
MicroRNA Analysis of In Vitro Differentiation of Spermatogonial Stem Cells Using a 3D Human Testis Organoid System
 , , , and
, , , and
Abstract
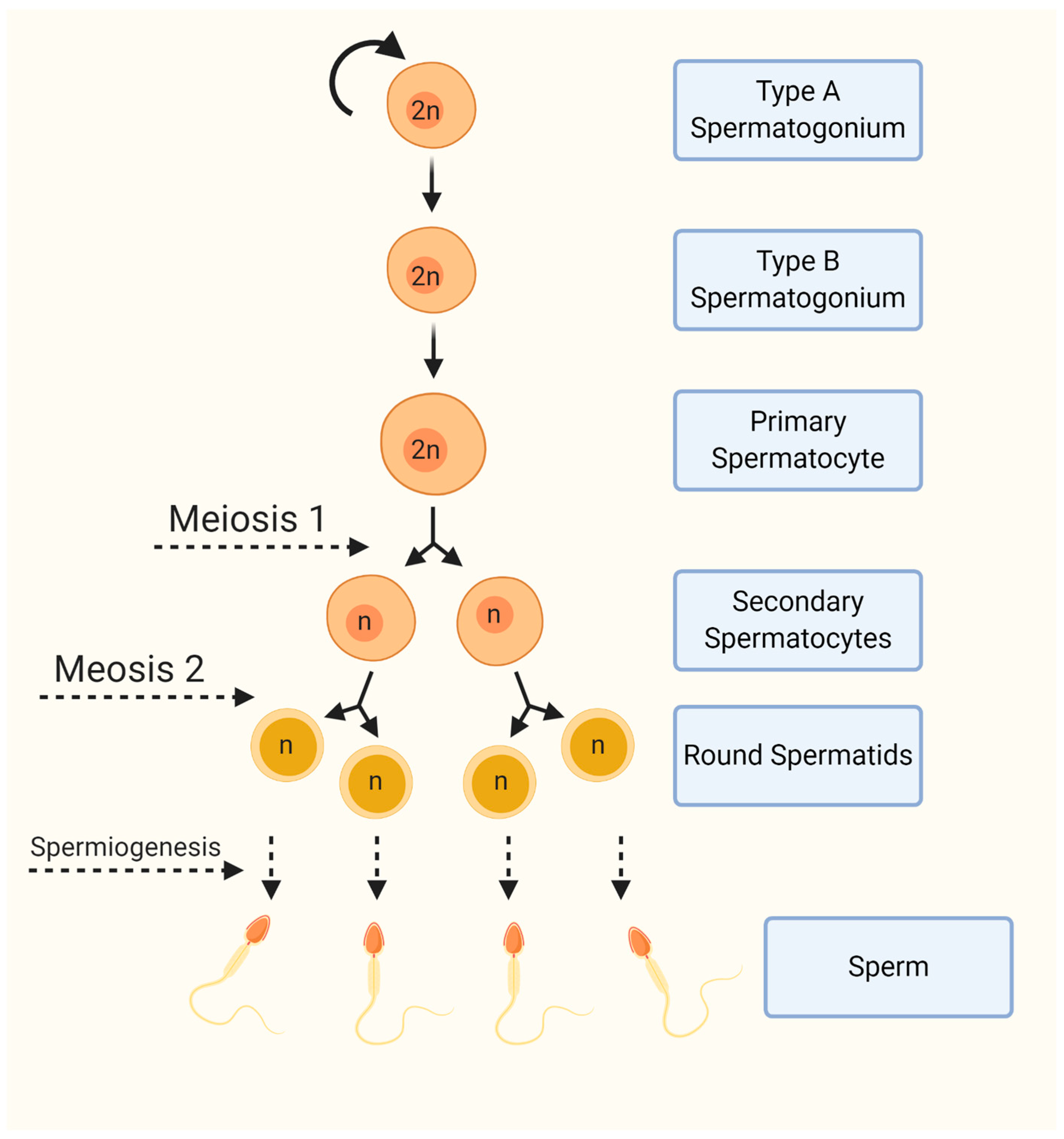
:1. Introduction
2. Materials and Methods
2.1. Human Materials
2.2. Testicular Cell Isolation and Culture
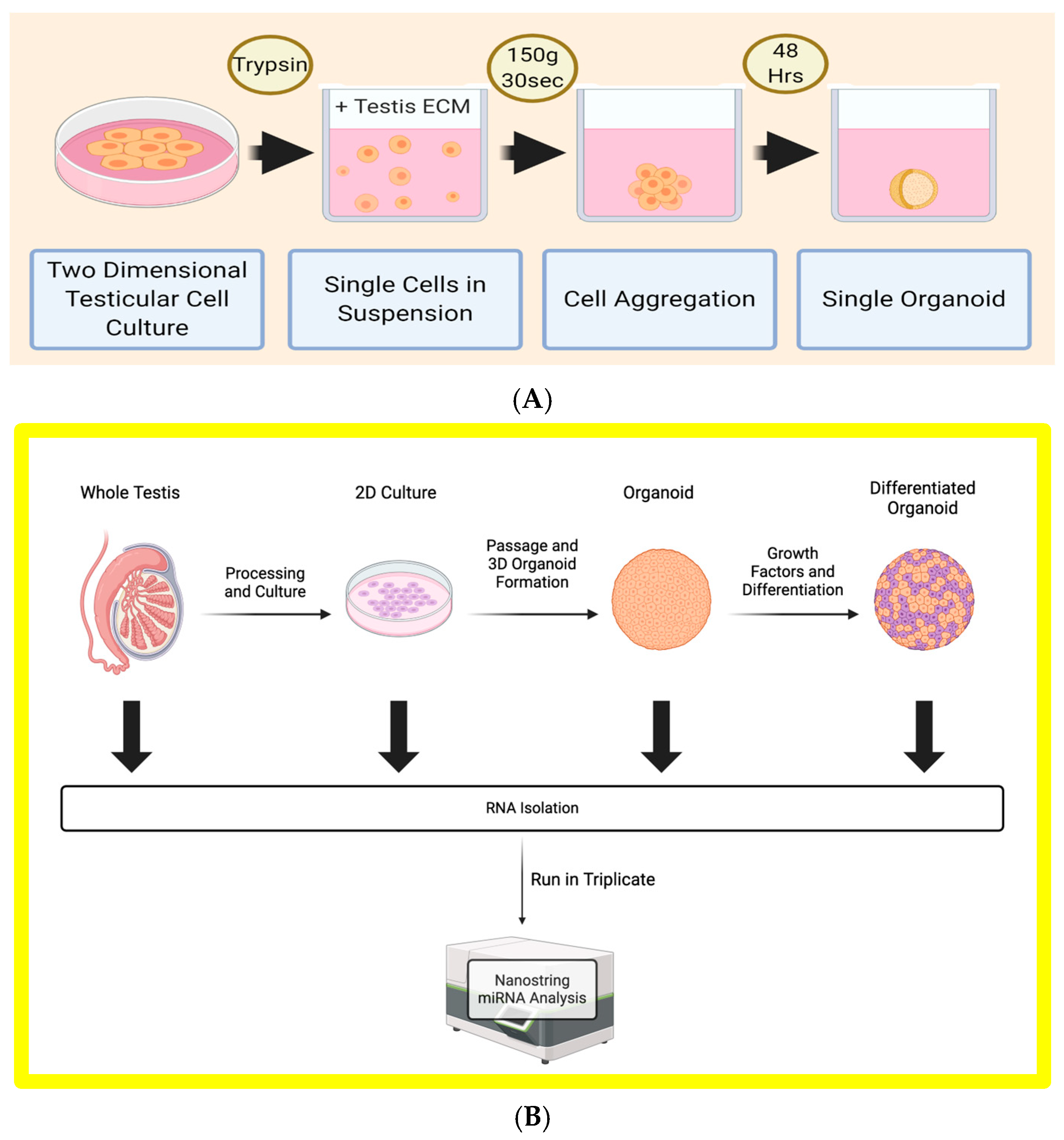
2.3. Formation of Three-Dimensional Human Testi Organoids (HTOs)
2.4. Organoid Differentiation
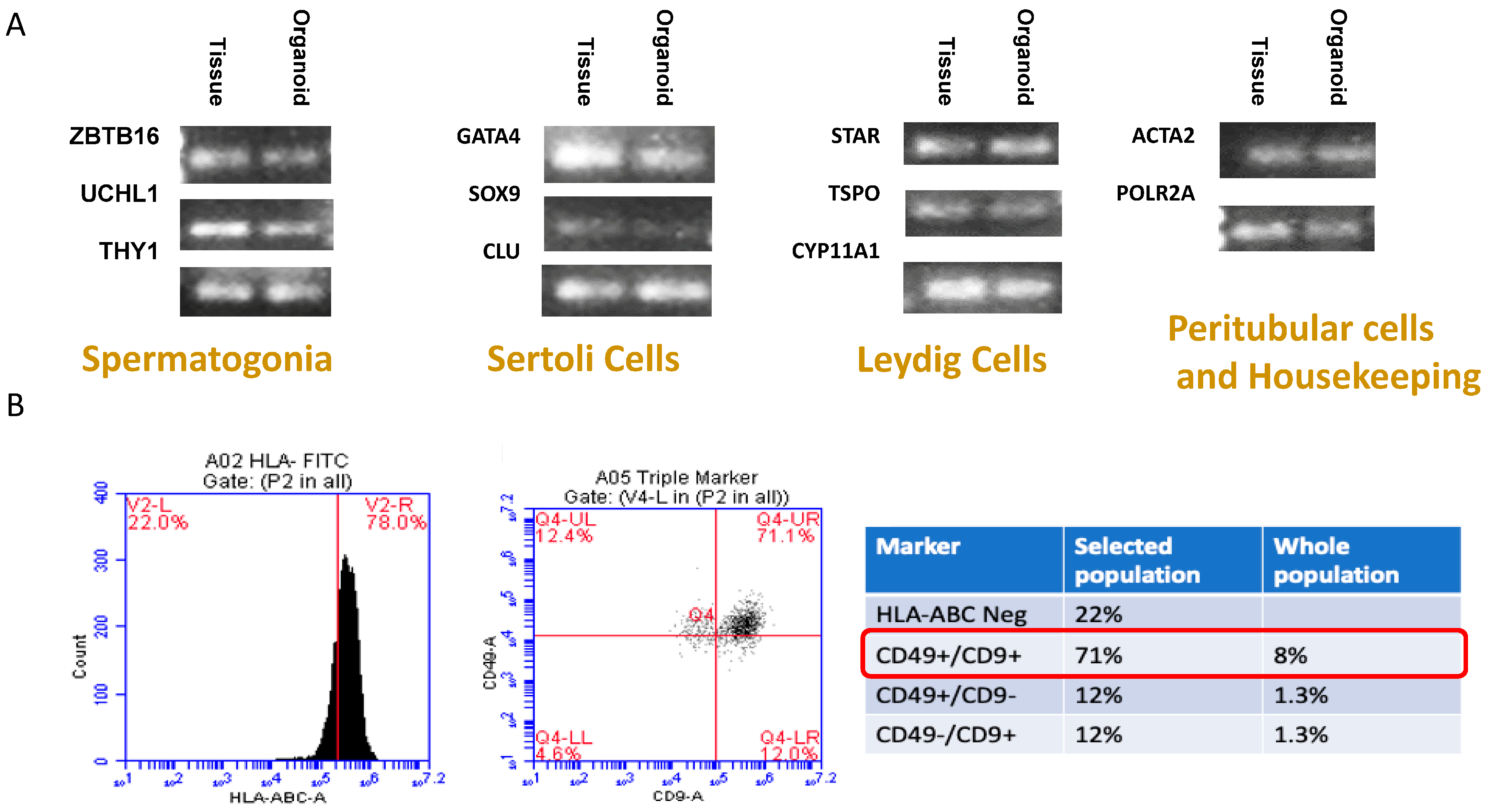
2.5. Testicular Cell Culture Characterization
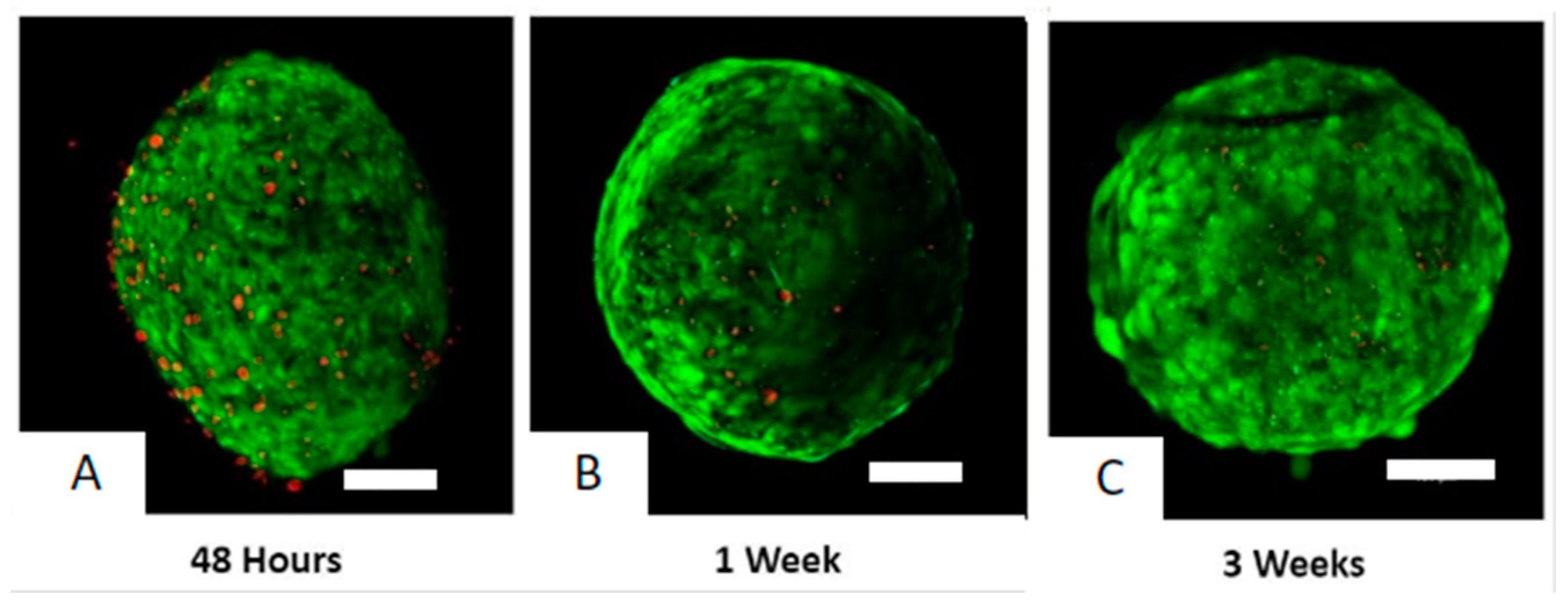
2.6. Organoid Viability Assays
2.7. miRNA Analysis via Nanostring Sprint Profiler
2.8. Data Analysis
3. Results
3.1. Organoid Viability
3.2. Culture Characterization
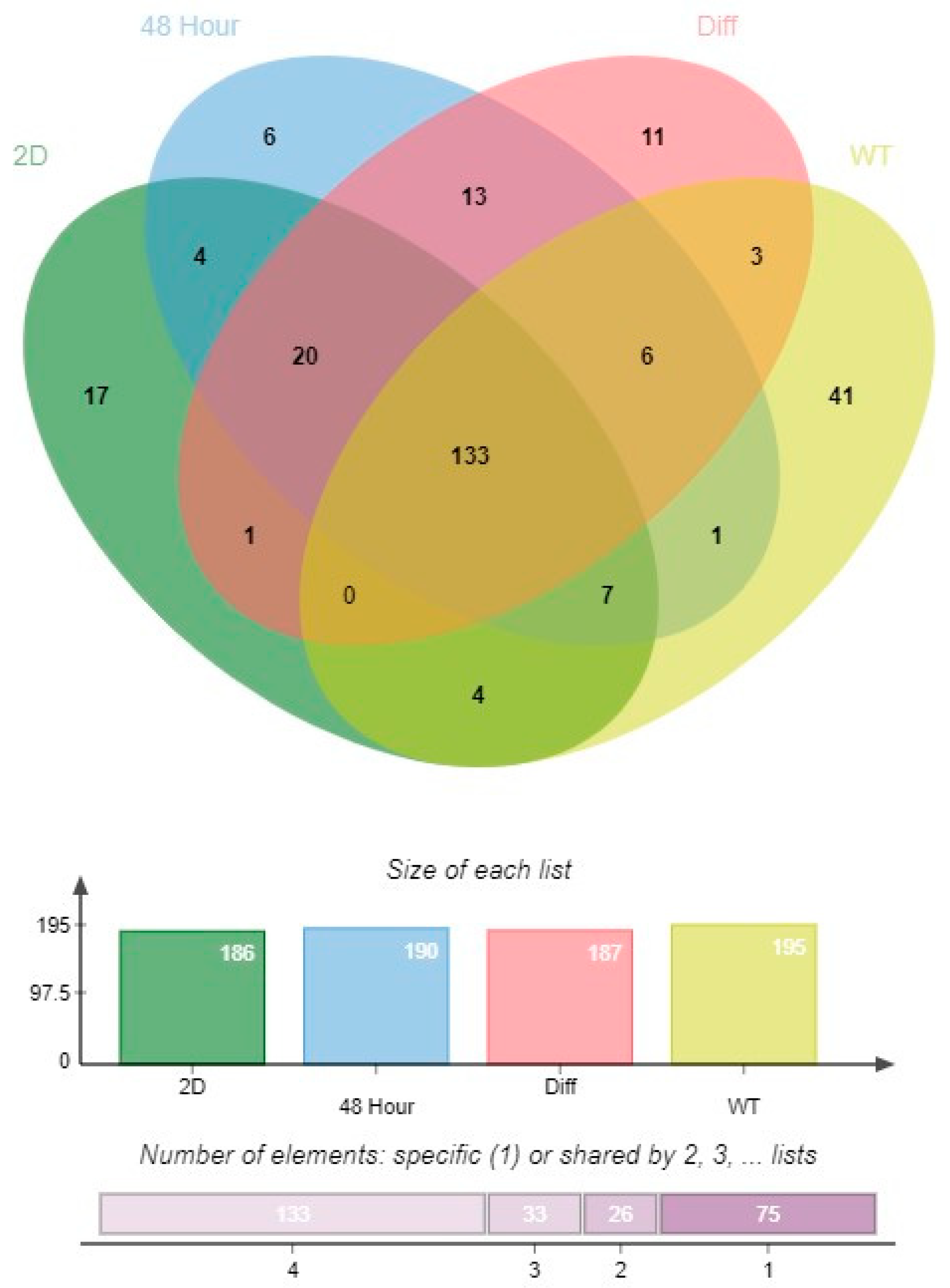
3.3. miRNA Expression Profiles
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kanatsu-Shinohara, M.; Shinohara, T. Spermatogonial stem cell self-renewal and development. Annu. Rev. Cell Dev. Biol. 2013, 29, 163–187. [Google Scholar] [CrossRef] [PubMed]
- Rato, L.; Alves, M.G.; Socorro, S.; Duarte, A.I.; Cavaco, J.E.; Oliveira, P.F. Metabolic regulation is important for spermatogenesis. Nat. Rev. Urol. 2012, 9, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Ruwanpura, S.M.; McLachlan, R.I.; Meachem, S.J. Hormonal regulation of male germ cell development. J. Endocrinol. 2010, 205, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Chalmel, F.; Rolland, A.D.; Niederhauser-Wiederkehr, C.; Chung, S.S.; Demougin, P.; Gattiker, A.; Moore, J.; Patard, J.J.; Wolgemuth, D.J.; Jegou, B.; et al. The conserved transcriptome in human and rodent male gametogenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 8346–8351. [Google Scholar] [CrossRef] [PubMed]
- Laiho, A.; Kotaja, N.; Gyenesei, A.; Sironen, A. Transcriptome profiling of the murine testis during the first wave of spermatogenesis. PLoS ONE 2013, 8, e61558. [Google Scholar] [CrossRef] [PubMed]
- Guttman, M.; Rinn, J.L. Modular regulatory principles of large non-coding RNAs. Nature 2012, 482, 339–346. [Google Scholar] [CrossRef]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef]
- Brannan, C.I.; Dees, E.C.; Ingram, R.S.; Tilghman, S.M. The product of the H19 gene may function as an RNA. Mol. Cell Biol. 1990, 10, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.J.; Ballabio, A.; Rupert, J.L.; Lafreniere, R.G.; Grompe, M.; Tonlorenzi, R.; Willard, H.F. A gene from the region of the human X inactivation centre is expressed exclusively from the inactive X chromosome. Nature 1991, 349, 38–44. [Google Scholar] [CrossRef]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E.; et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar] [CrossRef]
- Eddy, S.R. Non-coding RNA genes and the modern RNA world. Nat. Rev. Genet. 2001, 2, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Khawar, M.B.; Mehmood, R.; Roohi, N. MicroRNAs: Recent insights towards their role in male infertility and reproductive cancers. Bosn. J. Basic Med. Sci. 2019, 19, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Kotaja, N. MicroRNAs and spermatogenesis. Fertil. Steril. 2014, 101, 1552–1562. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Carninci, P.; Kasukawa, T.; Katayama, S.; Gough, J.; Frith, M.C.; Maeda, N.; Oyama, R.; Ravasi, T.; Lenhard, B.; Wells, C.; et al. The transcriptional landscape of the mammalian genome. Science 2005, 309, 1559–1563. [Google Scholar] [CrossRef] [PubMed]
- Castellano, L.; Stebbing, J. Deep sequencing of small RNAs identifies canonical and non-canonical miRNA and endogenous siRNAs in mammalian somatic tissues. Nucleic Acids Res. 2013, 41, 3339–3351. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef] [PubMed]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef]
- Pratt, A.J.; MacRae, I.J. The RNA-induced silencing complex: A versatile gene-silencing machine. J. Biol. Chem. 2009, 284, 17897–17901. [Google Scholar] [CrossRef]
- Holt, J.E.; Stanger, S.J.; Nixon, B.; McLaughlin, E.A. Non-coding RNA in Spermatogenesis and Epididymal Maturation. Adv. Exp. Med. Biol. 2016, 886, 95–120. [Google Scholar] [CrossRef] [PubMed]
- Joshi, M.; Sethi, S.; Mehta, P.; Kumari, A.; Rajender, S. Small RNAs, spermatogenesis, and male infertility: A decade of retrospect. Reprod. Biol. Endocrinol. 2023, 21, 106. [Google Scholar] [CrossRef] [PubMed]
- Novotny, G.W.; Sonne, S.B.; Nielsen, J.E.; Jonstrup, S.P.; Hansen, M.A.; Skakkebaek, N.E.; Rajpert-De Meyts, E.; Kjems, J.; Leffers, H. Translational repression of E2F1 mRNA in carcinoma in situ and normal testis correlates with expression of the miR-17-92 cluster. Cell Death Differ. 2007, 14, 879–882. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Sun, J.; Han, J.; Ma, Z.; Pan, M.; Du, Z. MiR-181a Promotes Spermatogenesis by Targeting the S6K1 Pathway. Int. J. Stem Cells 2021, 14, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Bouhallier, F.; Allioli, N.; Lavial, F.; Chalmel, F.; Perrard, M.H.; Durand, P.; Samarut, J.; Pain, B.; Rouault, J.P. Role of miR-34c microRNA in the late steps of spermatogenesis. RNA 2010, 16, 720–731. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.; Goodyear, S.M.; Rao, S.; Wu, X.; Tobias, J.W.; Avarbock, M.R.; Brinster, R.L. MicroRNA-21 regulates the self-renewal of mouse spermatogonial stem cells. Proc. Natl. Acad Sci. USA 2011, 108, 12740–12745. [Google Scholar] [CrossRef] [PubMed]
- Sadri-Ardekani, H.; Mizrak, S.C.; van Daalen, S.K.; Korver, C.M.; Roepers-Gajadien, H.L.; Koruji, M.; Hovingh, S.; de Reijke, T.M.; de la Rosette, J.J.; van der Veen, F.; et al. Propagation of human spermatogonial stem cells in vitro. JAMA 2009, 302, 2127–2134. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Katagiri, K.; Gohbara, A.; Inoue, K.; Ogonuki, N.; Ogura, A.; Kubota, Y.; Ogawa, T. In vitro production of functional sperm in cultured neonatal mouse testes. Nature 2011, 471, 504–507. [Google Scholar] [CrossRef]
- Sato, T.; Katagiri, K.; Yokonishi, T.; Kubota, Y.; Inoue, K.; Ogonuki, N.; Matoba, S.; Ogura, A.; Ogawa, T. In vitro production of fertile sperm from murine spermatogonial stem cell lines. Nat. Commun. 2011, 2, 472. [Google Scholar] [CrossRef]
- Baert, Y.; De Kock, J.; Alves-Lopes, J.P.; Soder, O.; Stukenborg, J.B.; Goossens, E. Primary Human Testicular Cells Self-Organize into Organoids with Testicular Properties. Stem Cell Rep. 2017, 8, 30–38. [Google Scholar] [CrossRef]
- Baert, Y.; Rombaut, C.; Goossens, E. Scaffold-Based and Scaffold-Free Testicular Organoids from Primary Human Testicular Cells. Methods Mol. Biol. 2019, 1576, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Richer, G.; Baert, Y.; Goossens, E. In-vitro spermatogenesis through testis modelling: Toward the generation of testicular organoids. Andrology 2019, 8, 879–891. [Google Scholar] [CrossRef] [PubMed]
- Pendergraft, S.S.; Sadri-Ardekani, H.; Atala, A.; Bishop, C.E. Three-dimensional testicular organoid: A novel tool for the study of human spermatogenesis and gonadotoxicity in vitro. Biol. Reprod. 2017, 96, 720–732. [Google Scholar] [CrossRef]
- van Pelt, A.M.; Morena, A.R.; van Dissel-Emiliani, F.M.; Boitani, C.; Gaemers, I.C.; de Rooij, D.G.; Stefanini, M. Isolation of the synchronized A spermatogonia from adult vitamin A-deficient rat testes. Biol. Reprod. 1996, 55, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Bardou, P.; Mariette, J.; Escudie, F.; Djemiel, C.; Klopp, C. jvenn: An interactive Venn diagram viewer. BMC Bioinform. 2014, 15, 293. [Google Scholar] [CrossRef] [PubMed]
- Tong, M.H.; Mitchell, D.; Evanoff, R.; Griswold, M.D. Expression of Mirlet7 family microRNAs in response to retinoic acid-induced spermatogonial differentiation in mice. Biol. Reprod. 2011, 85, 189–197. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Jiang, J.; Kokkinaki, M.; Tang, L.; Zeng, W.; Gallicano, I.; Dobrinski, I.; Dym, M. MiRNA-20 and mirna-106a regulate spermatogonial stem cell renewal at the post-transcriptional level via targeting STAT3 and Ccnd1. Stem Cells 2013, 31, 2205–2217. [Google Scholar] [CrossRef] [PubMed]
- Moritoki, Y.; Hayashi, Y.; Mizuno, K.; Kamisawa, H.; Nishio, H.; Kurokawa, S.; Ugawa, S.; Kojima, Y.; Kohri, K. Expression profiling of microRNA in cryptorchid testes: miR-135a contributes to the maintenance of spermatogonial stem cells by regulating FoxO1. J. Urol. 2014, 191, 1174–1180. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Hao, L.; Hu, F.; Dong, Y.; Gou, L.; Zhang, W.; Wang, X.; Zhao, Y.; Jia, M.; Hu, S.; et al. MicroRNA profiles and potential regulatory pattern during the early stage of spermatogenesis in mice. Sci. China Life Sci. 2015, 58, 442–450. [Google Scholar] [CrossRef]
- Rahbar, S.; Pashaiasl, M.; Ezzati, M.; Ahmadi AsrBadr, Y.; Mohammadi-Dehcheshmeh, M.; Mohammadi, S.A.; Ghaffari Novin, M. MicroRNA-based regulatory circuit involved in sperm infertility. Andrologia 2020, 52, e13453. [Google Scholar] [CrossRef]
- Yadav, S.K.; Pandey, A.; Kumar, L.; Devi, A.; Kushwaha, B.; Vishvkarma, R.; Maikhuri, J.P.; Rajender, S.; Gupta, G. The thermo-sensitive gene expression signatures of spermatogenesis. Reprod. Biol. Endocrinol. 2018, 16, 56. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Tang, C.; Zhang, Y.; Wu, J.; Bao, J.; Zheng, H.; Xu, C.; Yan, W. mir-34b/c and mir-449a/b/c are required for spermatogenesis, but not for the first cleavage division in mice. Biol. Open 2015, 4, 212–223. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Micro RNA | Accession # | Count: Differentiated HTO | SD | Count: Whole Testis | SD | Ratio: Differentiated HTO vs. Whole Testis | p Value | FDR Adjusted p Value |
|---|---|---|---|---|---|---|---|---|
| hsa-miR-10b-5p | MIMAT0000254 | 1200.23 | 60.36 | 383.78 | 16.77 | 3.13 | <0.001 | 0.02 |
| hsa-miR-21-5p | MIMAT0000076 | 16,760.51 | 2262.3 | 1388.78 | 246.48 | 12.07 | <0.001 | 0.04 |
| hsa-miR-191-5p | MIMAT0000440 | 1616.98 | 87.59 | 829.35 | 57.15 | 1.95 | <0.001 | 0.11 |
| hsa-miR-4443 | MIMAT0018961 | 137.95 | 6.13 | 36.62 | 3.53 | 3.77 | <0.001 | 0.13 |
| hsa-miR-34a-5p | MIMAT0000255 | 4227.95 | 968.56 | 514.26 | 90.02 | 8.22 | <0.001 | 0.13 |
| hsa-miR-149-5p | MIMAT0000450 | 14.41 | 2.1 | 44.99 | 5.38 | −3.12 | 0.001 | 0.15 |
| hsa-miR-100-5p | MIMAT0000098 | 7127.82 | 1549.68 | 599.72 | 53.35 | 11.89 | 0.001 | 0.15 |
| hsa-miR-20a-5p+hsa-miR-20b-5p | MIMAT0000075 | 185.38 | 18.53 | 756.22 | 22.04 | −4.08 | 0.001 | 0.18 |
| hsa-miR-4516 | MIMAT0019053 | 1319.43 | 936.22 | 16.81 | 10.91 | 78.49 | 0.001 | 0.18 |
| hsa-miR-199a-3p+hsa-miR-199b-3p | MIMAT0000232 | 16,327.05 | 3656.88 | 3713.74 | 744.86 | 4.4 | 0.001 | 0.21 |
| hsa-miR-132-3p | MIMAT0000426 | 1383.47 | 239.95 | 140.24 | 42.71 | 9.86 | 0.001 | 0.22 |
| hsa-miR-199b-5p | MIMAT0000263 | 3168.82 | 570.56 | 277.43 | 88.37 | 11.42 | 0.001 | 0.27 |
| hsa-miR-324-5p | MIMAT0000761 | 127.1 | 20.66 | 47.54 | 7.2 | 2.67 | 0.002 | 0.27 |
| hsa-miR-126-3p | MIMAT0000445 | 26.69 | 9.5 | 1407.69 | 1063.11 | −52.75 | 0.002 | 0.29 |
| hsa-miR-656-3p | MIMAT0003332 | 112.54 | 17.51 | 17.88 | 5.52 | 6.29 | 0.002 | 0.29 |
| hsa-miR-574-3p | MIMAT0003239 | 232.68 | 50.37 | 70.34 | 14.97 | 3.31 | 0.002 | 0.29 |
| hsa-miR-1915-3p | MIMAT0007892 | 326.46 | 154.96 | 10 | 6.76 | 32.65 | 0.002 | 0.29 |
| hsa-miR-28-5p | MIMAT0000085 | 243.28 | 34.32 | 107.6 | 14.82 | 2.26 | 0.002 | 0.29 |
| hsa-miR-299-5p | MIMAT0002890 | 255.86 | 47.42 | 88.84 | 11.52 | 2.88 | 0.003 | 0.34 |
| hsa-miR-494-3p | MIMAT0002816 | 85.63 | 19.8 | 21.8 | 2.89 | 3.93 | 0.003 | 0.35 |
| hsa-miR-374a-5p | MIMAT0000727 | 3429.96 | 329.54 | 788.5 | 178.22 | 4.35 | 0.003 | 0.36 |
| hsa-miR-454-3p | MIMAT0003885 | 36.02 | 7.62 | 11.21 | 1.15 | 3.21 | 0.003 | 0.38 |
| hsa-miR-19a-3p | MIMAT0000073 | 112.43 | 9.37 | 305.66 | 58.44 | −2.72 | 0.004 | 0.44 |
| hsa-miR-337-3p | MIMAT0000754 | 86.29 | 28.61 | 20.35 | 4.91 | 4.24 | 0.004 | 0.44 |
| hsa-let-7d-5p | MIMAT0000065 | 1133.24 | 190.53 | 545.61 | 76.95 | 2.08 | 0.004 | 0.48 |
| hsa-miR-24-3p | MIMAT0000080 | 347.75 | 90.56 | 90.88 | 27.43 | 3.83 | 0.005 | 0.5 |
| hsa-miR-497-5p | MIMAT0002820 | 165.53 | 74.19 | 1130.1 | 366.99 | −6.83 | 0.007 | 0.64 |
| hsa-let-7a-5p | MIMAT0000062 | 28,438.82 | 4218.04 | 6958.53 | 2037.02 | 4.09 | 0.007 | 0.66 |
| hsa-miR-340-5p | MIMAT0004692 | 39.73 | 1.37 | 60.28 | 5.17 | −1.52 | 0.007 | 0.66 |
| hsa-miR-382-5p | MIMAT0000737 | 410 | 52.18 | 69.43 | 24.2 | 5.91 | 0.007 | 0.67 |
| hsa-miR-199a-5p | MIMAT0000231 | 7133.99 | 2265.32 | 1925.62 | 241.68 | 3.7 | 0.008 | 0.68 |
| hsa-miR-4454+hsa-miR-7975 | MIMAT0018976 | 7363.49 | 3535.37 | 60,798.29 | 31,832.32 | −8.26 | 0.008 | 0.7 |
| hsa-miR-195-5p | MIMAT0000461 | 73.33 | 32.71 | 316.97 | 90.77 | −4.32 | 0.008 | 0.7 |
| hsa-miR-29b-3p | MIMAT0000100 | 10,994.69 | 3044.41 | 3690.16 | 844.93 | 2.98 | 0.008 | 0.7 |
| hsa-miR-411-5p | MIMAT0003329 | 252.66 | 50.11 | 71.26 | 23.61 | 3.55 | 0.010 | 0.78 |
| hsa-miR-181b-5p+hsa-miR-181d-5p | MIMAT0000257 | 128.86 | 18.38 | 13.64 | 6.68 | 9.45 | 0.012 | 0.92 |
| hsa-miR-214-3p | MIMAT0000271 | 440.69 | 182.59 | 85.74 | 7.23 | 5.14 | 0.013 | 0.93 |
| hsa-miR-140-5p | MIMAT0000431 | 219.29 | 31.37 | 134.62 | 20.04 | 1.63 | 0.014 | 0.98 |
| hsa-let-7e-5p | MIMAT0000066 | 1530.35 | 514.82 | 368.91 | 12.64 | 4.15 | 0.014 | 0.98 |
| hsa-miR-222-3p | MIMAT0000279 | 1291.16 | 137.36 | 99.33 | 72.13 | 13 | 0.016 | 1 |
| hsa-miR-125b-5p | MIMAT0000423 | 55,042.04 | 2371.08 | 14,946.62 | 4495.45 | 3.68 | 0.018 | 1 |
| hsa-let-7i-5p | MIMAT0000415 | 3609.14 | 1781.31 | 270.52 | 213.17 | 13.34 | 0.020 | 1 |
| hsa-miR-423-3p | MIMAT0001340 | 75.8 | 16.61 | 36.53 | 2.02 | 2.07 | 0.025 | 1 |
| hsa-miR-151a-5p | MIMAT0004697 | 111.64 | 9.04 | 61.83 | 12.31 | 1.81 | 0.027 | 1 |
| hsa-miR-93-5p | MIMAT0000093 | 475.06 | 115.72 | 254.65 | 52.54 | 1.87 | 0.027 | 1 |
| hsa-miR-181a-5p | MIMAT0000256 | 3259.44 | 852.17 | 1104.04 | 553.64 | 2.95 | 0.029 | 1 |
| hsa-miR-15a-5p | MIMAT0000068 | 1994.78 | 916.55 | 678.24 | 116.92 | 2.94 | 0.030 | 1 |
| hsa-miR-19b-3p | MIMAT0000074 | 322.42 | 111.7 | 929.39 | 74.56 | −2.88 | 0.030 | 1 |
| hsa-miR-25-3p | MIMAT0000081 | 967.8 | 180.1 | 546.29 | 19 | 1.77 | 0.030 | 1 |
| hsa-miR-155-5p | MIMAT0000646 | 156.89 | 54.94 | 30.87 | 18.87 | 5.08 | 0.032 | 1 |
| hsa-miR-337-5p | MIMAT0004695 | 132.27 | 28.81 | 32.91 | 21.57 | 4.02 | 0.032 | 1 |
| hsa-miR-379-5p | MIMAT0000733 | 579.51 | 97.5 | 193.29 | 93.59 | 3 | 0.032 | 1 |
| hsa-miR-125a-5p | MIMAT0000443 | 1721.59 | 105.77 | 572.19 | 206.74 | 3.01 | 0.036 | 1 |
| hsa-miR-362-3p | MIMAT0004683 | 196.94 | 89.63 | 56.66 | 2.75 | 3.48 | 0.037 | 1 |
| hsa-miR-409-3p | MIMAT0001639 | 287.22 | 68.71 | 22 | 20.61 | 13.06 | 0.040 | 1 |
| hsa-miR-23a-3p | MIMAT0000078 | 9417.15 | 422.13 | 3390.98 | 1375.87 | 2.78 | 0.040 | 1 |
| hsa-miR-130a-3p | MIMAT0000425 | 2008.96 | 43.27 | 1103.44 | 234.44 | 1.82 | 0.041 | 1 |
| hsa-miR-30a-3p | MIMAT0000088 | 100.16 | 21.95 | 42.26 | 16.07 | 2.37 | 0.044 | 1 |
| hsa-let-7g-5p | MIMAT0000414 | 3305.55 | 290.45 | 1874.25 | 434.71 | 1.76 | 0.045 | 1 |
| hsa-miR-107 | MIMAT0000104 | 236.27 | 24.03 | 110.42 | 34.11 | 2.14 | 0.046 | 1 |
| hsa-miR-500a-5p+hsa-miR-501-5p | MIMAT0004773 | 113.98 | 74.09 | 33.9 | 16.41 | 3.36 | 0.049 | 1 |
| Micro RNA | Accession # | Count: Differentiated HTO | SD | Count: Whole Testis | SD | Ratio: Differentiated HTO vs. Whole Testis | p Value | FDR Adjusted p Value |
|---|---|---|---|---|---|---|---|---|
| hsa-miR-30e-5p | MIMAT0000692 | 158.98 | 26.11 | 197.36 | 112.12 | −1.24 | 0.533 | 1 |
| hsa-miR-3144-3p | MIMAT0015015 | 35.92 | 17.54 | 44.6 | 48.22 | −1.24 | 0.726 | 1 |
| hsa-miR-18a-5p | MIMAT0000072 | 38.09 | 6.77 | 46.93 | 6.5 | −1.23 | 0.190 | 1 |
| hsa-miR-450a-5p | MIMAT0001545 | 1022.21 | 531.67 | 1243.36 | 268.27 | −1.22 | 0.585 | 1 |
| hsa-miR-335-5p | MIMAT0000765 | 31.79 | 19.97 | 37.85 | 6.93 | −1.19 | 0.655 | 1 |
| hsa-miR-1260a | MIMAT0005911 | 41.89 | 25.67 | 48.15 | 5.83 | −1.15 | 0.719 | 1 |
| hsa-miR-29c-3p | MIMAT0000681 | 1039.99 | 184.25 | 1188.48 | 265.07 | −1.14 | 0.446 | 1 |
| hsa-miR-612 | MIMAT0003280 | 168.68 | 61.15 | 193.04 | 235.53 | −1.14 | 0.829 | 1 |
| hsa-miR-148a-3p | MIMAT0000243 | 906.63 | 173.39 | 970.51 | 195.8 | −1.07 | 0.691 | 1 |
| hsa-miR-128-3p | MIMAT0000424 | 34.62 | 16.66 | 36.77 | 5.55 | −1.06 | 0.843 | 1 |
| hsa-miR-186-5p | MIMAT0000456 | 117.72 | 32.73 | 117.35 | 18.55 | 1 | 0.987 | 1 |
| hsa-miR-106b-5p | MIMAT0000680 | 93.14 | 59.01 | 93.02 | 26.47 | 1 | 0.997 | 1 |
| hsa-miR-450a-1-3p | MIMAT0022700 | 13.42 | 5.57 | 13.4 | 11.42 | 1 | 0.998 | 1 |
| hsa-miR-30d-5p | MIMAT0000245 | 190.08 | 46.72 | 189.11 | 48.36 | 1.01 | 0.981 | 1 |
| hsa-miR-23b-3p | MIMAT0000418 | 499.24 | 138.72 | 488.41 | 33.92 | 1.02 | 0.905 | 1 |
| hsa-let-7c-5p | MIMAT0000064 | 2970.6 | 648.38 | 2890.79 | 360.98 | 1.03 | 0.862 | 1 |
| hsa-miR-218-5p | MIMAT0000275 | 84.49 | 38.09 | 80.06 | 37.7 | 1.06 | 0.889 | 1 |
| hsa-miR-664a-3p | MIMAT0005949 | 69.42 | 17.13 | 64.64 | 32.73 | 1.07 | 0.829 | 1 |
| hsa-miR-548ar-5p | MIMAT0022265 | 43.61 | 2.38 | 40.82 | 40.42 | 1.07 | 0.911 | 1 |
| hsa-miR-30e-3p | MIMAT0000693 | 94.53 | 31.32 | 87.2 | 8.68 | 1.08 | 0.737 | 1 |
| hsa-miR-361-3p | MIMAT0004682 | 101.28 | 16.6 | 93.93 | 48.41 | 1.08 | 0.812 | 1 |
| hsa-miR-590-5p | MIMAT0003258 | 30.92 | 12.26 | 25.75 | 9.11 | 1.2 | 0.573 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cohen, A.B.; Nikmehr, B.; Abdelaal, O.A.; Escott, M.; Walker, S.J.; Atala, A.; Sadri-Ardekani, H. MicroRNA Analysis of In Vitro Differentiation of Spermatogonial Stem Cells Using a 3D Human Testis Organoid System. Biomedicines 2024, 12, 1774. https://doi.org/10.3390/biomedicines12081774
Cohen AB, Nikmehr B, Abdelaal OA, Escott M, Walker SJ, Atala A, Sadri-Ardekani H. MicroRNA Analysis of In Vitro Differentiation of Spermatogonial Stem Cells Using a 3D Human Testis Organoid System. Biomedicines. 2024; 12(8):1774. https://doi.org/10.3390/biomedicines12081774
Chicago/Turabian StyleCohen, Adam B., Banafsheh Nikmehr, Omar A. Abdelaal, Megan Escott, Stephen J. Walker, Anthony Atala, and Hooman Sadri-Ardekani. 2024. "MicroRNA Analysis of In Vitro Differentiation of Spermatogonial Stem Cells Using a 3D Human Testis Organoid System" Biomedicines 12, no. 8: 1774. https://doi.org/10.3390/biomedicines12081774