gericke, A. Aging in Ocular Blood Vessels: Molecular Insights and the Role of Oxidative Stress. Biomedicines 2024, 12, 817. https://doi.org/10.3390/biomedicines12040817
Cui X, Buonfiglio F, Pfeiffer N, Gericke A. Aging in Ocular Blood Vessels: Molecular Insights and the Role of Oxidative Stress. Biomedicines. 2024; 12(4):817. https://doi.org/10.3390/biomedicines12040817
Chicago/Turabian StyleCui, Xiuting, Francesco Buonfiglio, Norbert Pfeiffer, and Adrian Gericke. 2024. "Aging in Ocular Blood Vessels: Molecular Insights and the Role of Oxidative Stress" Biomedicines 12, no. 4: 817. https://doi.org/10.3390/biomedicines12040817
APA StyleCui, X., Buonfiglio, F., Pfeiffer, N., & Gericke, A. (2024). Aging in Ocular Blood Vessels: Molecular Insights and the Role of Oxidative Stress. Biomedicines, 12(4), 817. https://doi.org/10.3390/biomedicines12040817