

Pyruvate Carboxylase Attenuates Myocardial Ischemia–Reperfusion Injury in Heart Transplantation via Wnt/β-Catenin-Mediated Glutamine Metabolism
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Reagents
2.3. Retrieval and Preservation of Donor Hearts
2.4. Heterotopic Heart Transplantation
2.5. Neonatal Rat Cardiomyocyte Isolation and Culture
2.6. In Vitro Hypoxia–Reoxygenation Model
2.7. Adenovirus Construction and Transduction
2.8. RNA Interference In Vitro
2.9. Luciferase Reporter Assay
2.10. Terminal Deoxynucleotidyl Transferase Dutp Nick-End Labeling Assay
2.11. Mitochondrial Oxidative Stress Evaluation, Inflammatory Cytokines Assessment, LDH and cTnI Detection
2.12. Intracellular Reactive Oxygen Species Detection
2.13. Real-Time Quantitative PCR
2.14. Western Blot Analysis
2.15. Statistical Analysis
3. Results
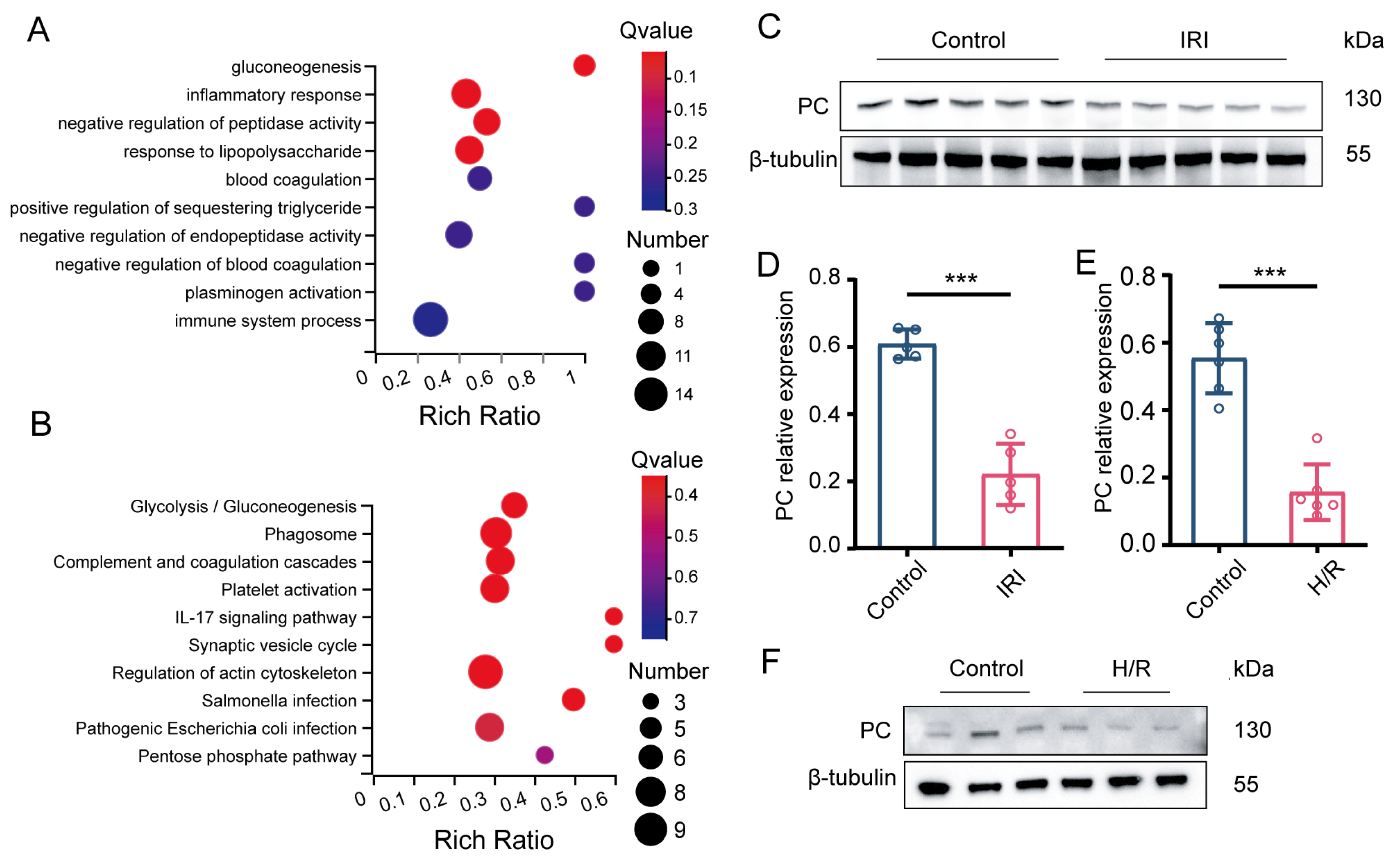
3.1. PC Is Significantly Downregulated during Myocardial IRI
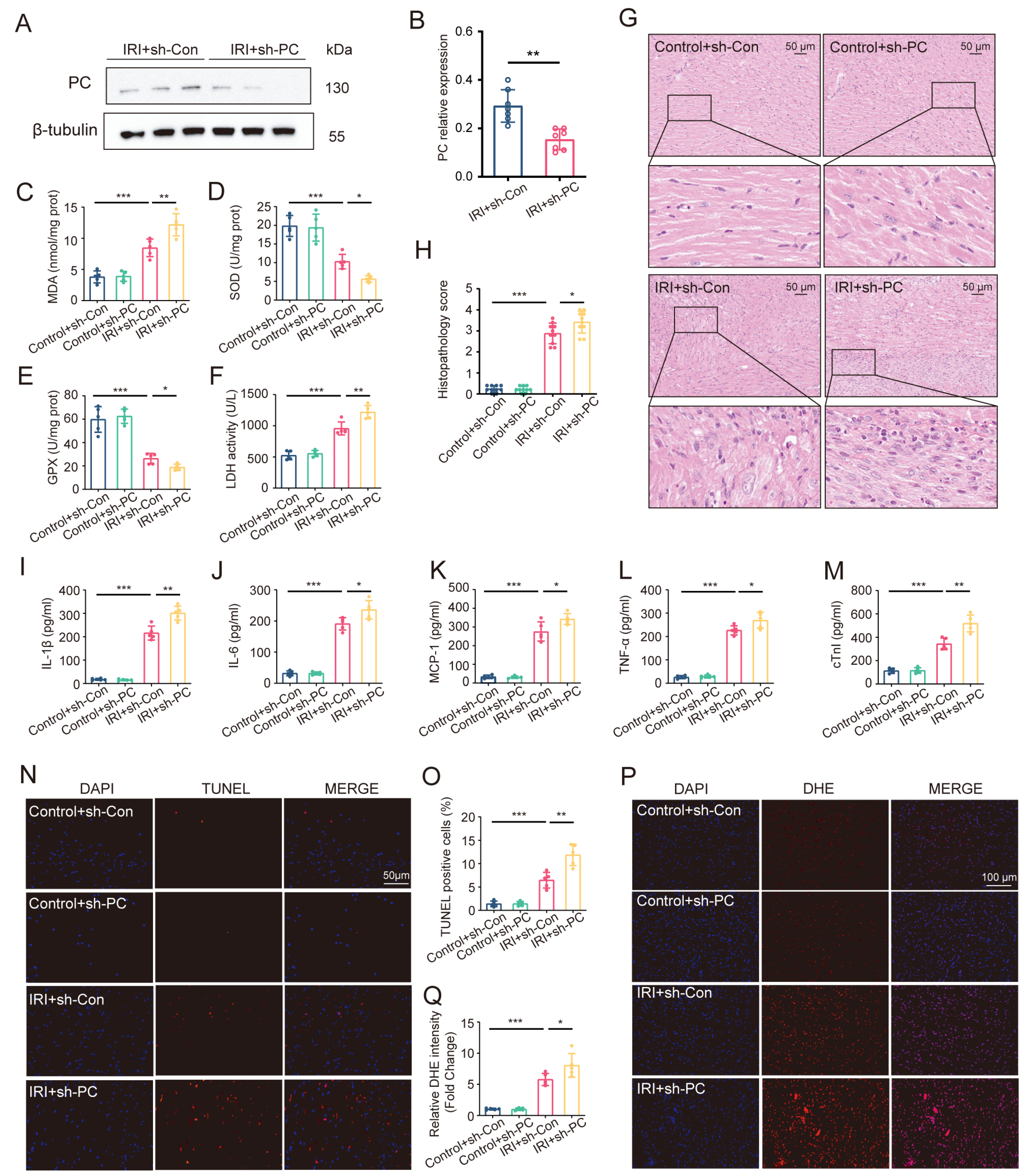
3.2. PC Knockdown Aggravates Myocardial IRI in Heart Transplantation
3.3. PC Knockdown Aggravates H/R Injury In Vitro
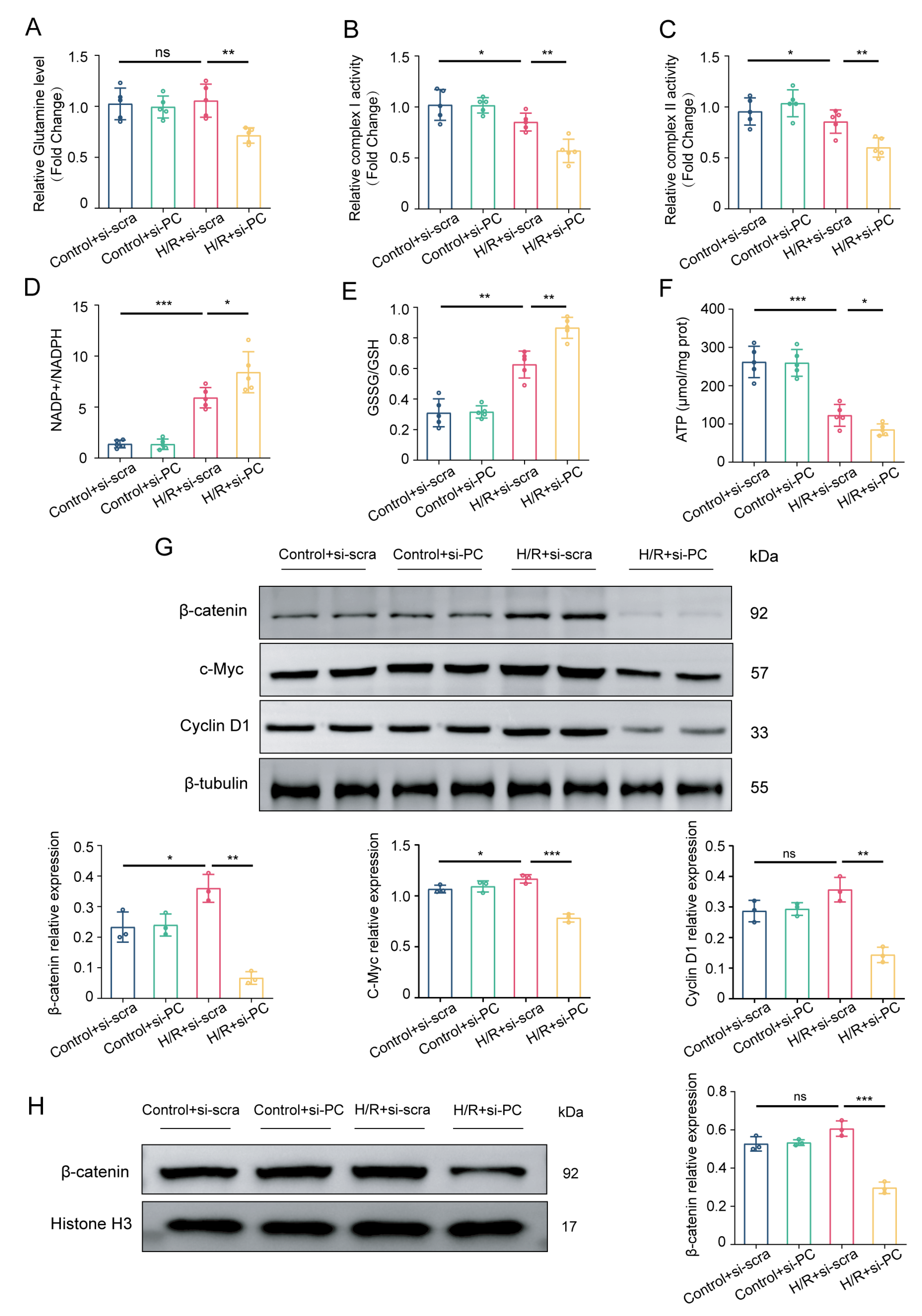
3.4. Knocking Down PC Suppresses the Wnt/β-Catenin Pathway and Glutamine Metabolism
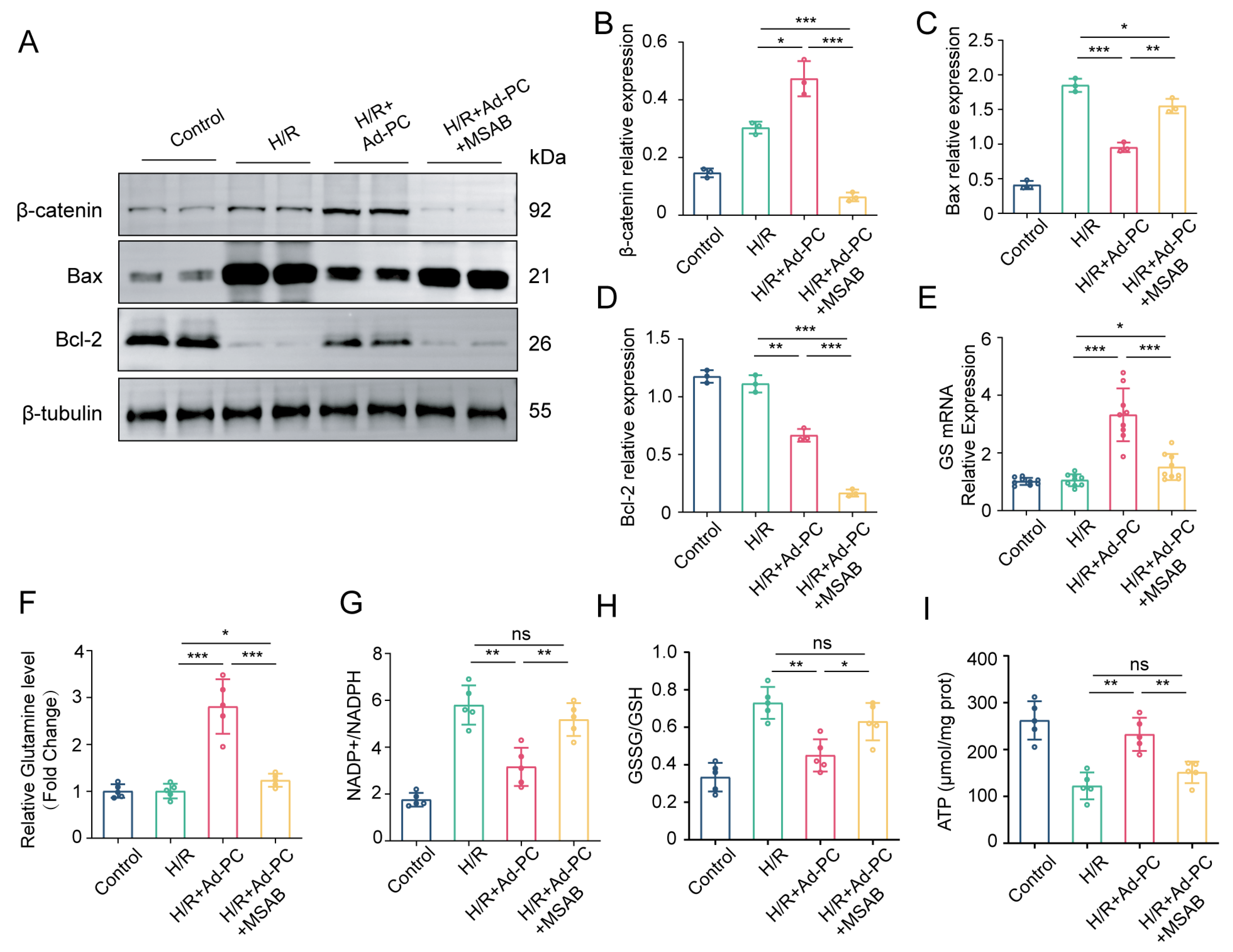
3.5. Wnt/β-Catenin Pathway Is Essential to Alleviate H/R Injury via PC Overexpression
3.6. PC Alleviates Myocardial IRI during Heart Transplantation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Capuzzimati, M.; Hough, O.; Liu, M. Cell death and ischemia-reperfusion injury in lung transplantation. J. Heart Lung Transpl. 2022, 41, 1003–1013. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.F.; Hosgood, S.A.; Nicholson, M.L. Ischemia-reperfusion injury in renal transplantation: 3 key signaling pathways in tubular epithelial cells. Kidney Int. 2019, 95, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.H.; Huckaby, L.V.; Sloan, B.; Pope, N.H.; Witer, L.J.; Tedford, R.J.; Houston, B.A.; Hashmi, Z.A.; Katz, M.R.; Kilic, A. Prolonged ischemia times for heart transplantation: Impact of the 2018 allocation change. Ann. Thorac. Surg. 2022, 114, 1386–1394. [Google Scholar] [CrossRef] [PubMed]
- Valero-Masa, M.J.; González-Vílchez, F.; Almenar-Bonet, L.; Crespo-Leiro, M.G.; Manito-Lorite, N.; Sobrino-Márquez, J.M.; Gómez-Bueno, M.; Delgado-Jiménez, J.F.; Pérez-Villa, F.; Brossa Loidi, V.; et al. Cold ischemia >4 hours increases heart transplantation mortality. An analysis of the Spanish heart transplantation registry. Int. J. Cardiol. 2020, 319, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; O’Brien, M.E.; Yu, J.; Xu, C.; Zhang, Q.; Lu, S.; Liang, L.; An, X.; McDyer, J.F.; Mallampalli, R.K. Prolonged cold ischemia induces necroptotic cell death in ischemia-reperfusion injury and contributes to primary graft dysfunction after lung transplantation. Am. J. Respir. Cell Mol. Biol. 2019, 61, 244–256. [Google Scholar] [CrossRef]
- Chambers, D.C.; Yusen, R.D.; Cherikh, W.S.; Goldfarb, S.B.; Kucheryavaya, A.Y.; Khusch, K.; Levvey, B.J.; Lund, L.H.; Meiser, B.; Rossano, J.W.; et al. The registry of the international society for heart and lung transplantation: Thirty-fourth adult lung and heart-lung transplantation report-2017; focus theme: Allograft ischemic time. J. Heart Lung Transpl. 2017, 36, 1047–1059. [Google Scholar] [CrossRef]
- Lin, Q.; He, Y.; Wang, X.; Zhang, Y.; Hu, M.; Guo, W.; He, Y.; Zhang, T.; Lai, L.; Sun, Z.; et al. Targeting Pyruvate Carboxylase by a Small Molecule Suppresses Breast Cancer Progression. Adv. Sci. 2020, 7, 1903483. [Google Scholar] [CrossRef]
- Kerr, A.G.; Wang, Z.; Wang, N.; Kwok, K.H.M.; Jalkanen, J.; Ludzki, A.; Lecoutre, S.; Langin, D.; Bergo, M.O.; Dahlman, I.; et al. The long noncoding RNA ADIPINT regulates human adipocyte metabolism via pyruvate carboxylase. Nat. Commun. 2022, 13, 2958. [Google Scholar] [CrossRef]
- Li, X.; Cheng, K.K.Y.; Liu, Z.; Yang, J.K.; Wang, B.; Jiang, X.; Zhou, Y.; Hallenborg, P.; Hoo, R.L.C.; Lam, K.S.L.; et al. The MDM2-p53-pyruvate carboxylase signalling axis couples mitochondrial metabolism to glucose-stimulated insulin secretion in pancreatic beta-cells. Nat. Commun. 2016, 7, 11740. [Google Scholar] [CrossRef]
- Cappel, D.A.; Deja, S.; Duarte, J.A.G.; Kucejova, B.; Iñigo, M.; Fletcher, J.A.; Fu, X.; Berglund, E.D.; Liu, T.; Elmquist, J.K.; et al. Pyruvate-carboxylase-mediated anaplerosis promotes antioxidant capacity by sustaining tca cycle and redox metabolism in liver. Cell Metab. 2019, 29, 1291–1305. [Google Scholar] [CrossRef]
- Wilmanski, T.; Zhou, X.; Zheng, W.; Shinde, A.; Donkin, S.S.; Wendt, M.; Burgess, J.R.; Teegarden, D. Inhibition of pyruvate carboxylase by 1α,25-dihydroxyvitamin D promotes oxidative stress in early breast cancer progression. Cancer Lett. 2017, 411, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Zhang, Y.; Li, W.; Chen, K.; Xiang, M.; Ma, H. Glutamine metabolism: From proliferating cells to cardiomyocytes. Metabolism 2021, 121, 154778. [Google Scholar] [CrossRef] [PubMed]
- Kennel, P.J.; Liao, X.; Saha, A.; Ji, R.; Zhang, X.; Castillero, E.; Brunjes, D.; Takayama, H.; Naka, Y.; Thomas, T.; et al. Impairment of myocardial glutamine homeostasis induced by suppression of the amino acid carrier SLC1A5 in failing myocardium. Circ.-Heart Fail. 2019, 12, e6336. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.H.; Zhang, X.J.; Shang, H.Q.; Wang, X.; Rong, D. Glutamine protects myocardial ischemia-reperfusion injury in rats through the PI3K/Akt signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 444–451. [Google Scholar] [PubMed]
- Kou, Y.; Zheng, W.T.; Zhang, Y.R. Inhibition of miR-23 protects myocardial function from ischemia-reperfusion injury through restoration of glutamine metabolism. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4286–4293. [Google Scholar] [PubMed]
- Liu, S.; Yang, Y.; Song, Y.Q.; Geng, J.; Chen, Q.L. Protective effects of N(2)-L-alanyl-L-glutamine mediated by the JAK2/STAT3 signaling pathway on myocardial ischemia reperfusion. Mol. Med. Rep. 2018, 17, 5102–5108. [Google Scholar] [CrossRef] [PubMed]
- Fu, A.; van Rooyen, L.; Evans, L.; Armstrong, N.; Avizonis, D.; Kin, T.; Bird, G.H.; Reddy, A.; Chouchani, E.T.; Liesa-Roig, M.; et al. Glucose metabolism and pyruvate carboxylase enhance glutathione synthesis and restrict oxidative stress in pancreatic islets. Cell Rep. 2021, 37, 110037. [Google Scholar] [CrossRef] [PubMed]
- Loganathan, S.; Korkmaz-Icöz, S.; Radovits, T.; Li, S.; Mikles, B.; Barnucz, E.; Hirschberg, K.; Karck, M.; Szabó, G. Effects of soluble guanylate cyclase activation on heart transplantation in a rat model. J. Heart Lung Transpl. 2015, 34, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Su, Y.; Zhu, C.; Quan, D.; Skaro, A.I.; McAlister, V.; Lacefield, J.C.; Jiang, J.; Xue, P.; Wang, Y.; et al. An addition of U0126 protecting heart grafts from prolonged cold ischemia-reperfusion injury in heart transplantation: A new preservation strategy. Transplantation 2021, 105, 308–317. [Google Scholar] [CrossRef]
- Zhu, C.; Su, Y.; Juriasingani, S.; Zheng, H.; Veramkovich, V.; Jiang, J.; Sener, A.; Whiteman, M.; Lacefield, J.; Nagpal, D.; et al. Supplementing preservation solution with mitochondria-targeted H2 S donor AP39 protects cardiac grafts from prolonged cold ischemia-reperfusion injury in heart transplantation. Am. J. Transplant. 2019, 19, 3139–3148. [Google Scholar] [CrossRef]
- Adebayo Michael, A.O.; Ko, S.; Tao, J.; Moghe, A.; Yang, H.; Xu, M.; Russell, J.O.; Pradhan-Sundd, T.; Liu, S.; Singh, S.; et al. Inhibiting glutamine-dependent mTORC1 activation ameliorates liver cancers driven by β-catenin mutations. Cell Metab. 2019, 29, 1135–1150. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, C.; Pan, Y.; Zhou, J.; Ju, H.; Zhang, Y. Pyruvate carboxylase promotes malignant transformation of papillary thyroid carcinoma and reduces iodine uptake. Cell Death Discov. 2022, 8, 423. [Google Scholar] [CrossRef] [PubMed]
- Kiesel, V.A.; Sheeley, M.P.; Coleman, M.F.; Cotul, E.K.; Donkin, S.S.; Hursting, S.D.; Wendt, M.K.; Teegarden, D. Pyruvate carboxylase and cancer progression. Cancer Metab. 2021, 9, 20. [Google Scholar] [CrossRef]
- Pereira, R.M.; Rodrigues, K.C.D.C.; Sant’Ana, M.R.; Peruca, G.F.; Morelli, A.P.; Simabuco, F.M.; da Silva, A.S.R.; Cintra, D.E.; Ropelle, E.R.; Pauli, J.R.; et al. Strength exercise reduces hepatic pyruvate carboxylase and gluconeogenesis in DIO mice. J. Endocrinol. 2020, 247, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Lao-On, U.; Attwood, P.V.; Jitrapakdee, S. Roles of pyruvate carboxylase in human diseases: From diabetes to cancers and infection. J. Mol. Med. 2018, 96, 237–247. [Google Scholar] [CrossRef]
- Cao, Z.; Zhou, Y.; Zhu, S.; Feng, J.; Chen, X.; Liu, S.; Peng, N.; Yang, X.; Xu, G.; Zhu, Y. Pyruvate carboxylase activates the RIG-I-like receptor-mediated antiviral immune response by targeting the MAVS signalosome. Sci. Rep. 2016, 6, 22002. [Google Scholar] [CrossRef]
- Yim, S.A.; Lim, Y.S.; Kim, J.W.; Hwang, S.B. Nonstructural 5A protein of hepatitis C virus interacts with pyruvate carboxylase and modulates viral propagation. PLoS ONE 2013, 8, e68170. [Google Scholar] [CrossRef]
- Durante, W. The emerging role of l-glutamine in cardiovascular health and disease. Nutrients 2019, 11, 2092. [Google Scholar] [CrossRef] [PubMed]
- Tewari, D.; Bawari, S.; Sharma, S.; DeLiberto, L.K.; Bishayee, A. Targeting the crosstalk between canonical Wnt/β-catenin and inflammatory signaling cascades: A novel strategy for cancer prevention and therapy. Pharmacol. Therapeut 2021, 227, 107876. [Google Scholar] [CrossRef]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/β-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef]
- Sassi, Y.; Avramopoulos, P.; Ramanujam, D.; Grüter, L.; Werfel, S.; Giosele, S.; Brunner, A.D.; Esfandyari, D.; Papadopoulou, A.S.; De Strooper, B.; et al. Cardiac myocyte miR-29 promotes pathological remodeling of the heart by activating Wnt signaling. Nat. Commun. 2017, 8, 1614. [Google Scholar] [CrossRef] [PubMed]
- Sierra-Pagan, J.E.; Dsouza, N.; Das, S.; Larson, T.A.; Sorensen, J.R.; Ma, X.; Stan, P.; Wanberg, E.J.; Shi, X.; Garry, M.G.; et al. FOXK1 regulates Wnt signalling to promote cardiogenesis. Cardiovasc. Res. 2023, 119, 1728–1739. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Wei, W. Therapeutic effect of a novel Wnt pathway inhibitor on cardiac regeneration after myocardial infarction. J. Am. Coll. Cardiol. 2018, 72, C69. [Google Scholar] [CrossRef]
- Buikema, J.W.; Mady, A.S.; Mittal, N.V.; Atmanli, A.; Caron, L.; Doevendans, P.A.; Sluijter, J.P.; Domian, I.J. Wnt/beta-catenin signaling directs the regional expansion of first and second heart field-derived ventricular cardiomyocytes. Development 2013, 140, 4165–4176. [Google Scholar] [CrossRef] [PubMed]
- Meyer, I.S.; Li, X.; Meyer, C.; Voloshanenko, O.; Pohl, S.; Boutros, M.; Katus, H.A.; Frey, N.; Leuschner, F. Blockade of Wnt secretion attenuates myocardial ischemia-reperfusion injury by modulating the inflammatory response. Int. J. Mol. Sci. 2022, 23, 12252. [Google Scholar] [CrossRef] [PubMed]
- Moskowitzova, K.; Shin, B.; Liu, K.; Ramirez-Barbieri, G.; Guariento, A.; Blitzer, D.; Thedsanamoorthy, J.K.; Yao, R.; Snay, E.R.; Inkster, J.A.H.; et al. Mitochondrial transplantation prolongs cold ischemia time in murine heart transplantation. J. Heart Lung Transpl. 2019, 38, 92–99. [Google Scholar] [CrossRef]
- Cai, S.; Ichimaru, N.; Zhao, M.; Fujino, M.; Ito, H.; Ota, U.; Nakajima, M.; Tanaka, T.; Nonomura, N.; Li, X.K.; et al. Prolonged mouse cardiac graft cold storage via attenuating ischemia-reperfusion injury using a new antioxidant-based preservation solution. Transplantation 2016, 100, 1032–1040. [Google Scholar] [CrossRef]
- Benke, K.; Jász, D.K.; Szilágyi, Á.L.; Baráth, B.; Tuboly, E.; Márton, A.R.; Varga, P.; Mohácsi, Á.; Szabó, A.; Széll, Z.; et al. Methane supplementation improves graft function in experimental heart transplantation. J. Heart Lung Transpl. 2021, 40, 183–192. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Lan, H.; Wang, Y.; Zheng, Q.; Li, C.; Wang, K.; Xiong, T.; Wu, Q.; Dong, N. Pyruvate Carboxylase Attenuates Myocardial Ischemia–Reperfusion Injury in Heart Transplantation via Wnt/β-Catenin-Mediated Glutamine Metabolism. Biomedicines 2024, 12, 1826. https://doi.org/10.3390/biomedicines12081826
Wang Z, Lan H, Wang Y, Zheng Q, Li C, Wang K, Xiong T, Wu Q, Dong N. Pyruvate Carboxylase Attenuates Myocardial Ischemia–Reperfusion Injury in Heart Transplantation via Wnt/β-Catenin-Mediated Glutamine Metabolism. Biomedicines. 2024; 12(8):1826. https://doi.org/10.3390/biomedicines12081826
Chicago/Turabian StyleWang, Zihao, Hongwen Lan, Yixuan Wang, Qiang Zheng, Chenghao Li, Kan Wang, Tixiusi Xiong, Qingping Wu, and Nianguo Dong. 2024. "Pyruvate Carboxylase Attenuates Myocardial Ischemia–Reperfusion Injury in Heart Transplantation via Wnt/β-Catenin-Mediated Glutamine Metabolism" Biomedicines 12, no. 8: 1826. https://doi.org/10.3390/biomedicines12081826