
Progastrin: An Overview of Its Crucial Role in the Tumorigenesis of Gastrointestinal Cancers

 , ,
, ,  , , ,
, , , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. hPG80 Gene and Receptor
2.1. Wnt/β-Catenin Signaling Pathway
2.2. K-Ras, MEK-ERK Signaling Pathways
2.3. PI3K/Akt, NF-κB, SMAD4
2.4. Progastrin Receptor
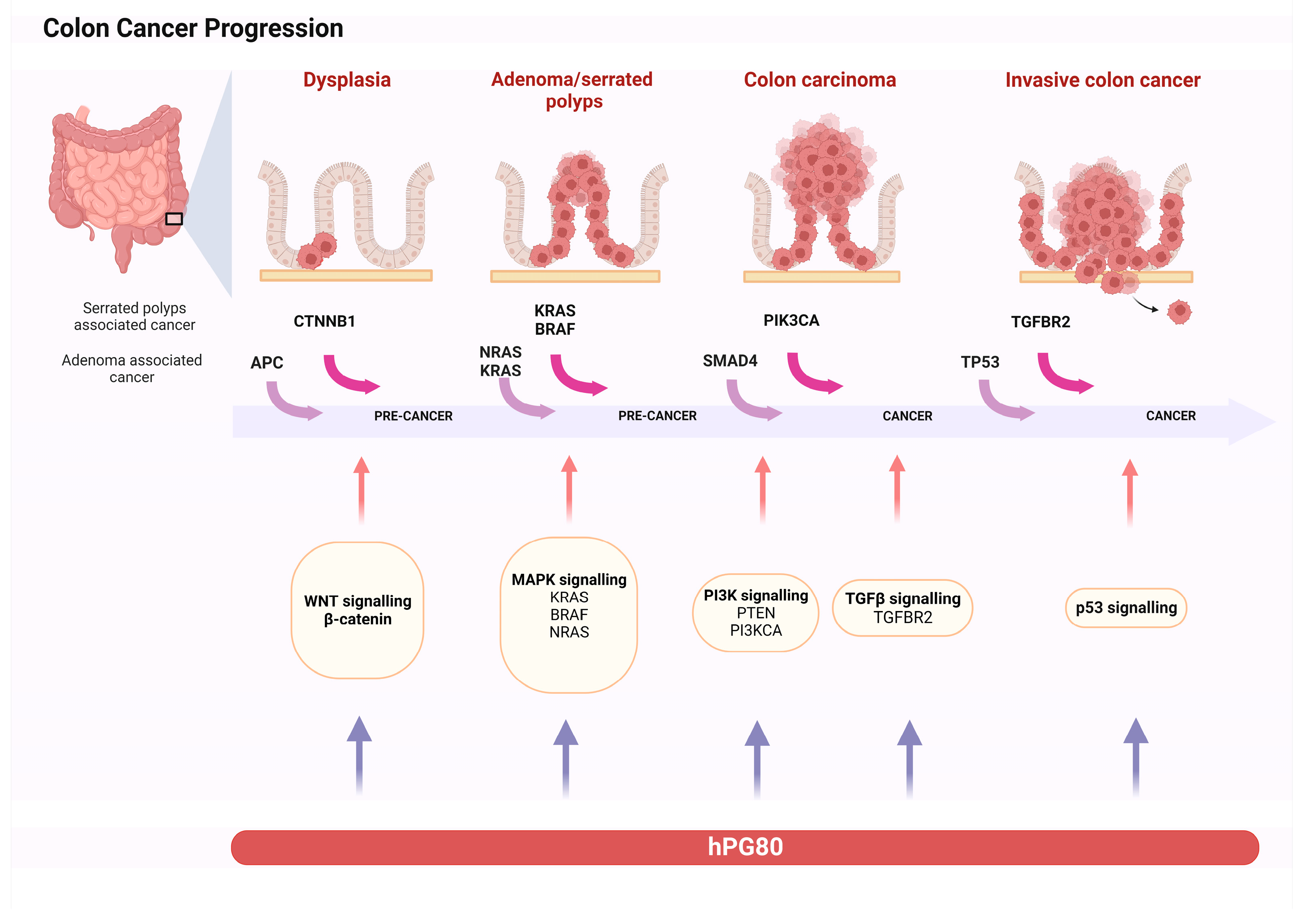
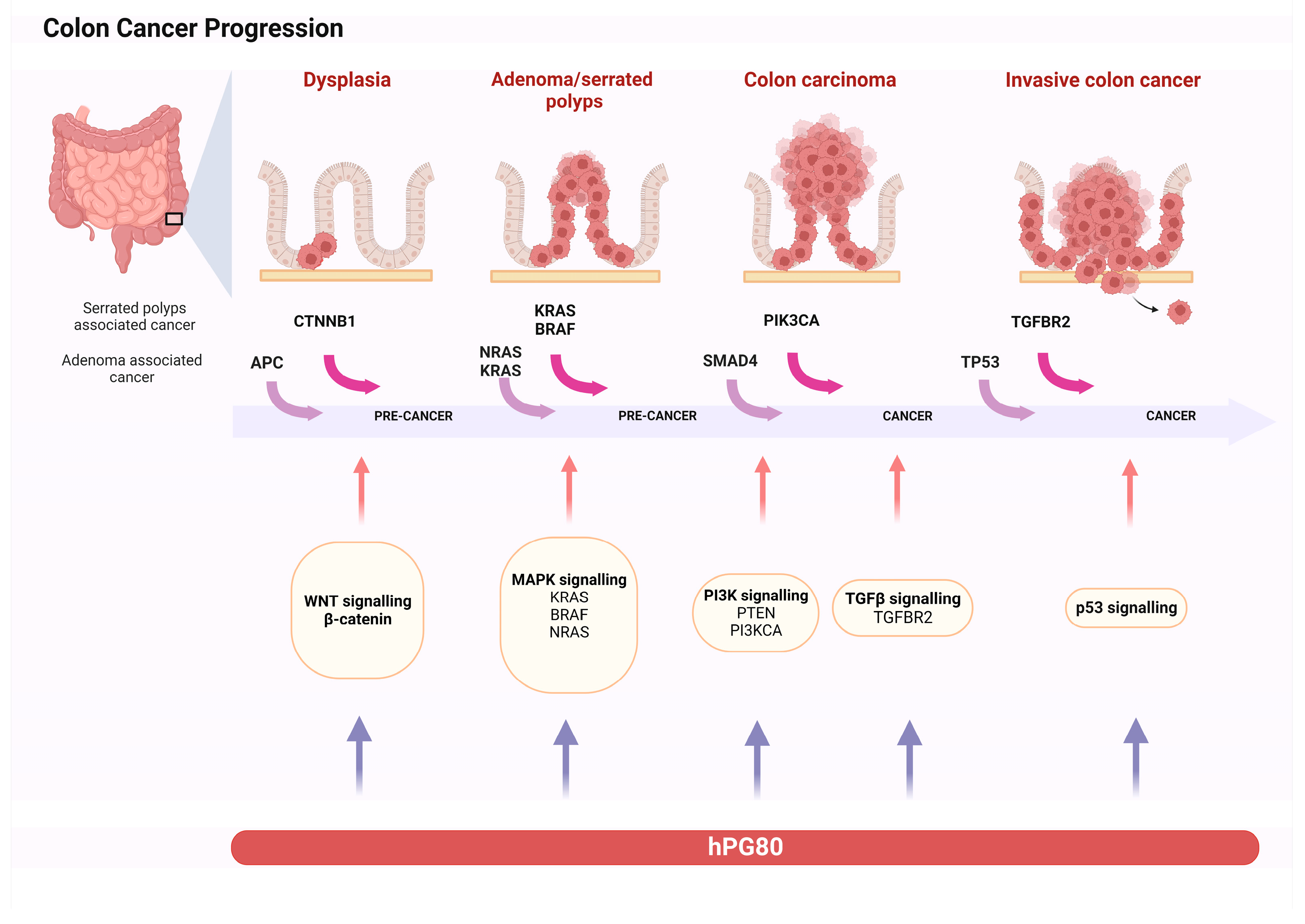
3. Progastrin Role on Colorectal Cancer Cells
3.1. Progastrin as a Factor in Tumorigenesis
3.2. Progastrin Is Essential for the Survival of Tumor Stem Cells
3.3. Progastrin Decreases Apoptosis
3.4. Progastrin Is a Pro-Angiogenic Factor
3.5. Progastrin Regulates Adhesions and Tight Junctions
3.6. Progastrin and Tumor-Reactive Stroma
4. Progastrin Role in Other Gi Cancers
4.1. Progastrin and Gastric Cancer
4.2. Progastrin and Liver
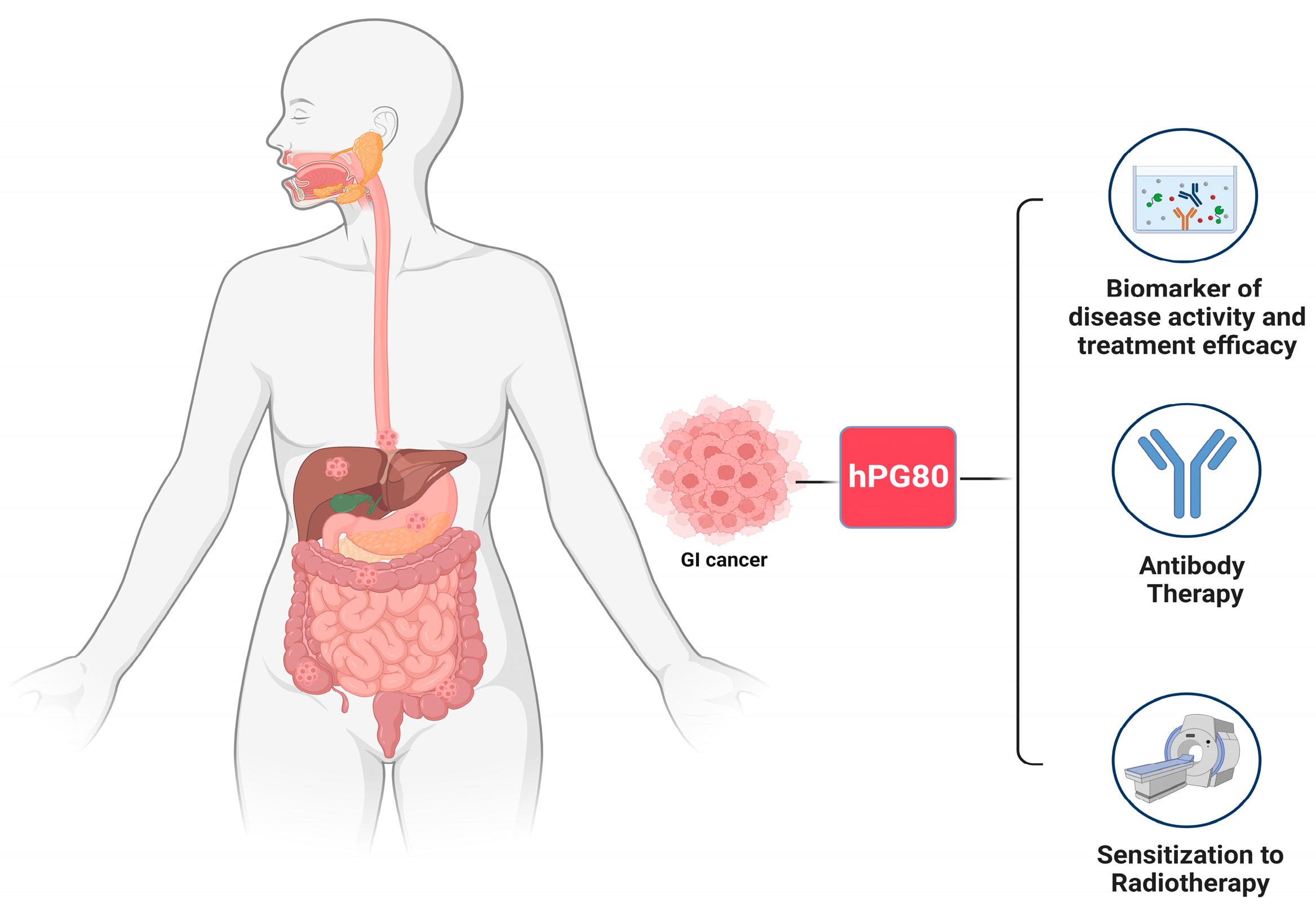
5. Clinical Implications
5.1. Monitoring Disease Activity and Treatment Efficacy
5.2. Antibody Therapy
5.3. Sensitization to Radiotherapy
5.4. Clinical Trials
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Duan, S.; Rico, K.; Merchant, J.L. Gastrin: From Physiology to Gastrointestinal Malignancies. Function 2021, 3, zqab062. [Google Scholar] [CrossRef]
- MacNicol, J.L.; Pearson, W. Gastrin and Nitric Oxide Production in Cultured Gastric Antral Mucosa Are Altered in Response to a Gastric Digest of a Dietary Supplement. Front. Vet. Sci. 2021, 8, 684203. [Google Scholar] [CrossRef]
- Zeng, Q.; Ou, L.; Wang, W.; Guo, D.-Y. Gastrin, Cholecystokinin, Signaling, and Biological Activities in Cellular Processes. Front. Endocrinol. 2020, 11, 112. [Google Scholar] [CrossRef] [PubMed]
- Kohli, M.; Tan, W.; Vire, B.; Liaud, P.; Blairvacq, M.; Berthier, F.; Rouison, D.; Garnier, G.; Payen, L.; Cousin, T.; et al. Prognostic Value of Plasma HPG80 (Circulating Progastrin) in Metastatic Renal Cell Carcinoma. Cancers 2021, 13, 375. [Google Scholar] [CrossRef] [PubMed]
- Dupuy, M.; Iltache, S.; Rivière, B.; Prieur, A.; Pageaux, G.P.; Bedoya, J.U.; Faure, S.; Guillaumée, H.; Assenat, E. Plasma HPG80 (Circulating Progastrin) as a Novel Prognostic Biomarker for Hepatocellular Carcinoma. Cancers 2022, 14, 402. [Google Scholar] [CrossRef] [PubMed]
- You, B.; Mercier, F.; Assenat, E.; Langlois-Jacques, C.; Glehen, O.; Soulé, J.; Payen, L.; Kepenekian, V.; Dupuy, M.; Belouin, F.; et al. The Oncogenic and Druggable HPG80 (Progastrin) Is Overexpressed in Multiple Cancers and Detected in the Blood of Patients. EBioMedicine 2020, 51, 102574. [Google Scholar] [CrossRef]
- Bardram, L. Progastrin in Serum from Zollinger-Ellison Patients. An Indicator of Malignancy? Gastroenterology 1990, 98, 1420–1426. [Google Scholar] [CrossRef]
- Siddheshwar, R.K.; Gray, J.C.; Kelly, S.B. Plasma Levels of Progastrin but Not Amidated Gastrin or Glycine Extended Gastrin Are Elevated in Patients with Colorectal Carcinoma. Gut 2001, 48, 47–52. [Google Scholar] [CrossRef]
- Singh, P.; Xu, Z.; Dai, B.; Rajaraman, S.; Rubin, N.; Dhruva, B. Incomplete Processing of Progastrin Expressed by Human Colon Cancer Cells: Role of Noncarboxyamidated Gastrins. Am. J. Physiol. 1994, 266, G459–G468. [Google Scholar] [CrossRef]
- Prieur, A.; Joubert, D. Abstract LB-127: Progastrin: A Specific Early Cancer Screening Biomarker and a Breakthrough Innovative Cancer Target. Cancer Res. 2017, 77, LB-127. [Google Scholar] [CrossRef]
- Konturek, P.C.; Bielanski, W.; Konturek, S.J.; Hartwich, A.; Pierzchalski, P.; Gonciarz, M.; Marlicz, K.; Starzynska, T.; Zuchowicz, M.; Darasz, Z.; et al. Progastrin and Cyclooxygenase-2 in Colorectal Cancer. Dig. Dis. Sci. 2002, 47, 1984–1991. [Google Scholar] [CrossRef] [PubMed]
- Lund, T.; Geurts van Kessel, A.H.M.; Haun, S.; Dixon, J.E. The Genes for Human Gastrin and Cholecystokinin Are Located on Different Chromosomes. Hum. Genet. 1986, 73, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Owlia, A.; Varro, A.; Dai, B.; Rajaraman, S.; Wood, T. Gastrin Gene Expression Is Required for the Proliferation and Tumorigenicity of Human Colon Cancer Cells. Cancer Res. 1996, 56, 4111–4115. [Google Scholar] [PubMed]
- Zhao, H.; Ming, T.; Tang, S.; Ren, S.; Yang, H.; Liu, M.; Tao, Q.; Xu, H. Wnt Signaling in Colorectal Cancer: Pathogenic Role and Therapeutic Target. Mol. Cancer 2022, 21, 144. [Google Scholar] [CrossRef] [PubMed]
- Chakladar, A.; Dubeykovskiy, A.; Wojtukiewicz, L.J.; Pratap, J.; Lei, S.; Wang, T.C. Synergistic Activation of the Murine Gastrin Promoter by Oncogenic Ras and Beta-Catenin Involves SMAD Recruitment. Biochem. Biophys. Res. Commun. 2005, 336, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Maharati, A.; Moghbeli, M. PI3K/AKT Signaling Pathway as a Critical Regulator of Epithelial-Mesenchymal Transition in Colorectal Tumor Cells. Cell Commun. Signal 2023, 21, 201. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Sun, M.M.; Zhang, G.G.; Yang, J.; Chen, K.S.; Xu, W.W.; Li, B. Targeting PI3K/Akt Signal Transduction for Cancer Therapy. Signal Transduct. Target. Ther. 2021, 6, 425. [Google Scholar] [CrossRef]
- Itoh, N.; Semba, S.; Ito, M.; Takeda, H.; Kawata, S.; Yamakawa, M. Phosphorylation of Akt/PKB Is Required for Suppression of Cancer Cell Apoptosis and Tumor Progression in Human Colorectal Carcinoma. Cancer 2002, 94, 3127–3134. [Google Scholar] [CrossRef]
- Ferrand, A.; Bertrand, C.; Portolan, G.; Cui, G.; Carlson, J.; Pradayrol, L.; Fourmy, D.; Dufresne, M.; Wang, T.C.; Seva, C. Signaling Pathways Associated with Colonic Mucosa Hyperproliferation in Mice Overexpressing Gastrin Precursors. Cancer Res. 2005, 65, 2770–2777. [Google Scholar] [CrossRef]
- Dubeykovskiy, A.; Nguyen, T.; Dubeykovskaya, Z.; Lei, S.; Wang, T.C. Flow Cytometric Detection of Progastrin Interaction with Gastrointestinal Cells. Regul. Pept. 2008, 151, 106–114. [Google Scholar] [CrossRef]
- Sarkar, S.; Swiercz, R.; Kantara, C.; Hajjar, K.A.; Singh, P. Annexin A2 Mediates Up-Regulation of NF-ΚB, β-Catenin, and Stem Cell in Response to Progastrin in Mice and HEK-293 Cells. Gastroenterology 2011, 140, 583–595.e4. [Google Scholar] [CrossRef]
- Han, F.; Shrestha, S.; Huang, H.; Lv, H.-Y.; Nie, C.; Lin, L.; Lu, M.-L. Expression of Annexin II in Gastric Carcinoma and Its Role in Gastric Cancer Metastasis. World J. Gastroenterol. 2017, 23, 7009–7015. [Google Scholar] [CrossRef]
- Jin, G.; Sakitani, K.; Wang, H.; Jin, Y.; Dubeykovskiy, A.; Worthley, D.L.; Tailor, Y.; Wang, T.C. The G-Protein Coupled Receptor 56, Expressed in Colonic Stem and Cancer Cells, Binds Progastrin to Promote Proliferation and Carcinogenesis. Oncotarget 2017, 8, 40606–40619. [Google Scholar] [CrossRef] [PubMed]
- Fearon, E.R.; Vogelstein, B. A Genetic Model for Colorectal Tumorigenesis. Cell 1990, 61, 759–767. [Google Scholar] [CrossRef]
- Sarantis, P.; Trifylli, E.M.; Koustas, E.; Papavassiliou, K.A.; Karamouzis, M.V.; Papavassiliou, A.G. Immune Microenvironment and Immunotherapeutic Management in Virus-Associated Digestive System Tumors. Int. J. Mol. Sci. 2022, 23, 13612. [Google Scholar] [CrossRef] [PubMed]
- Bader, J.E.; Voss, K.; Rathmell, J.C. Targeting Metabolism to Improve the Tumor Microenvironment for Cancer Immunotherapy. Mol. Cell 2020, 78, 1019–1033. [Google Scholar] [CrossRef] [PubMed]
- Hollande, F.; Imdahl, A.; Mantamadiotis, T.; Ciccotosto, G.D.; Shulkes, A.; Baldwin, G.S. Glycine-Extended Gastrin Acts as an Autocrine Growth Factor in a Nontransformed Colon Cell Line. Gastroenterology 1997, 113, 1576–1588. [Google Scholar] [CrossRef]
- Stepan, V.M.; Krametter, D.F.; Matsushima, M.; Todisco, A.; Delvalle, J.; Dickinson, C.J. Glycine-Extended Gastrin Regulates HEK Cell Growth. Am. J. Physiol. 1999, 277, R572–R581. [Google Scholar] [CrossRef]
- Wang, T.C.; Koh, T.J.; Varro, A.; Cahill, R.J.; Dangler, C.A.; Fox, J.G.; Dockray, G.J. Processing and Proliferative Effects of Human Progastrin in Transgenic Mice. J. Clin. Investig. 1996, 98, 1918–1929. [Google Scholar] [CrossRef]
- Singh, P.; Velasco, M.; Given, R.; Wargovich, M.; Varro, A.; Wang, T.C. Mice Overexpressing Progastrin Are Predisposed for Developing Aberrant Colonic Crypt Foci in Response to AOM. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 278, G390–G399. [Google Scholar] [CrossRef]
- Koh, T.J.; Bulitta, C.J.; Fleming, J.V.; Dockray, G.J.; Varro, A.; Wang, T.C. Gastrin Is a Target of the Beta-Catenin/TCF-4 Growth-Signaling Pathway in a Model of Intestinal Polyposis. J. Clin. Investig. 2000, 106, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Pannequin, J.; Delaunay, N.; Buchert, M.; Surrel, F.; Bourgaux, J.-F.; Ryan, J.; Boireau, S.; Coelho, J.; Pélegrin, A.; Singh, P.; et al. Beta-Catenin/Tcf-4 Inhibition after Progastrin Targeting Reduces Growth and Drives Differentiation of Intestinal Tumors. Gastroenterology 2007, 133, 1554–1568. [Google Scholar] [CrossRef] [PubMed]
- Prieur, A.; Cappellini, M.; Habif, G.; Lefranc, M.-P.; Mazard, T.; Morency, E.; Pascussi, J.-M.; Flacelière, M.; Cahuzac, N.; Vire, B.; et al. Targeting the Wnt Pathway and Cancer Stem Cells with Anti-Progastrin Humanized Antibodies as a Potential Treatment for K-RAS-Mutated Colorectal Cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2017, 23, 5267–5280. [Google Scholar] [CrossRef]
- Ferrand, A.; Sandrin, M.S.; Shulkes, A.; Baldwin, G.S. Expression of Gastrin Precursors by CD133-Positive Colorectal Cancer Cells Is Crucial for Tumour Growth. Biochim. Et Biophys. Acta 2009, 1793, 477–488. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, L.; Li, C.; Qiu, Z.; Kuang, T.; Wu, Z.; Deng, W. Effects of Hormones on Intestinal Stem Cells. Stem Cell Res. Ther. 2023, 14, 105. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Sarkar, S.; Kantara, C.; Maxwell, C. Progastrin Peptides Increase the Risk of Developing Colonic Tumors: Impact on Colonic Stem Cells. Curr. Color. Cancer Rep. 2012, 8, 277–289. [Google Scholar] [CrossRef]
- Giraud, J.; Failla, L.M.; Pascussi, J.-M.; Lagerqvist, E.L.; Ollier, J.; Finetti, P.; Bertucci, F.; Ya, C.; Gasmi, I.; Bourgaux, J.-F.; et al. Autocrine Secretion of Progastrin Promotes the Survival and Self-Renewal of Colon Cancer Stem-like Cells. Cancer Res. 2016, 76, 3618–3628. [Google Scholar] [CrossRef]
- Wu, H.; Owlia, A.; Singh, P. Precursor Peptide Progastrin(1-80) Reduces Apoptosis of Intestinal Epithelial Cells and Upregulates Cytochrome c Oxidase Vb Levels and Synthesis of ATP. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G1097–G1110. [Google Scholar] [CrossRef]
- Najib, S.; Kowalski-Chauvel, A.; Do, C.; Roche, S.; Cohen-Jonathan-Moyal, E.; Seva, C. Progastrin a New Pro-Angiogenic Factor in Colorectal Cancer. Oncogene 2015, 34, 3120–3130. [Google Scholar] [CrossRef]
- Xiao, L.; Kovac, S.; Chang, M.; Shulkes, A.; Baldwin, G.S.; Patel, O. Induction of Gastrin Expression in Gastrointestinal Cells by Hypoxia or Cobalt Is Independent of Hypoxia-Inducible Factor (HIF). Endocrinology 2012, 153, 3006–3016. [Google Scholar] [CrossRef]
- Horst, B.; Pradhan, S.; Chaudhary, R.; Listik, E.; Quintero-Macias, L.; Choi, A.S.; Southard, M.; Liu, Y.; Whitaker, R.; Hempel, N.; et al. Hypoxia-Induced Inhibin Promotes Tumor Growth and Vascular Permeability in Ovarian Cancers. Commun. Biol. 2022, 5, 536. [Google Scholar] [CrossRef] [PubMed]
- Laval, M.; Baldwin, G.S.; Shulkes, A.; Marshall, K.M. Increased Gastrin Gene Expression Provides a Physiological Advantage to Mice under Hypoxic Conditions. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G76–G84. [Google Scholar] [CrossRef] [PubMed]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular Principles of Metastasis: A Hallmark of Cancer Revisited. Signal Transduct. Target. Ther. 2020, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, T.N. Editorial: Metabolism and Cell Adhesion in Cancer. Front. Cell Dev. Biol. 2022, 10, 871471. [Google Scholar] [CrossRef] [PubMed]
- Hollande, F.; Lee, D.J.; Choquet, A.; Roche, S.; Baldwin, G.S. Adherens Junctions and Tight Junctions Are Regulated via Different Pathways by Progastrin in Epithelial Cells. J. Cell Sci. 2003, 116, 1187–1197. [Google Scholar] [CrossRef] [PubMed]
- Fénié, N.; Bertrand, C.; Lacombe, A.; Roche, S.; Bousquet, C.; Gouazé-Andersson, V.; Toulas, C.; Moyal, E.; Ferrand, A. Abstract B16: Progastrin Activates Colon Fibroblasts and Participates to the Dialogue between Tumor Epithelial Cells and Stromal Fibroblasts in Colorectal Cancer. Cancer Res. 2015, 75, B16. [Google Scholar] [CrossRef]
- Duckworth, C.A.; Clyde, D.; Worthley, D.L.; Wang, T.C.; Varro, A.; Pritchard, D.M. Progastrin-Induced Secretion of Insulin-like Growth Factor 2 from Colonic Myofibroblasts Stimulates Colonic Epithelial Proliferation in Mice. Gastroenterology 2013, 145, 197–208.e3. [Google Scholar] [CrossRef] [PubMed]
- Stange, D.E.; Koo, B.-K.; Huch, M.; Sibbel, G.; Basak, O.; Lyubimova, A.; Kujala, P.; Bartfeld, S.; Koster, J.; Geahlen, J.H.; et al. Differentiated Troy+ Chief Cells Act as Reserve Stem Cells to Generate All Lineages of the Stomach Epithelium. Cell 2013, 155, 357–368. [Google Scholar] [CrossRef]
- Xiao, S.; Zhou, L. Gastric Stem Cells: Physiological and Pathological Perspectives. Front. Cell Dev. Biol. 2020, 8, 571536. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Jin, G.; Wang, H.; Chen, X.; Westphalen, C.B.; Asfaha, S.; Renz, B.W.; Ariyama, H.; Dubeykovskaya, Z.A.; Takemoto, Y.; et al. CCK2R Identifies and Regulates Gastric Antral Stem Cell States and Carcinogenesis. Gut 2015, 64, 544–553. [Google Scholar] [CrossRef]
- Berger, H.; Marques, M.S.; Zietlow, R.; Meyer, T.F.; Machado, J.C.; Figueiredo, C. Gastric Cancer Pathogenesis. Helicobacter 2016, 21 (Suppl. S1), 34–38. [Google Scholar] [CrossRef] [PubMed]
- Amjadi, O.; Alizadeh-Navaei, R.; Rezapour, M.; Omrani-Nava, V.; Moosazadeh, M.; Azadeh, H.; Zaboli, E.; Ahmadi, M.; Hedayatizadeh-Omran, A. Association between Serum Progastrin Biomarker Level and Gastric Cancer. Asian Pac. J. Cancer Prev. APJCP 2022, 23, 3595–3599. [Google Scholar] [CrossRef] [PubMed]
- Konturek, S.J.; Gonciarz, M.; Gonciarz, Z.; Bielanski, W.; Mazur, W.; Mularczyk, A.; Konturek, P.C.; Goetze, J.P.; Rehfeld, J.F. Progastrin and Its Products from Patients with Chronic Viral Hepatitis and Liver Cirrhosis. Scand. J. Gastroenterol. 2003, 38, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Caplin, M.; Khan, K.; Savage, K.; Rode, J.; Varro, A.; Michaeli, D.; Grimes, S.; Brett, B.; Pounder, R.; Dhillon, A. Expression and Processing of Gastrin in Hepatocellular Carcinoma, Fibrolamellar Carcinoma and Cholangiocarcinoma. J. Hepatol. 1999, 30, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Westwood, D.A.; Patel, O.; Christophi, C.; Shulkes, A.; Baldwin, G.S. Progastrin: A Potential Predictive Marker of Liver Metastasis in Colorectal Cancer. Int. J. Color. Dis. 2017, 32, 1061–1064. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.C.; Macrae, F.A.; Pizzey, C.; Baldwin, G.S.; Shulkes, A. Circulating Gastrin Concentrations in Patients at Increased Risk of Developing Colorectal Carcinoma. J. Gastroenterol. Hepatol. 2014, 29, 480–486. [Google Scholar] [CrossRef]
- Kowalski-Chauvel, A.; Gouaze-Andersson, V.; Vignolle-Vidoni, A.; Delmas, C.; Toulas, C.; Cohen-Jonathan-Moyal, E.; Seva, C. Targeting Progastrin Enhances Radiosensitization of Colorectal Cancer Cells. Oncotarget 2017, 8, 58587–58600. [Google Scholar] [CrossRef] [PubMed]
- You, B.; Kepenekian, V.; Prieur, A.; Caceres, M.; Payen-Gay, L.F.; Liaud, P.; Flaceliere, M.; Tod, M.; Villeneuve, L.; Bibeau, F.; et al. Progastrin, a New Blood Biomarker for the Diagnostic and Therapeutic Monitoring, in Gastro-Intestinal Cancers: A BIG-RENAPE Project. Ann. Oncol. 2018, 29, viii37. [Google Scholar] [CrossRef]
- You, B.; Couraud, S.; Ceruse, P.; Badet, L.; Paparel, P.; Walter, T.; Guillet, M.; Merle, P.; Philip, C.-A.; Ducray, F.; et al. Diagnostic Value of HPG80, as a New Multi-Cancer Blood Biomarker, in 16 Different Cancers: Results of the ONCOPRO Prospective Study. J. Clin. Oncol. 2023, 41, 3033. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fioretzaki, R.; Sarantis, P.; Charalampakis, N.; Christofidis, K.; Mylonakis, A.; Koustas, E.; Karamouzis, M.V.; Sakellariou, S.; Schizas, D. Progastrin: An Overview of Its Crucial Role in the Tumorigenesis of Gastrointestinal Cancers. Biomedicines 2024, 12, 885. https://doi.org/10.3390/biomedicines12040885
Fioretzaki R, Sarantis P, Charalampakis N, Christofidis K, Mylonakis A, Koustas E, Karamouzis MV, Sakellariou S, Schizas D. Progastrin: An Overview of Its Crucial Role in the Tumorigenesis of Gastrointestinal Cancers. Biomedicines. 2024; 12(4):885. https://doi.org/10.3390/biomedicines12040885
Chicago/Turabian StyleFioretzaki, Rodanthi, Panagiotis Sarantis, Nikolaos Charalampakis, Konstantinos Christofidis, Adam Mylonakis, Evangelos Koustas, Michalis V. Karamouzis, Stratigoula Sakellariou, and Dimitrios Schizas. 2024. "Progastrin: An Overview of Its Crucial Role in the Tumorigenesis of Gastrointestinal Cancers" Biomedicines 12, no. 4: 885. https://doi.org/10.3390/biomedicines12040885