
Mesenchymal Stem Cells and Purinergic Signaling in Autism Spectrum Disorder: Bridging the Gap between Cell-Based Strategies and Neuro-Immune Modulation


Abstract
1. Introduction
2. What Is Autism Spectrum Disorder?
3. Etiology of Autism Spectrum Disorder
3.1. Environmental Factors
3.2. Genetic Factors
3.3. Immune Dysregulation
3.3.1. Prenatal Inflammation
3.3.2. Postnatal Inflammation
4. How Does Purinergic Signaling Affect Neuroinflammation?
5. Functional Attributes of Mesenchymal Stem Cells Revisited
5.1. Immunomodulatory Properties of MSCs
5.2. Purinergic Signaling of MSCs

{kind=link}
{kind=link}
5.3. Release of Extracellular Vesicles
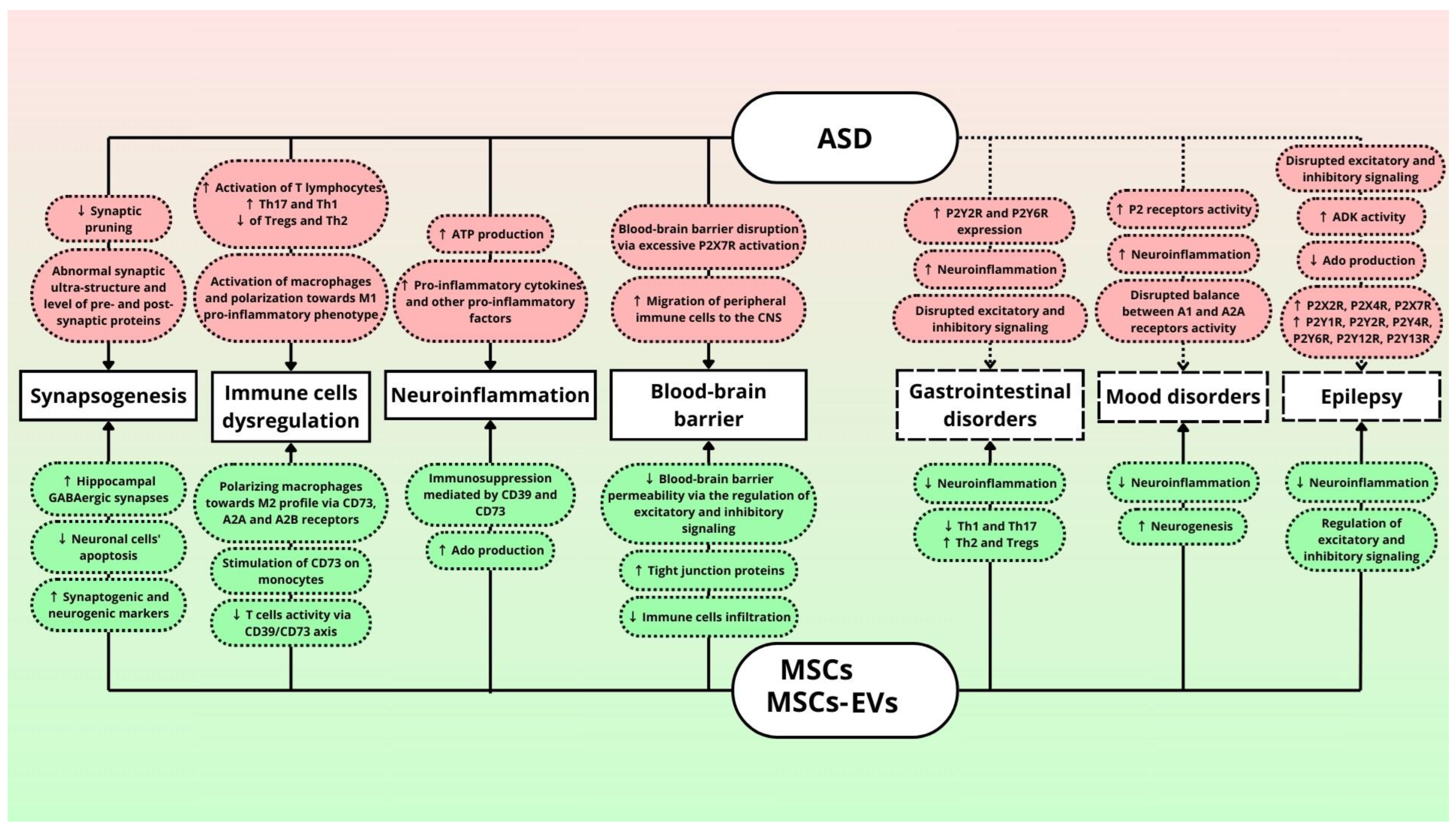
6. Role of Purinergic Signaling in ASD and Comorbidities
6.1. Purinergic Signaling Disturbances in ASD Etiology
6.2. Purinergic Signaling Disturbances in ASD-Related Comorbidities
6.2.1. Epilepsy
6.2.2. Mood Disorders
6.2.3. Gastrointestinal Disorders
7. Role of Mesenchymal Stem Cells in ASD Treatment
7.1. Purinergic Implications of MSCs in Neuroinflammation
7.2. MSCs Influence on Immune Cells
7.3. Purinergic Implications of MSCs in Blood–Brain Barrier Permeability
7.4. Current MSC Applications in ASD Treatment in Animal Models and Clinical Trials
8. Concluding Remarks and Future Perspectives
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maenner, M.J.; Warren, Z.; Williams, A.R.; Amoakohene, E.; Bakian, A.V.; Bilder, D.A.; Durkin, M.S.; Fitzgerald, R.T.; Furnier, S.M.; Hughes, M.M.; et al. Prevalence and Characteristics of Autism Spectrum Disorder Among Children Aged 8 Years—Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2020. MMWR Surveill. Summ. 2023, 72, 1–14. [Google Scholar] [CrossRef]
- Salari, N.; Rasoulpoor, S.; Rasoulpoor, S.; Shohaimi, S.; Jafarpour, S.; Abdoli, N.; Khaledi-Paveh, B.; Mohammadi, M. The Global Prevalence of Autism Spectrum Disorder: A Comprehensive Systematic Review and Meta-Analysis. Ital. J. Pediatr. 2022, 48, 112. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Xu, X.; Yan, W.; Zou, X.; Wu, L.; Luo, X.; Li, T.; Huang, Y.; Guan, H.; Chen, X.; et al. Prevalence of Autism Spectrum Disorder in China: A Nationwide Multi-Center Population-Based Study Among Children Aged 6 to 12 Years. Neurosci. Bull. 2020, 36, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Kularatna, S.; Jadambaa, A.; Senanayake, S.; Brain, D.; Hawker, N.; Kasparian, N.A.; Abell, B.; Auld, B.; Eagleson, K.; Justo, R.; et al. The Cost of Neurodevelopmental Disability: Scoping Review of Economic Evaluation Methods. Clin. Outcomes Res. 2022, 14, 665–682. [Google Scholar] [CrossRef]
- Edelson, S.M.; Nicholas, D.B.; Stoddart, K.P.; Bauman, M.B.; Mawlam, L.; Lawson, W.B.; Jose, C.; Morris, R.; Wright, S.D. Strategies for Research, Practice, and Policy for Autism in Later Life: A Report from a Think Tank on Aging and Autism. J. Autism Dev. Disord. 2021, 51, 382–390. [Google Scholar] [CrossRef]
- Dückert, S.; Gewohn, P.; König, H.; Schöttle, D.; Konnopka, A.; Rahlff, P.; Erik, F.; Vogeley, K.; Schulz, H.; David, N.; et al. Barriers and Needs in Mental Healthcare of Adults with Autism Spectrum Disorder in Germany: A Qualitative Study in Autistic Adults, Relatives, and Healthcare Providers. BMC Psychiatry 2023, 23, 528. [Google Scholar] [CrossRef] [PubMed]
- Andrzejewska, A.; Lukomska, B.; Janowski, M. Concise Review: Mesenchymal Stem Cells: From Roots to Boost. Stem Cells 2019, 37, 855–864. [Google Scholar] [CrossRef]
- Gomez-Salazar, M.; Gonzalez-Galofre, Z.N.; Casamitjana, J.; Crisan, M.; James, A.W.; Péault, B. Five Decades Later, Are Mesenchymal Stem Cells Still Relevant? Front. Bioeng. Biotechnol. 2020, 8, 148. [Google Scholar] [CrossRef]
- Search for: Mesenchymal Stem Cells|Card Results|ClinicalTrials.Gov. Available online: https://clinicaltrials.gov/search?intr=Mesenchymal%20Stem%20Cells (accessed on 24 April 2024).
- Arbuck, D. Could Stem Cells Have a Role in Treating Mental Illnesses? Curr. Psychiatry 2021, 20, 35–41. [Google Scholar] [CrossRef]
- Ballen, K.; Kurtzberg, J. Exploring New Therapies for Children with Autism: “Do No Harm” Does Not Mean Do Not Try. Stem Cells Transl. Med. 2021, 10, 823–825. [Google Scholar] [CrossRef]
- Burnstock, G. Purine and Purinergic Receptors. Brain Neurosci. Adv. 2018, 2, 2398212818817494. [Google Scholar] [CrossRef]
- Yegutkin, G.G. Enzymes Involved in Metabolism of Extracellular Nucleotides and Nucleosides: Functional Implications and Measurement of Activities. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 473–497. [Google Scholar] [CrossRef] [PubMed]
- Reichert, K.P.; Castro, M.F.V.; Assmann, C.E.; Bottari, N.B.; Miron, V.V.; Cardoso, A.; Stefanello, N.; Morsch, V.M.M.; Schetinger, M.R.C. Diabetes and Hypertension: Pivotal Involvement of Purinergic Signaling. Biomed. Pharmacother. 2021, 137, 111273. [Google Scholar] [CrossRef] [PubMed]
- Zarrinmayeh, H.; Territo, P.R. Purinergic Receptors of the Central Nervous System: Biology, PET Ligands, and Their Applications. Mol. Imaging 2020, 19, 1536012120927609. [Google Scholar] [CrossRef]
- North, R.A. P2X Receptors. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150427. [Google Scholar] [CrossRef] [PubMed]
- Savio, L.E.B.; de Andrade Mello, P.; da Silva, C.G.; Coutinho-Silva, R. The P2X7 Receptor in Inflammatory Diseases: Angel or Demon? Front. Pharmacol. 2018, 9, 52. [Google Scholar] [CrossRef]
- Zimmermann, H. History of Ectonucleotidases and Their Role in Purinergic Signaling. Biochem. Pharmacol. 2021, 187, 114322. [Google Scholar] [CrossRef]
- Sanchack, K.E.; Thomas, C.A. Autism Spectrum Disorder: Primary Care Principles. Am. Fam. Physician 2016, 94, 972–979. [Google Scholar]
- Diagnostic and Statistical Manual of Mental Disorders: DSM-5, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013; ISBN 978-0-89042-554-1.
- Tsang, L.P.M.; How, C.H.; Yeleswarapu, S.P.; Wong, C.M. Autism Spectrum Disorder: Early Identification and Management in Primary Care. Singap. Med. J. 2019, 60, 324–328. [Google Scholar] [CrossRef]
- Wong, R.S.Y. Neuroinflammation in autism spectrum disorders: Potential target for mesenchymal stem cell-based therapy. Egypt. J. Neurol. Psychiatry Neurosurg. 2022, 58, 91. [Google Scholar] [CrossRef]
- Ng, M.; de Montigny, J.G.; Ofner, M.; Do, M.T. Environmental Factors Associated with Autism Spectrum Disorder: A Scoping Review for the Years 2003-2013. Health Promot. Chronic. Dis. Prev. Can. 2017, 37, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Eskenazi, B.; Widjaja, F.; Cordero, J.F.; Hendren, R.L. Improving Autism Perinatal Risk Factors: A Systematic Review. Med. Hypotheses 2019, 127, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Lyall, K.; Schmidt, R.J.; Hertz-Picciotto, I. Maternal Lifestyle and Environmental Risk Factors for Autism Spectrum Disorders. Int. J. Epidemiol. 2014, 43, 443–464. [Google Scholar] [CrossRef] [PubMed]
- Bölte, S.; Girdler, S.; Marschik, P.B. The Contribution of Environmental Exposure to the Etiology of Autism Spectrum Disorder. Cell. Mol. Life Sci. 2019, 76, 1275–1297. [Google Scholar] [CrossRef] [PubMed]
- Xiang, A.H.; Wang, X.; Martinez, M.P.; Page, K.; Buchanan, T.A.; Feldman, R.K. Maternal Type 1 Diabetes and Risk of Autism in Offspring. JAMA 2018, 320, 89–91. [Google Scholar] [CrossRef] [PubMed]
- Styles, M.; Alsharshani, D.; Samara, M.; Alsharshani, M.; Khattab, A.; Qoronfleh, M.W.; Al-Dewik, N.I. Risk Factors, Diagnosis, Prognosis and Treatment of Autism. Front. Biosci. (Landmark Ed.) 2020, 25, 1682–1717. [Google Scholar] [CrossRef] [PubMed]
- Lyall, K.; Croen, L.; Daniels, J.; Fallin, M.D.; Ladd-Acosta, C.; Lee, B.K.; Park, B.Y.; Snyder, N.W.; Schendel, D.; Volk, H.; et al. The Changing Epidemiology of Autism Spectrum Disorders. Annu. Rev. Public Health 2017, 38, 81–102. [Google Scholar] [CrossRef] [PubMed]
- Kapra, O.; Rotem, R.; Gross, R. The Association Between Prenatal Exposure to Antidepressants and Autism: Some Research and Public Health Aspects. Front. Psychiatry 2020, 11, 555740. [Google Scholar] [CrossRef] [PubMed]
- Modabbernia, A.; Velthorst, E.; Reichenberg, A. Environmental Risk Factors for Autism: An Evidence-Based Review of Systematic Reviews and Meta-Analyses. Mol. Autism 2017, 8, 13. [Google Scholar] [CrossRef]
- Chaste, P.; Leboyer, M. Autism Risk Factors: Genes, Environment, and Gene-Environment Interactions. Dialogues Clin. Neurosci. 2012, 14, 281–292. [Google Scholar] [CrossRef]
- Ahearn, W.H. What Every Behavior Analyst Should Know About the “MMR Causes Autism” Hypothesis. Behav. Anal. Pract. 2010, 3, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Masini, E.; Loi, E.; Vega-Benedetti, A.F.; Carta, M.; Doneddu, G.; Fadda, R.; Zavattari, P. An Overview of the Main Genetic, Epigenetic and Environmental Factors Involved in Autism Spectrum Disorder Focusing on Synaptic Activity. Int. J. Mol. Sci. 2020, 21, 8290. [Google Scholar] [CrossRef] [PubMed]
- Wiśniowiecka-Kowalnik, B.; Nowakowska, B.A. Genetics and Epigenetics of Autism Spectrum Disorder—Current Evidence in the Field. J. Appl. Genet. 2019, 60, 37–47. [Google Scholar] [CrossRef] [PubMed]
- DeThorne, L.S.; Ceman, S. Genetic Testing and Autism: Tutorial for Communication Sciences and Disorders. J. Commun. Disord. 2018, 74, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Woodbury-Smith, M.; Scherer, S.W. Progress in the Genetics of Autism Spectrum Disorder. Dev. Med. Child Neurol. 2018, 60, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Chahrour, M.; Kleiman, R.J.; Manzini, M.C. Translating Genetic and Preclinical Findings into Autism Therapies. Dialogues Clin. Neurosci. 2017, 19, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Cheroni, C.; Caporale, N.; Testa, G. Autism Spectrum Disorder at the Crossroad between Genes and Environment: Contributions, Convergences, and Interactions in ASD Developmental Pathophysiology. Mol. Autism 2020, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Havdahl, A.; Niarchou, M.; Starnawska, A.; Uddin, M.; van der Merwe, C.; Warrier, V. Genetic Contributions to Autism Spectrum Disorder. Psychol. Med. 2021, 51, 2260–2273. [Google Scholar] [CrossRef]
- Kereszturi, É. Diversity and Classification of Genetic Variations in Autism Spectrum Disorder. Int. J. Mol. Sci. 2023, 24, 16768. [Google Scholar] [CrossRef]
- Solek, C.M.; Farooqi, N.; Verly, M.; Lim, T.K.; Ruthazer, E.S. Maternal Immune Activation in Neurodevelopmental Disorders. Dev. Dyn. 2018, 247, 588–619. [Google Scholar] [CrossRef]
- Zawadzka, A.; Cieślik, M.; Adamczyk, A. The Role of Maternal Immune Activation in the Pathogenesis of Autism: A Review of the Evidence, Proposed Mechanisms and Implications for Treatment. Int. J. Mol. Sci. 2021, 22, 11516. [Google Scholar] [CrossRef]
- Lampiasi, N.; Bonaventura, R.; Deidda, I.; Zito, F.; Russo, R. Inflammation and the Potential Implication of Macrophage-Microglia Polarization in Human ASD: An Overview. Int. J. Mol. Sci. 2023, 24, 2703. [Google Scholar] [CrossRef]
- Usui, N.; Kobayashi, H.; Shimada, S. Neuroinflammation and Oxidative Stress in the Pathogenesis of Autism Spectrum Disorder. Int. J. Mol. Sci. 2023, 24, 5487. [Google Scholar] [CrossRef] [PubMed]
- McLellan, J.; Kim, D.H.J.; Bruce, M.; Ramirez-Celis, A.; Van de Water, J. Maternal Immune Dysregulation and Autism-Understanding the Role of Cytokines, Chemokines and Autoantibodies. Front. Psychiatry 2022, 13, 834910. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.-K.; Choi, G.B.; Huh, J.R. Maternal Inflammation and Its Ramifications on Fetal Neurodevelopment. Trends Immunol. 2022, 43, 230–244. [Google Scholar] [CrossRef]
- Beversdorf, D.Q.; Stevens, H.E.; Jones, K.L. Prenatal Stress, Maternal Immune Dysregulation, and Their Association With Autism Spectrum Disorders. Curr. Psychiatry Rep. 2018, 20, 76. [Google Scholar] [CrossRef]
- Sotgiu, S.; Manca, S.; Gagliano, A.; Minutolo, A.; Melis, M.C.; Pisuttu, G.; Scoppola, C.; Bolognesi, E.; Clerici, M.; Guerini, F.R.; et al. Immune Regulation of Neurodevelopment at the Mother-Foetus Interface: The Case of Autism. Clin. Transl. Immunol. 2020, 9, e1211. [Google Scholar] [CrossRef]
- Siniscalco, D.; Schultz, S.; Brigida, A.L.; Antonucci, N. Inflammation and Neuro-Immune Dysregulations in Autism Spectrum Disorders. Pharmaceuticals 2018, 11, 56. [Google Scholar] [CrossRef] [PubMed]
- Heidari, A.; Rostam-Abadi, Y.; Rezaei, N. The Immune System and Autism Spectrum Disorder: Association and Therapeutic Challenges. Acta Neurobiol. Exp. (Wars) 2021, 81, 249–263. [Google Scholar] [CrossRef]
- Gładysz, D.; Krzywdzińska, A.; Hozyasz, K.K. Immune Abnormalities in Autism Spectrum Disorder—Could They Hold Promise for Causative Treatment? Mol. Neurobiol. 2018, 55, 6387–6435. [Google Scholar] [CrossRef]
- Erbescu, A.; Papuc, S.M.; Budisteanu, M.; Arghir, A.; Neagu, M. Re-Emerging Concepts of Immune Dysregulation in Autism Spectrum Disorders. Front. Psychiatry 2022, 13, 1006612. [Google Scholar] [CrossRef]
- Masi, A.; Glozier, N.; Dale, R.; Guastella, A.J. The Immune System, Cytokines, and Biomarkers in Autism Spectrum Disorder. Neurosci. Bull. 2017, 33, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Hughes, H.K.; Moreno, R.J.; Ashwood, P. Innate Immune Dysfunction and Neuroinflammation in Autism Spectrum Disorder (ASD). Brain Behav. Immun. 2023, 108, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Hughes, H.K.; Mills Ko, E.; Rose, D.; Ashwood, P. Immune Dysfunction and Autoimmunity as Pathological Mechanisms in Autism Spectrum Disorders. Front. Cell. Neurosci. 2018, 12, 405. [Google Scholar] [CrossRef]
- Ellul, P.; Rosenzwajg, M.; Peyre, H.; Fourcade, G.; Mariotti-Ferrandiz, E.; Trebossen, V.; Klatzmann, D.; Delorme, R. Regulatory T Lymphocytes/Th17 Lymphocytes Imbalance in Autism Spectrum Disorders: Evidence from a Meta-Analysis. Mol. Autism 2021, 12, 68. [Google Scholar] [CrossRef] [PubMed]
- Moaaz, M.; Youssry, S.; Elfatatry, A.; El Rahman, M.A. Th17/Treg Cells Imbalance and Their Related Cytokines (IL-17, IL-10 and TGF-β) in Children with Autism Spectrum Disorder. J. Neuroimmunol. 2019, 337, 577071. [Google Scholar] [CrossRef]
- Meltzer, A.; Van de Water, J. The Role of the Immune System in Autism Spectrum Disorder. Neuropsychopharmacology 2017, 42, 284–298. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.F.; Zoheir, K.M.A.; Ansari, M.A.; Nadeem, A.; Bakheet, S.A.; Al-Ayadhi, L.Y.; Alzahrani, M.Z.; Al-Shabanah, O.A.; Al-Harbi, M.M.; Attia, S.M. Dysregulation of Th1, Th2, Th17, and T Regulatory Cell-Related Transcription Factor Signaling in Children with Autism. Mol. Neurobiol. 2017, 54, 4390–4400. [Google Scholar] [CrossRef]
- Akintunde, M.E.; Rose, M.; Krakowiak, P.; Heuer, L.; Ashwood, P.; Hansen, R.; Hertz-Picciotto, I.; Van de Water, J. Increased Production of IL-17 in Children with Autism Spectrum Disorders and Co-Morbid Asthma. J. Neuroimmunol. 2015, 286, 33–41. [Google Scholar] [CrossRef]
- Hu, C.; Li, H.; Li, J.; Luo, X.; Hao, Y. Microglia: Synaptic Modulator in Autism Spectrum Disorder. Front. Psychiatry 2022, 13, 958661. [Google Scholar] [CrossRef]
- Xiong, Y.; Chen, J.; Li, Y. Microglia and Astrocytes Underlie Neuroinflammation and Synaptic Susceptibility in Autism Spectrum Disorder. Front. Neurosci. 2023, 17, 1125428. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, F.; Quintana, F.J. The Role of Astrocytes in CNS Inflammation. Trends Immunol. 2020, 41, 805–819. [Google Scholar] [CrossRef] [PubMed]
- Linnerbauer, M.; Wheeler, M.A.; Quintana, F.J. Astrocyte Crosstalk in CNS Inflammation. Neuron 2020, 108, 608–622. [Google Scholar] [CrossRef] [PubMed]
- Cekanaviciute, E.; Buckwalter, M.S. Astrocytes: Integrative Regulators of Neuroinflammation in Stroke and Other Neurological Diseases. Neurotherapeutics 2016, 13, 685–701. [Google Scholar] [CrossRef] [PubMed]
- Jo, M.; Kim, J.-H.; Song, G.J.; Seo, M.; Hwang, E.M.; Suk, K. Astrocytic Orosomucoid-2 Modulates Microglial Activation and Neuroinflammation. J. Neurosci. 2017, 37, 2878–2894. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.-L.; Cai, Y.-Q.; Zhu, M.-C.; Xing, L.-J.; Wang, X. The Yin and Yang Functions of Extracellular ATP and Adenosine in Tumor Immunity. Cancer Cell Int. 2020, 20, 110. [Google Scholar] [CrossRef] [PubMed]
- Di Virgilio, F.; Vultaggio-Poma, V.; Falzoni, S.; Giuliani, A.L. Extracellular ATP: A Powerful Inflammatory Mediator in the Central Nervous System. Neuropharmacology 2023, 224, 109333. [Google Scholar] [CrossRef] [PubMed]
- Pasquini, S.; Contri, C.; Borea, P.A.; Vincenzi, F.; Varani, K. Adenosine and Inflammation: Here, There and Everywhere. Int. J. Mol. Sci. 2021, 22, 7685. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Sarti, A.C.; Coutinho-Silva, R. Purinergic Signaling, DAMPs, and Inflammation. Am. J. Physiol. Cell Physiol. 2020, 318, C832–C835. [Google Scholar] [CrossRef]
- Calzaferri, F.; Ruiz-Ruiz, C.; de Diego, A.M.G.; de Pascual, R.; Méndez-López, I.; Cano-Abad, M.F.; Maneu, V.; de Los Ríos, C.; Gandía, L.; García, A.G. The Purinergic P2X7 Receptor as a Potential Drug Target to Combat Neuroinflammation in Neurodegenerative Diseases. Med. Res. Rev. 2020, 40, 2427–2465. [Google Scholar] [CrossRef]
- Territo, P.R.; Zarrinmayeh, H. P2X7 Receptors in Neurodegeneration: Potential Therapeutic Applications From Basic to Clinical Approaches. Front. Cell. Neurosci. 2021, 15, 617036. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; You, H.; Hu, X.; Luo, Y.; Zhang, Z.; Song, Y.; An, J.; Lu, H. Microglia-Astrocyte Interaction in Neural Development and Neural Pathogenesis. Cells 2023, 12, 1942. [Google Scholar] [CrossRef]
- Matejuk, A.; Ransohoff, R.M. Crosstalk Between Astrocytes and Microglia: An Overview. Front. Immunol. 2020, 11, 1416. [Google Scholar] [CrossRef]
- Babiec, L.; Wilkaniec, A.; Matuszewska, M.; Pałasz, E.; Cieślik, M.; Adamczyk, A. Alterations of Purinergic Receptors Levels and Their Involvement in the Glial Cell Morphology in a Pre-Clinical Model of Autism Spectrum Disorders. Brain Sci. 2023, 13, 1088. [Google Scholar] [CrossRef]
- Antonioli, L.; Fornai, M.; Blandizzi, C.; Pacher, P.; Haskó, G. Adenosine Signaling and the Immune System: When a Lot Could Be Too Much. Immunol. Lett. 2019, 205, 9–15. [Google Scholar] [CrossRef]
- Roszek, K.; Wujak, M. How to Influence the Mesenchymal Stem Cells Fate? Emerging Role of Ectoenzymes Metabolizing Nucleotides. J. Cell. Physiol. 2018, 234, 320–334. [Google Scholar] [CrossRef]
- Jiang, W.; Xu, J. Immune Modulation by Mesenchymal Stem Cells. Cell Prolif. 2020, 53, e12712. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Scholtemeijer, M.; Shah, K. Mesenchymal Stem Cell Immunomodulation: Mechanisms and Therapeutic Potential. Trends Pharmacol. Sci. 2020, 41, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Perez, E.; Gonzalez-Pujana, A.; Igartua, M.; Santos-Vizcaino, E.; Hernandez, R.M. Mesenchymal Stromal Cell Secretome for the Treatment of Immune-Mediated Inflammatory Diseases: Latest Trends in Isolation, Content Optimization and Delivery Avenues. Pharmaceutics 2021, 13, 1802. [Google Scholar] [CrossRef]
- Liu, J.; Gao, J.; Liang, Z.; Gao, C.; Niu, Q.; Wu, F.; Zhang, L. Mesenchymal Stem Cells and Their Microenvironment. Stem Cell Res. Ther. 2022, 13, 429. [Google Scholar] [CrossRef]
- Villa, C.; Combi, R.; Conconi, D.; Lavitrano, M. Patient-Derived Induced Pluripotent Stem Cells (iPSCs) and Cerebral Organoids for Drug Screening and Development in Autism Spectrum Disorder: Opportunities and Challenges. Pharmaceutics 2021, 13, 280. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Huang, W.; Liu, J.; Tian, J.; Wang, S.; Rui, K. Effects of Mesenchymal Stem Cell-Derived Exosomes on Autoimmune Diseases. Front. Immunol. 2021, 12, 749192. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Jiang, J.; Gu, Z.; Zhang, J.; Chen, Y.; Liu, X. Mesenchymal Stromal Cell Therapies: Immunomodulatory Properties and Clinical Progress. Stem Cell Res. Ther. 2020, 11, 345. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Guan, Y.; Li, C.; Zhang, T.; Meng, F.; Zhang, J.; Li, J.; Chen, S.; Wang, Q.; Wang, Y.; et al. Immunomodulatory Effects of Mesenchymal Stem Cells in Peripheral Nerve Injury. Stem Cell Res. Ther. 2022, 13, 18. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Yang, J.; Fang, J.; Zhou, Y.; Candi, E.; Wang, J.; Hua, D.; Shao, C.; Shi, Y. The Secretion Profile of Mesenchymal Stem Cells and Potential Applications in Treating Human Diseases. Signal Transduct. Target. Ther. 2022, 7, 92. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wu, Q.; Tam, P.K.H. Immunomodulatory Mechanisms of Mesenchymal Stem Cells and Their Potential Clinical Applications. Int. J. Mol. Sci. 2022, 23, 10023. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Fan, X.; Liu, Y.; Jie, P.; Mazhar, M.; Liu, Y.; Dechsupa, N.; Wang, L. Immunomodulatory Mechanisms and Therapeutic Potential of Mesenchymal Stem Cells. Stem Cell Rev. Rep. 2023, 19, 1214–1231. [Google Scholar] [CrossRef]
- Coppi, E.; Pugliese, A.M.; Urbani, S.; Melani, A.; Cerbai, E.; Mazzanti, B.; Bosi, A.; Saccardi, R.; Pedata, F. ATP Modulates Cell Proliferation and Elicits Two Different Electrophysiological Responses in Human Mesenchymal Stem Cells. Stem Cells 2007, 25, 1840–1849. [Google Scholar] [CrossRef]
- Katebi, M.; Soleimani, M.; Cronstein, B.N. Adenosine A2A Receptors Play an Active Role in Mouse Bone Marrow-Derived Mesenchymal Stem Cell Development. J. Leukoc. Biol. 2009, 85, 438–444. [Google Scholar] [CrossRef]
- Ferrari, D.; Gulinelli, S.; Salvestrini, V.; Lucchetti, G.; Zini, R.; Manfredini, R.; Caione, L.; Piacibello, W.; Ciciarello, M.; Rossi, L.; et al. Purinergic Stimulation of Human Mesenchymal Stem Cells Potentiates Their Chemotactic Response to CXCL12 and Increases the Homing Capacity and Production of Proinflammatory Cytokines. Exp. Hematol. 2011, 39, 360–374.e5. [Google Scholar] [CrossRef]
- Iser, I.C.; Bracco, P.A.; Gonçalves, C.E.I.; Zanin, R.F.; Nardi, N.B.; Lenz, G.; Battastini, A.M.O.; Wink, M.R. Mesenchymal Stem Cells from Different Murine Tissues Have Differential Capacity to Metabolize Extracellular Nucleotides. J. Cell. Biochem. 2014, 115, 1673–1682. [Google Scholar] [CrossRef]
- Glaser, T.; Cappellari, A.R.; Pillat, M.M.; Iser, I.C.; Wink, M.R.; Battastini, A.M.O.; Ulrich, H. Perspectives of Purinergic Signaling in Stem Cell Differentiation and Tissue Regeneration. Purinergic Signal. 2012, 8, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Kaebisch, C.; Schipper, D.; Babczyk, P.; Tobiasch, E. The Role of Purinergic Receptors in Stem Cell Differentiation. Comput. Struct. Biotechnol. J. 2015, 13, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Roszek, K.; Błaszczak, A.; Wujak, M.; Komoszyński, M. Nucleotides Metabolizing Ectoenzymes as Possible Markers of Mesenchymal Stem Cell Osteogenic Differentiation. Biochem. Cell Biol. 2013, 91, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Roszek, K.; Porowińska, D.; Bajek, A.; Hołysz, M.; Czarnecka, J. Chondrogenic Differentiation of Human Mesenchymal Stem Cells Results in Substantial Changes of Ecto-Nucleotides Metabolism. J. Cell. Biochem. 2015, 116, 2915–2923. [Google Scholar] [CrossRef]
- Wang, L.; Roger, S.; Yang, X.B.; Jiang, L.-H. Role of the Store-Operated Ca2+ Channel in ATP-Induced Ca2+ Signalling in Mesenchymal Stem Cells and Regulation of Cell Functions. Front. Biosci. (Landmark Ed.) 2021, 26, 1737–1745. [Google Scholar] [CrossRef]
- Galgaro, B.C.; Beckenkamp, L.R.; van den M Nunnenkamp, M.; Korb, V.G.; Naasani, L.I.S.; Roszek, K.; Wink, M.R. The Adenosinergic Pathway in Mesenchymal Stem Cell Fate and Functions. Med. Res. Rev. 2021, 41, 2316–2349. [Google Scholar] [CrossRef]
- Riddle, R.C.; Taylor, A.F.; Rogers, J.R.; Donahue, H.J. ATP Release Mediates Fluid Flow-Induced Proliferation of Human Bone Marrow Stromal Cells. J. Bone Miner. Res. 2007, 22, 589–600. [Google Scholar] [CrossRef]
- Czarnecka, J.; Porowińska, D.; Bajek, A.; Hołysz, M.; Roszek, K. Neurogenic Differentiation of Mesenchymal Stem Cells Induces Alterations in Extracellular Nucleotides Metabolism. J. Cell. Biochem. 2017, 118, 478–486. [Google Scholar] [CrossRef]
- Peng, H.; Hao, Y.; Mousawi, F.; Roger, S.; Li, J.; Sim, J.A.; Ponnambalam, S.; Yang, X.; Jiang, L.-H. Purinergic and Store-Operated Ca2+ Signaling Mechanisms in Mesenchymal Stem Cells and Their Roles in ATP-Induced Stimulation of Cell Migration. Stem Cells 2016, 34, 2102–2114. [Google Scholar] [CrossRef]
- Leung, Y.-M. P2X7 Receptor as a Double-Edged Sword: Neurotrophic and Neurotoxic Effects. BioMedicine 2011, 1, 16–20. [Google Scholar] [CrossRef]
- Miras-Portugal, M.T.; Gomez-Villafuertes, R.; Gualix, J.; Diaz-Hernandez, J.I.; Artalejo, A.R.; Ortega, F.; Delicado, E.G.; Perez-Sen, R. Nucleotides in Neuroregeneration and Neuroprotection. Neuropharmacology 2016, 104, 243–254. [Google Scholar] [CrossRef]
- Sun, D.; Junger, W.G.; Yuan, C.; Zhang, W.; Bao, Y.; Qin, D.; Wang, C.; Tan, L.; Qi, B.; Zhu, D.; et al. Shockwaves Induce Osteogenic Differentiation of Human Mesenchymal Stem Cells through ATP Release and Activation of P2X7 Receptors. Stem Cells 2013, 31, 1170–1180. [Google Scholar] [CrossRef] [PubMed]
- Orriss, I.R.; Key, M.L.; Hajjawi, M.O.R.; Arnett, T.R. Extracellular ATP Released by Osteoblasts Is a Key Local Inhibitor of Bone Mineralisation. PLoS ONE 2013, 8, e69057. [Google Scholar] [CrossRef]
- Zippel, N.; Limbach, C.A.; Ratajski, N.; Urban, C.; Luparello, C.; Pansky, A.; Kassack, M.U.; Tobiasch, E. Purinergic Receptors Influence the Differentiation of Human Mesenchymal Stem Cells. Stem Cells Dev. 2012, 21, 884–900. [Google Scholar] [CrossRef]
- Burnstock, G.; Arnett, T.R.; Orriss, I.R. Purinergic Signalling in the Musculoskeletal System. Purinergic Signal. 2013, 9, 541–572. [Google Scholar] [CrossRef]
- Carluccio, M.; Ziberi, S.; Zuccarini, M.; Giuliani, P.; Caciagli, F.; Di Iorio, P.; Ciccarelli, R. Adult Mesenchymal Stem Cells: Is There a Role for Purine Receptors in Their Osteogenic Differentiation? Purinergic Signal. 2020, 16, 263–287. [Google Scholar] [CrossRef]
- Corciulo, C.; Lendhey, M.; Wilder, T.; Schoen, H.; Cornelissen, A.S.; Chang, G.; Kennedy, O.D.; Cronstein, B.N. Endogenous Adenosine Maintains Cartilage Homeostasis and Exogenous Adenosine Inhibits Osteoarthritis Progression. Nat. Commun. 2017, 8, 15019. [Google Scholar] [CrossRef] [PubMed]
- Gharibi, B.; Abraham, A.A.; Ham, J.; Evans, B.A.J. Adenosine Receptor Subtype Expression and Activation Influence the Differentiation of Mesenchymal Stem Cells to Osteoblasts and Adipocytes. J. Bone Miner. Res. 2011, 26, 2112–2124. [Google Scholar] [CrossRef]
- Tan, L.; Zhao, B.; Ge, F.-T.; Sun, D.-H.; Yu, T. Shockwaves Inhibit Chondrogenic Differentiation of Human Mesenchymal Stem Cells in Association with Adenosine and A2B Receptors. Sci. Rep. 2017, 7, 14377. [Google Scholar] [CrossRef]
- Scarfì, S. Purinergic Receptors and Nucleotide Processing Ectoenzymes: Their Roles in Regulating Mesenchymal Stem Cell Functions. World J. Stem Cells 2014, 6, 153–162. [Google Scholar] [CrossRef]
- Bazzoni, R.; Takam Kamga, P.; Tanasi, I.; Krampera, M. Extracellular Vesicle-Dependent Communication Between Mesenchymal Stromal Cells and Immune Effector Cells. Front. Cell Dev. Biol. 2020, 8, 596079. [Google Scholar] [CrossRef]
- Kou, M.; Huang, L.; Yang, J.; Chiang, Z.; Chen, S.; Liu, J.; Guo, L.; Zhang, X.; Zhou, X.; Xu, X.; et al. Mesenchymal Stem Cell-Derived Extracellular Vesicles for Immunomodulation and Regeneration: A next Generation Therapeutic Tool? Cell Death Dis. 2022, 13, 580. [Google Scholar] [CrossRef]
- Ivosevic, Z.; Ljujic, B.; Pavlovic, D.; Matovic, V.; Gazdic Jankovic, M. Mesenchymal Stem Cell-Derived Extracellular Vesicles: New Soldiers in the War on Immune-Mediated Diseases. Cell Transplant. 2023, 32, 9636897231207194. [Google Scholar] [CrossRef]
- Adinolfi, E.; De Marchi, E.; Grignolo, M.; Szymczak, B.; Pegoraro, A. The P2X7 Receptor in Oncogenesis and Metastatic Dissemination: New Insights on Vesicular Release and Adenosinergic Crosstalk. Int. J. Mol. Sci. 2023, 24, 13906. [Google Scholar] [CrossRef]
- Winzer, R.; Nguyen, D.H.; Schoppmeier, F.; Cortesi, F.; Gagliani, N.; Tolosa, E. Purinergic Enzymes on Extracellular Vesicles: Immune Modulation on the Go. Front. Immunol. 2024, 15, 1362996. [Google Scholar] [CrossRef]
- Lombardi, M.; Gabrielli, M.; Adinolfi, E.; Verderio, C. Role of ATP in Extracellular Vesicle Biogenesis and Dynamics. Front. Pharmacol. 2021, 12, 654023. [Google Scholar] [CrossRef]
- Golia, M.T.; Gabrielli, M.; Verderio, C. P2X7 Receptor and Extracellular Vesicle Release. Int. J. Mol. Sci. 2023, 24, 9805. [Google Scholar] [CrossRef]
- Pegoraro, A.; De Marchi, E.; Ferracin, M.; Orioli, E.; Zanoni, M.; Bassi, C.; Tesei, A.; Capece, M.; Dika, E.; Negrini, M.; et al. P2X7 Promotes Metastatic Spreading and Triggers Release of miRNA-Containing Exosomes and Microvesicles from Melanoma Cells. Cell Death Dis. 2021, 12, 1088. [Google Scholar] [CrossRef]
- Caplan, H.; Olson, S.D.; Kumar, A.; George, M.; Prabhakara, K.S.; Wenzel, P.; Bedi, S.; Toledano-Furman, N.E.; Triolo, F.; Kamhieh-Milz, J.; et al. Mesenchymal Stromal Cell Therapeutic Delivery: Translational Challenges to Clinical Application. Front. Immunol. 2019, 10, 1645. [Google Scholar] [CrossRef]
- Turano, E.; Scambi, I.; Virla, F.; Bonetti, B.; Mariotti, R. Extracellular Vesicles from Mesenchymal Stem Cells: Towards Novel Therapeutic Strategies for Neurodegenerative Diseases. Int. J. Mol. Sci. 2023, 24, 2917. [Google Scholar] [CrossRef]
- Harrell, C.R.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Mesenchymal Stem Cell-Derived Exosomes and Other Extracellular Vesicles as New Remedies in the Therapy of Inflammatory Diseases. Cells 2019, 8, 1605. [Google Scholar] [CrossRef]
- Alessio, N.; Brigida, A.L.; Peluso, G.; Antonucci, N.; Galderisi, U.; Siniscalco, D. Stem Cell-Derived Exosomes in Autism Spectrum Disorder. Int. J. Environ. Res. Public Health 2020, 17, 944. [Google Scholar] [CrossRef]
- Dai, S.; Lin, J.; Hou, Y.; Luo, X.; Shen, Y.; Ou, J. Purine Signaling Pathway Dysfunction in Autism Spectrum Disorders: Evidence from Multiple Omics Data. Front. Mol. Neurosci. 2023, 16, 1089871. [Google Scholar] [CrossRef]
- Boccazzi, M.; Raffaele, S.; Zanettin, T.; Abbracchio, M.P.; Fumagalli, M. Altered Purinergic Signaling in Neurodevelopmental Disorders: Focus on P2 Receptors. Biomolecules 2023, 13, 856. [Google Scholar] [CrossRef]
- Babiec, L.; Wilkaniec, A.; Adamczyk, A. Prenatal Exposure to Valproic Acid Induces Alterations in the Expression and Activity of Purinergic Receptors in the Embryonic Rat Brain. Folia Neuropathol. 2022, 60, 390–402. [Google Scholar] [CrossRef]
- Meng, Q.; Zhang, W.; Wang, X.; Jiao, C.; Xu, S.; Liu, C.; Tang, B.; Chen, C. Human Forebrain Organoids Reveal Connections between Valproic Acid Exposure and Autism Risk. Transl. Psychiatry 2022, 12, 130. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, Q.; Yan, T.; Zhang, Y.; Xu, H.-J.; Yu, H.-P.; Tu, Z.; Guo, X.; Jiang, Y.-H.; Li, X.-J.; et al. Maternal Valproic Acid Exposure Leads to Neurogenesis Defects and Autism-like Behaviors in Non-Human Primates. Transl. Psychiatry 2019, 9, 267. [Google Scholar] [CrossRef]
- Naviaux, R.K.; Curtis, B.; Li, K.; Naviaux, J.C.; Bright, A.T.; Reiner, G.E.; Westerfield, M.; Goh, S.; Alaynick, W.A.; Wang, L.; et al. Low-Dose Suramin in Autism Spectrum Disorder: A Small, Phase I/II, Randomized Clinical Trial. Ann. Clin. Transl. Neurol. 2017, 4, 491–505. [Google Scholar] [CrossRef]
- Rodrigues, R.J.; Marques, J.M.; Cunha, R.A. Purinergic Signalling and Brain Development. Semin. Cell Dev. Biol. 2019, 95, 34–41. [Google Scholar] [CrossRef]
- Lewis, M.H.; Rajpal, H.; Muehlmann, A.M. Reduction of Repetitive Behavior by Co-Administration of Adenosine Receptor Agonists in C58 Mice. Pharmacol. Biochem. Behav. 2019, 181, 110–116. [Google Scholar] [CrossRef]
- Pasquini, S.; Contri, C.; Merighi, S.; Gessi, S.; Borea, P.A.; Varani, K.; Vincenzi, F. Adenosine Receptors in Neuropsychiatric Disorders: Fine Regulators of Neurotransmission and Potential Therapeutic Targets. Int. J. Mol. Sci. 2022, 23, 1219. [Google Scholar] [CrossRef]
- Burnstock, G. P2X Ion Channel Receptors and Inflammation. Purinergic Signal. 2016, 12, 59–67. [Google Scholar] [CrossRef]
- Wyatt, L.R.; Godar, S.C.; Khoja, S.; Jakowec, M.W.; Alkana, R.L.; Bortolato, M.; Davies, D.L. Sociocommunicative and Sensorimotor Impairments in Male P2X4-Deficient Mice. Neuropsychopharmacology 2013, 38, 1993–2002. [Google Scholar] [CrossRef]
- Szabó, D.; Tod, P.; Gölöncsér, F.; Román, V.; Lendvai, B.; Otrokocsi, L.; Sperlágh, B. Maternal P2X7 Receptor Inhibition Prevents Autism-like Phenotype in Male Mouse Offspring through the NLRP3-IL-1β Pathway. Brain Behav. Immun. 2022, 101, 318–332. [Google Scholar] [CrossRef]
- Horváth, G.; Otrokocsi, L.; Beko, K.; Baranyi, M.; Kittel, Á.; Fritz-Ruenes, P.A.; Sperlágh, B. P2X7 Receptors Drive Poly(I:C) Induced Autism-like Behavior in Mice. J. Neurosci. 2019, 39, 2542–2561. [Google Scholar] [CrossRef]
- Baines, K.J.; Hillier, D.M.; Haddad, F.L.; Rajakumar, N.; Schmid, S.; Renaud, S.J. Maternal Immune Activation Alters Fetal Brain Development and Enhances Proliferation of Neural Precursor Cells in Rats. Front. Immunol. 2020, 11, 1145. [Google Scholar] [CrossRef]
- Naviaux, R.K.; Zolkipli, Z.; Wang, L.; Nakayama, T.; Naviaux, J.C.; Le, T.P.; Schuchbauer, M.A.; Rogac, M.; Tang, Q.; Dugan, L.L.; et al. Antipurinergic Therapy Corrects the Autism-like Features in the Poly(IC) Mouse Model. PLoS ONE 2013, 8, e57380. [Google Scholar] [CrossRef]
- Zhang, W.; Xiao, D.; Mao, Q.; Xia, H. Role of Neuroinflammation in Neurodegeneration Development. Signal Transduct. Target. Ther. 2023, 8, 267. [Google Scholar] [CrossRef]
- Kumagawa, T.; Moro, N.; Maeda, T.; Kobayashi, M.; Furukawa, Y.; Shijo, K.; Yoshino, A. Anti-Inflammatory Effect of P2Y1 Receptor Blocker MRS2179 in a Rat Model of Traumatic Brain Injury. Brain Res. Bull. 2022, 181, 46–54. [Google Scholar] [CrossRef]
- Nisar, S.; Bhat, A.A.; Masoodi, T.; Hashem, S.; Akhtar, S.; Ali, T.A.; Amjad, S.; Chawla, S.; Bagga, P.; Frenneaux, M.P.; et al. Genetics of Glutamate and Its Receptors in Autism Spectrum Disorder. Mol. Psychiatry 2022, 27, 2380–2392. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, R.J.; Figueira, A.S.; Marques, J.M. P2Y1 Receptor as a Catalyst of Brain Neurodegeneration. NeuroSci 2022, 3, 604–615. [Google Scholar] [CrossRef]
- Jing, F.; Zhang, Y.; Long, T.; He, W.; Qin, G.; Zhang, D.; Chen, L.; Zhou, J. P2Y12 Receptor Mediates Microglial Activation via RhoA/ROCK Pathway in the Trigeminal Nucleus Caudalis in a Mouse Model of Chronic Migraine. J. Neuroinflamm. 2019, 16, 217. [Google Scholar] [CrossRef] [PubMed]
- Dutto, I.; Gerhards, J.; Herrera, A.; Souckova, O.; Škopová, V.; Smak, J.A.; Junza, A.; Yanes, O.; Boeckx, C.; Burkhalter, M.D.; et al. Pathway-Specific Effects of ADSL Deficiency on Neurodevelopment. Elife 2022, 11, e70518. [Google Scholar] [CrossRef] [PubMed]
- Jurecka, A.; Zikanova, M.; Kmoch, S.; Tylki-Szymańska, A. Adenylosuccinate Lyase Deficiency. J. Inherit. Metab. Dis. 2015, 38, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liang, S.; Wang, M.; Gao, J.; Sun, C.; Wang, J.; Xia, W.; Wu, S.; Sumner, S.J.; Zhang, F.; et al. Potential Serum Biomarkers from a Metabolomics Study of Autism. J. Psychiatry Neurosci. 2016, 41, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Sun, X.; Sun, C.; Zou, M.; Chen, Y.; Huang, J.; Wu, L.; Chen, W.-X. Prevalence of Epilepsy in Autism Spectrum Disorders: A Systematic Review and Meta-Analysis. Autism 2022, 26, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Sinha, J.K.; Ghosh, S.; Sharma, H.; Bhaskar, R.; Narayanan, K.B. A Comprehensive Review of Emerging Trends and Innovative Therapies in Epilepsy Management. Brain Sci. 2023, 13, 1305. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Sinha, J.K.; Khan, T.; Devaraju, K.S.; Singh, P.; Vaibhav, K.; Gaur, P. Pharmacological and Therapeutic Approaches in the Treatment of Epilepsy. Biomedicines 2021, 9, 470. [Google Scholar] [CrossRef]
- Spanoghe, J.; Larsen, L.E.; Craey, E.; Manzella, S.; Van Dycke, A.; Boon, P.; Raedt, R. The Signaling Pathways Involved in the Anticonvulsive Effects of the Adenosine A1 Receptor. Int. J. Mol. Sci. 2020, 22, 320. [Google Scholar] [CrossRef]
- Guo, M.; Xie, P.; Liu, S.; Luan, G.; Li, T. Epilepsy and Autism Spectrum Disorder (ASD): The Underlying Mechanisms and Therapy Targets Related to Adenosine. Curr. Neuropharmacol. 2023, 21, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Wong, Z.W.; Engel, T. More than a Drug Target: Purinergic Signalling as a Source for Diagnostic Tools in Epilepsy. Neuropharmacology 2023, 222, 109303. [Google Scholar] [CrossRef] [PubMed]
- Beamer, E.; Kuchukulla, M.; Boison, D.; Engel, T. ATP and Adenosine—Two Players in the Control of Seizures and Epilepsy Development. Prog. Neurobiol. 2021, 204, 102105. [Google Scholar] [CrossRef] [PubMed]
- Boison, D. The Adenosine Kinase Hypothesis of Epileptogenesis. Prog. Neurobiol. 2008, 84, 249–262. [Google Scholar] [CrossRef]
- Jimenez-Pacheco, A.; Diaz-Hernandez, M.; Arribas-Blázquez, M.; Sanz-Rodriguez, A.; Olivos-Oré, L.A.; Artalejo, A.R.; Alves, M.; Letavic, M.; Miras-Portugal, M.T.; Conroy, R.M.; et al. Transient P2X7 Receptor Antagonism Produces Lasting Reductions in Spontaneous Seizures and Gliosis in Experimental Temporal Lobe Epilepsy. J. Neurosci. 2016, 36, 5920–5932. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Bravo, M.; Carvalho, J.L.; Saldanha-Araujo, F. Adenosine Production: A Common Path for Mesenchymal Stem-Cell and Regulatory T-Cell-Mediated Immunosuppression. Purinergic Signal. 2016, 12, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Lanser, A.J.; Rezende, R.M.; Rubino, S.; Lorello, P.J.; Donnelly, D.J.; Xu, H.; Lau, L.A.; Dulla, C.G.; Caldarone, B.J.; Robson, S.C.; et al. Disruption of the ATP/Adenosine Balance in CD39-/- Mice Is Associated with Handling-Induced Seizures. Immunology 2017, 152, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-Z.; Huang, Y.; Hong, C.-G.; Wang, X.; Duan, R.; Liu, J.-Y.; He, J.-L.; Duan, D.; Xie, H.; Lu, M. Autologous Olfactory Mucosa Mesenchymal Stem Cells Treatment Improves the Neural Network in Chronic Refractory Epilepsy. Stem Cell Res. Ther. 2023, 14, 237. [Google Scholar] [CrossRef]
- Hlebokazov, F.; Dakukina, T.; Potapnev, M.; Kosmacheva, S.; Moroz, L.; Misiuk, N.; Golubeva, T.; Slobina, E.; Krasko, O.; Shakhbazau, A.; et al. Clinical Benefits of Single vs Repeated Courses of Mesenchymal Stem Cell Therapy in Epilepsy Patients. Clin. Neurol. Neurosurg. 2021, 207, 106736. [Google Scholar] [CrossRef]
- Rakofsky, J.; Rapaport, M. Mood Disorders. Contin. Lifelong Learn. Neurol. 2018, 24, 804–827. [Google Scholar] [CrossRef]
- Jones, B.D.M.; Daskalakis, Z.J.; Carvalho, A.F.; Strawbridge, R.; Young, A.H.; Mulsant, B.H.; Husain, M.I. Inflammation as a Treatment Target in Mood Disorders: Review. BJPsych Open 2020, 6, e60. [Google Scholar] [CrossRef] [PubMed]
- Oakley, B.; Loth, E.; Murphy, D.G. Autism and Mood Disorders. Int. Rev. Psychiatry 2021, 33, 280–299. [Google Scholar] [CrossRef] [PubMed]
- Etyemez, S.; Saleh, A.; Hamilton, J.E.; Koshy, A.J.; Abraham, J.E.; Selek, S. Higher Prevalence of Mood Disorders in Admitted Patients with Autism. Neurol. Psychiatry Brain Res. 2020, 37, 87–90. [Google Scholar] [CrossRef]
- Ortiz, R.; Ulrich, H.; Zarate, C.A.; Machado-Vieira, R. Purinergic System Dysfunction in Mood Disorders: A Key Target for Developing Improved Therapeutics. Prog. Neuropsychopharmacol. Biol. Psychiatry 2015, 57, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Bartoli, F.; Burnstock, G.; Crocamo, C.; Carrà, G. Purinergic Signaling and Related Biomarkers in Depression. Brain Sci. 2020, 10, 160. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, H.; Gao, H.; Zhang, H.; Zhang, H.; Wang, Q.; Sun, Z. P2X7 Receptor Mediates NLRP3 Inflammasome Activation in Depression and Diabetes. Cell Biosci. 2020, 10, 28. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Altarejos, P.; Cabrera-Pastor, A.; Martínez-García, M.; Sánchez-Huertas, C.; Hernández, A.; Moreno-Manzano, V.; Felipo, V. Extracellular Vesicles from Mesenchymal Stem Cells Reduce Neuroinflammation in Hippocampus and Restore Cognitive Function in Hyperammonemic Rats. J. Neuroinflamm. 2023, 20, 1. [Google Scholar] [CrossRef]
- Kin, K.; Yasuhara, T.; Borlongan, C.V.; Date, I. Encapsulated Stem Cells Ameliorate Depressive-like Behavior via Growth Factor Secretion. Brain Circ. 2018, 4, 128–132. [Google Scholar] [CrossRef]
- Del Puerto, A.; Wandosell, F.; Garrido, J.J. Neuronal and Glial Purinergic Receptors Functions in Neuron Development and Brain Disease. Front. Cell. Neurosci. 2013, 7, 197. [Google Scholar] [CrossRef]
- Sun, B.-X.; Peng, A.-S.; Liu, P.-J.; Wang, M.-J.; Ding, H.-L.; Hu, Y.-S.; Kang, L. Neuroprotection of Exercise: P2X4R and P2X7R Regulate BDNF Actions. Purinergic Signal. 2023, 19, 297–303. [Google Scholar] [CrossRef]
- Li, J.; Wang, H.; Du, C.; Jin, X.; Geng, Y.; Han, B.; Ma, Q.; Li, Q.; Wang, Q.; Guo, Y.; et al. hUC-MSCs Ameliorated CUMS-Induced Depression by Modulating Complement C3 Signaling-Mediated Microglial Polarization during Astrocyte-Microglia Crosstalk. Brain Res. Bull. 2020, 163, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Fang, Z.; Lin, L.; Xu, H.; Huang, Q.; Zhang, H. Plasma Complement C3 and C3a Are Increased in Major Depressive Disorder Independent of Childhood Trauma. BMC Psychiatry 2022, 22, 741. [Google Scholar] [CrossRef] [PubMed]
- Wyss, M.T.; Heuer, C.; Herwerth, M. The Bumpy Road of Purinergic Inhibitors to Clinical Application in Immune-Mediated Diseases. Neural Regen. Res. 2024, 19, 1206–1211. [Google Scholar] [CrossRef] [PubMed]
- Ranjbar, R.; Ghasemian, M.; Maniati, M.; Khatami, S.H.; Jamali, N.; Taheri-Anganeh, M. Gastrointestinal Disorder Biomarkers. Clin. Chim. Acta 2022, 530, 13–26. [Google Scholar] [CrossRef]
- Penzol, M.J.; Salazar de Pablo, G.; Llorente, C.; Moreno, C.; Hernández, P.; Dorado, M.L.; Parellada, M. Functional Gastrointestinal Disease in Autism Spectrum Disorder: A Retrospective Descriptive Study in a Clinical Sample. Front. Psychiatry 2019, 10, 179. [Google Scholar] [CrossRef] [PubMed]
- Vuerich, M.; Mukherjee, S.; Robson, S.C.; Longhi, M.S. Control of Gut Inflammation by Modulation of Purinergic Signaling. Front. Immunol. 2020, 11, 1882. [Google Scholar] [CrossRef]
- Diezmos, E.F.; Bertrand, P.P.; Liu, L. Purinergic Signaling in Gut Inflammation: The Role of Connexins and Pannexins. Front. Neurosci. 2016, 10, 311. [Google Scholar] [CrossRef] [PubMed]
- Figliuolo, V.R.; Savio, L.E.B.; Safya, H.; Nanini, H.; Bernardazzi, C.; Abalo, A.; de Souza, H.S.P.; Kanellopoulos, J.; Bobé, P.; Coutinho, C.M.L.M.; et al. P2X7 Receptor Promotes Intestinal Inflammation in Chemically Induced Colitis and Triggers Death of Mucosal Regulatory T Cells. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Weilinger, N.L.; Yang, K.; Choi, H.B.; Groten, C.J.; Wendt, S.; Murugan, M.; Wicki-Stordeur, L.E.; Bernier, L.-P.; Velayudhan, P.S.; Zheng, J.; et al. Pannexin-1 Opening in Neuronal Edema Causes Cell Death but Also Leads to Protection via Increased Microglia Contacts. Cell Rep. 2023, 42, 113128. [Google Scholar] [CrossRef]
- Eiro, N.; Fraile, M.; González-Jubete, A.; González, L.O.; Vizoso, F.J. Mesenchymal (Stem) Stromal Cells Based as New Therapeutic Alternative in Inflammatory Bowel Disease: Basic Mechanisms, Experimental and Clinical Evidence, and Challenges. Int. J. Mol. Sci. 2022, 23, 8905. [Google Scholar] [CrossRef]
- Zhang, J.; Lv, S.; Liu, X.; Song, B.; Shi, L. Umbilical Cord Mesenchymal Stem Cell Treatment for Crohn’s Disease: A Randomized Controlled Clinical Trial. Gut Liver 2018, 12, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Barnhoorn, M.C.; Wasser, M.N.J.M.; Roelofs, H.; Maljaars, P.W.J.; Molendijk, I.; Bonsing, B.A.; Oosten, L.E.M.; Dijkstra, G.; van der Woude, C.J.; Roelen, D.L.; et al. Long-Term Evaluation of Allogeneic Bone Marrow-Derived Mesenchymal Stromal Cell Therapy for Crohn’s Disease Perianal Fistulas. J. Crohns Colitis 2020, 14, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Cicchinelli, S.; Pignataro, G.; Gemma, S.; Piccioni, A.; Picozzi, D.; Ojetti, V.; Franceschi, F.; Candelli, M. PAMPs and DAMPs in Sepsis: A Review of Their Molecular Features and Potential Clinical Implications. Int. J. Mol. Sci. 2024, 25, 962. [Google Scholar] [CrossRef] [PubMed]
- López-García, L.; Castro-Manrreza, M.E. TNF-α and IFN-γ Participate in Improving the Immunoregulatory Capacity of Mesenchymal Stem/Stromal Cells: Importance of Cell-Cell Contact and Extracellular Vesicles. Int. J. Mol. Sci. 2021, 22, 9531. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Chiu, S.M.; Motan, D.A.L.; Zhang, Z.; Chen, L.; Ji, H.-L.; Tse, H.-F.; Fu, Q.-L.; Lian, Q. Mesenchymal Stem Cells and Immunomodulation: Current Status and Future Prospects. Cell Death Dis. 2016, 7, e2062. [Google Scholar] [CrossRef] [PubMed]
- English, K.; Barry, F.P.; Field-Corbett, C.P.; Mahon, B.P. IFN-Gamma and TNF-Alpha Differentially Regulate Immunomodulation by Murine Mesenchymal Stem Cells. Immunol. Lett. 2007, 110, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Lotfi, R.; Steppe, L.; Hang, R.; Rojewski, M.; Massold, M.; Jahrsdörfer, B.; Schrezenmeier, H. ATP Promotes Immunosuppressive Capacities of Mesenchymal Stromal Cells by Enhancing the Expression of Indoleamine Dioxygenase. Immun. Inflamm. Dis. 2018, 6, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.Y.; Wang, L.; Zemskova, M.; Deppen, J.; Xu, K.; Strobel, F.; García, A.J.; Tirouvanziam, R.; Levit, R.D. Adenosine Production by Biomaterial-Supported Mesenchymal Stromal Cells Reduces the Innate Inflammatory Response in Myocardial Ischemia/Reperfusion Injury. J. Am. Heart Assoc. 2018, 7, e006949. [Google Scholar] [CrossRef] [PubMed]
- Masoodifar, M.; Hajihashemi, S.; Pazhoohan, S.; Nazemi, S.; Mojadadi, M.-S. Effect of the Conditioned Medium of Mesenchymal Stem Cells on the Expression Levels of P2X4 and P2X7 Purinergic Receptors in the Spinal Cord of Rats with Neuropathic Pain. Purinergic Signal. 2021, 17, 143–150. [Google Scholar] [CrossRef]
- Burr, A.; Parekkadan, B. Kinetics of MSC-Based Enzyme Therapy for Immunoregulation. J. Transl. Med. 2019, 17, 263. [Google Scholar] [CrossRef]
- Perets, N.; Betzer, O.; Shapira, R.; Brenstein, S.; Angel, A.; Sadan, T.; Ashery, U.; Popovtzer, R.; Offen, D. Golden Exosomes Selectively Target Brain Pathologies in Neurodegenerative and Neurodevelopmental Disorders. Nano Lett. 2019, 19, 3422–3431. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, E.A.; Orekhov, A.N. Monocyte Activation in Immunopathology: Cellular Test for Development of Diagnostics and Therapy. J. Immunol. Res. 2016, 2016, 4789279. [Google Scholar] [CrossRef] [PubMed]
- Austermann, J.; Roth, J.; Barczyk-Kahlert, K. The Good and the Bad: Monocytes’ and Macrophages’ Diverse Functions in Inflammation. Cells 2022, 11, 1979. [Google Scholar] [CrossRef]
- Teo, K.Y.W.; Zhang, S.; Loh, J.T.; Lai, R.C.; Hey, H.W.D.; Lam, K.-P.; Lim, S.K.; Toh, W.S. Mesenchymal Stromal Cell Exosomes Mediate M2-like Macrophage Polarization through CD73/Ecto-5′-Nucleotidase Activity. Pharmaceutics 2023, 15, 1489. [Google Scholar] [CrossRef] [PubMed]
- Monguió-Tortajada, M.; Roura, S.; Gálvez-Montón, C.; Franquesa, M.; Bayes-Genis, A.; Borràs, F.E. Mesenchymal Stem Cells Induce Expression of CD73 in Human Monocytes In Vitro and in a Swine Model of Myocardial Infarction In Vivo. Front. Immunol. 2017, 8, 1577. [Google Scholar] [CrossRef]
- Sattler, C.; Steinsdoerfer, M.; Offers, M.; Fischer, E.; Schierl, R.; Heseler, K.; Däubener, W.; Seissler, J. Inhibition of T-Cell Proliferation by Murine Multipotent Mesenchymal Stromal Cells Is Mediated by CD39 Expression and Adenosine Generation. Cell Transplant. 2011, 20, 1221–1230. [Google Scholar] [CrossRef]
- Saldanha-Araujo, F.; Ferreira, F.I.S.; Palma, P.V.; Araujo, A.G.; Queiroz, R.H.C.; Covas, D.T.; Zago, M.A.; Panepucci, R.A. Mesenchymal Stromal Cells Up-Regulate CD39 and Increase Adenosine Production to Suppress Activated T-Lymphocytes. Stem Cell Res. 2011, 7, 66–74. [Google Scholar] [CrossRef]
- Lee, J.J.; Jeong, H.J.; Kim, M.K.; Wee, W.R.; Lee, W.W.; Kim, S.U.; Sung, C.; Yang, Y.H. CD39-Mediated Effect of Human Bone Marrow-Derived Mesenchymal Stem Cells on the Human Th17 Cell Function. Purinergic Signal. 2014, 10, 357–365. [Google Scholar] [CrossRef]
- Mottahedin, A.; Ardalan, M.; Chumak, T.; Riebe, I.; Ek, J.; Mallard, C. Effect of Neuroinflammation on Synaptic Organization and Function in the Developing Brain: Implications for Neurodevelopmental and Neurodegenerative Disorders. Front. Cell. Neurosci. 2017, 11, 190. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, Z. The Impact of Microglia on Neurodevelopment and Brain Function in Autism. Biomedicines 2024, 12, 210. [Google Scholar] [CrossRef]
- Cieślik, M.; Gassowska-Dobrowolska, M.; Zawadzka, A.; Frontczak-Baniewicz, M.; Gewartowska, M.; Dominiak, A.; Czapski, G.A.; Adamczyk, A. The Synaptic Dysregulation in Adolescent Rats Exposed to Maternal Immune Activation. Front. Mol. Neurosci. 2020, 13, 555290. [Google Scholar] [CrossRef]
- Kumar, H.; Sharma, B.M.; Sharma, B. Benefits of Agomelatine in Behavioral, Neurochemical and Blood Brain Barrier Alterations in Prenatal Valproic Acid Induced Autism Spectrum Disorder. Neurochem. Int. 2015, 91, 34–45. [Google Scholar] [CrossRef]
- Takeda, K.; Watanabe, T.; Oyabu, K.; Tsukamoto, S.; Oba, Y.; Nakano, T.; Kubota, K.; Katsurabayashi, S.; Iwasaki, K. Valproic Acid-Exposed Astrocytes Impair Inhibitory Synapse Formation and Function. Sci. Rep. 2021, 11, 23. [Google Scholar] [CrossRef]
- Mauri, M.; Lentini, D.; Gravati, M.; Foudah, D.; Biella, G.; Costa, B.; Toselli, M.; Parenti, M.; Coco, S. Mesenchymal Stem Cells Enhance GABAergic Transmission in Co-Cultured Hippocampal Neurons. Mol. Cell. Neurosci. 2012, 49, 395–405. [Google Scholar] [CrossRef]
- Doshmanziari, M.; Shirian, S.; Kouchakian, M.-R.; Moniri, S.F.; Jangnoo, S.; Mohammadi, N.; Zafari, F. Mesenchymal Stem Cells Act as Stimulators of Neurogenesis and Synaptic Function in a Rat Model of Alzheimer’s Disease. Heliyon 2021, 7, e07996. [Google Scholar] [CrossRef]
- Xin, D.; Li, T.; Chu, X.; Ke, H.; Liu, D.; Wang, Z. MSCs-Extracellular Vesicles Attenuated Neuroinflammation, Synapse Damage and Microglial Phagocytosis after Hypoxia-Ischemia Injury by Preventing Osteopontin Expression. Pharmacol. Res. 2021, 164, 105322. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic Signalling: Therapeutic Developments. Front. Pharmacol. 2017, 8, 661. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, Y.; Wang, J.; Dong, L.; Liu, S.; Li, S.; Wu, Q. Purinergic Signaling: A Gatekeeper of Blood-Brain Barrier Permeation. Front. Pharmacol. 2023, 14, 1112758. [Google Scholar] [CrossRef]
- Oliveira-Giacomelli, Á.; Petiz, L.L.; Andrejew, R.; Turrini, N.; Silva, J.B.; Sack, U.; Ulrich, H. Role of P2X7 Receptors in Immune Responses During Neurodegeneration. Front. Cell. Neurosci. 2021, 15, 662935. [Google Scholar] [CrossRef]
- Grygorowicz, T.; Dąbrowska-Bouta, B.; Strużyńska, L. Administration of an Antagonist of P2X7 Receptor to EAE Rats Prevents a Decrease of Expression of Claudin-5 in Cerebral Capillaries. Purinergic Signal. 2018, 14, 385–393. [Google Scholar] [CrossRef]
- Bynoe, M.S.; Viret, C.; Yan, A.; Kim, D.-G. Adenosine Receptor Signaling: A Key to Opening the Blood-Brain Door. Fluids Barriers CNS 2015, 12, 20. [Google Scholar] [CrossRef]
- Carman, A.J.; Mills, J.H.; Krenz, A.; Kim, D.-G.; Bynoe, M.S. Adenosine Receptor Signaling Modulates Permeability of the Blood-Brain Barrier. J. Neurosci. 2011, 31, 13272–13280. [Google Scholar] [CrossRef]
- Kim, D.-G.; Bynoe, M.S. A2A Adenosine Receptor Regulates the Human Blood-Brain Barrier Permeability. Mol. Neurobiol. 2015, 52, 664–678. [Google Scholar] [CrossRef]
- Mills, J.H.; Thompson, L.F.; Mueller, C.; Waickman, A.T.; Jalkanen, S.; Niemela, J.; Airas, L.; Bynoe, M.S. CD73 Is Required for Efficient Entry of Lymphocytes into the Central Nervous System during Experimental Autoimmune Encephalomyelitis. Proc. Natl. Acad. Sci. USA 2008, 105, 9325–9330. [Google Scholar] [CrossRef]
- Park, H.J.; Shin, J.Y.; Kim, H.N.; Oh, S.H.; Song, S.K.; Lee, P.H. Mesenchymal Stem Cells Stabilize the Blood-Brain Barrier through Regulation of Astrocytes. Stem Cell Res. Ther. 2015, 6, 187. [Google Scholar] [CrossRef]
- Jingyi, L.; Lin, W.; Yuan, C.; Lingling, Z.; Qianqian, J.; Anlong, X.; Yansong, G. Intravenous Transplantation of Bone Marrow-Derived Mesenchymal Stem Cells Improved Behavioral Deficits and Altered Fecal Microbiota Composition of BTBR Mice. Life Sci. 2024, 336, 122330. [Google Scholar] [CrossRef]
- Segal-Gavish, H.; Karvat, G.; Barak, N.; Barzilay, R.; Ganz, J.; Edry, L.; Aharony, I.; Offen, D.; Kimchi, T. Mesenchymal Stem Cell Transplantation Promotes Neurogenesis and Ameliorates Autism Related Behaviors in BTBR Mice. Autism Res. 2016, 9, 17–32. [Google Scholar] [CrossRef]
- Gobshtis, N.; Tfilin, M.; Wolfson, M.; Fraifeld, V.E.; Turgeman, G. Transplantation of Mesenchymal Stem Cells Reverses Behavioural Deficits and Impaired Neurogenesis Caused by Prenatal Exposure to Valproic Acid. Oncotarget 2017, 8, 17443–17452. [Google Scholar] [CrossRef]
- Perets, N.; Hertz, S.; London, M.; Offen, D. Intranasal Administration of Exosomes Derived from Mesenchymal Stem Cells Ameliorates Autistic-like Behaviors of BTBR Mice. Mol. Autism 2018, 9, 57. [Google Scholar] [CrossRef]
- Perets, N.; Oron, O.; Herman, S.; Elliott, E.; Offen, D. Exosomes Derived from Mesenchymal Stem Cells Improved Core Symptoms of Genetically Modified Mouse Model of Autism Shank3B. Mol. Autism 2020, 11, 65. [Google Scholar] [CrossRef]
- Panda, B.; Sharma, Y.; Gupta, S.; Mohanty, S. Mesenchymal Stem Cell-Derived Exosomes as an Emerging Paradigm for Regenerative Therapy and Nano-Medicine: A Comprehensive Review. Life 2021, 11, 784. [Google Scholar] [CrossRef]
- Sanchez-Diaz, M.; Quiñones-Vico, M.I.; Sanabria de la Torre, R.; Montero-Vílchez, T.; Sierra-Sánchez, A.; Molina-Leyva, A.; Arias-Santiago, S. Biodistribution of Mesenchymal Stromal Cells after Administration in Animal Models and Humans: A Systematic Review. J. Clin. Med. 2021, 10, 2925. [Google Scholar] [CrossRef]
- Poltavtseva, R.A.; Poltavtsev, A.V.; Lutsenko, G.V.; Svirshchevskaya, E.V. Myths, reality and future of mesenchymal stem cell therapy. Cell Tissue Res. 2019, 375, 563–574. [Google Scholar] [CrossRef]
- Tuazon, J.P.; Castelli, V.; Borlongan, C.V. Drug-like delivery methods of stem cells as biologics for stroke. Expert Opin. Drug Deliv. 2019, 16, 823–833. [Google Scholar] [CrossRef]
- Kandeel, M.; Morsy, M.A.; Alkhodair, K.M.; Alhojaily, S. Mesenchymal Stem Cell-Derived Extracellular Vesicles: An Emerging Diagnostic and Therapeutic Biomolecules for Neurodegenerative Disabilities. Biomolecules 2023, 13, 1250. [Google Scholar] [CrossRef]
- Noshadian, M.; Ragerdi Kashani, I.; Asadi-Golshan, R.; Zarini, D.; Ghafari, N.; Zahedi, E.; Pasbakhsh, P. Benefits of Bone Marrow Mesenchymal Stem Cells Compared to Their Conditioned Medium in Valproic Acid-Induced Autism in Rats. Mol. Biol. Rep. 2024, 51, 353. [Google Scholar] [CrossRef]
- Lv, Y.-T.; Zhang, Y.; Liu, M.; Qiuwaxi, J.; Ashwood, P.; Cho, S.C.; Huan, Y.; Ge, R.-C.; Chen, X.-W.; Wang, Z.-J.; et al. Transplantation of Human Cord Blood Mononuclear Cells and Umbilical Cord-Derived Mesenchymal Stem Cells in Autism. J. Transl. Med. 2013, 11, 196. [Google Scholar] [CrossRef]
- Sun, J.M.; Dawson, G.; Franz, L.; Howard, J.; McLaughlin, C.; Kistler, B.; Waters-Pick, B.; Meadows, N.; Troy, J.; Kurtzberg, J. Infusion of Human Umbilical Cord Tissue Mesenchymal Stromal Cells in Children with Autism Spectrum Disorder. Stem Cells Transl. Med. 2020, 9, 1137–1146. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wikarska, A.; Roszak, K.; Roszek, K. Mesenchymal Stem Cells and Purinergic Signaling in Autism Spectrum Disorder: Bridging the Gap between Cell-Based Strategies and Neuro-Immune Modulation. Biomedicines 2024, 12, 1310. https://doi.org/10.3390/biomedicines12061310
Wikarska A, Roszak K, Roszek K. Mesenchymal Stem Cells and Purinergic Signaling in Autism Spectrum Disorder: Bridging the Gap between Cell-Based Strategies and Neuro-Immune Modulation. Biomedicines. 2024; 12(6):1310. https://doi.org/10.3390/biomedicines12061310
Chicago/Turabian StyleWikarska, Agata, Kacper Roszak, and Katarzyna Roszek. 2024. "Mesenchymal Stem Cells and Purinergic Signaling in Autism Spectrum Disorder: Bridging the Gap between Cell-Based Strategies and Neuro-Immune Modulation" Biomedicines 12, no. 6: 1310. https://doi.org/10.3390/biomedicines12061310
APA StyleWikarska, A., Roszak, K., & Roszek, K. (2024). Mesenchymal Stem Cells and Purinergic Signaling in Autism Spectrum Disorder: Bridging the Gap between Cell-Based Strategies and Neuro-Immune Modulation. Biomedicines, 12(6), 1310. https://doi.org/10.3390/biomedicines12061310