
Cortisol Dose-Dependently Impairs Migration and Tube-like Formation in a Trophoblast Cell Line and Modulates Inflammatory and Angiogenic Genes

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
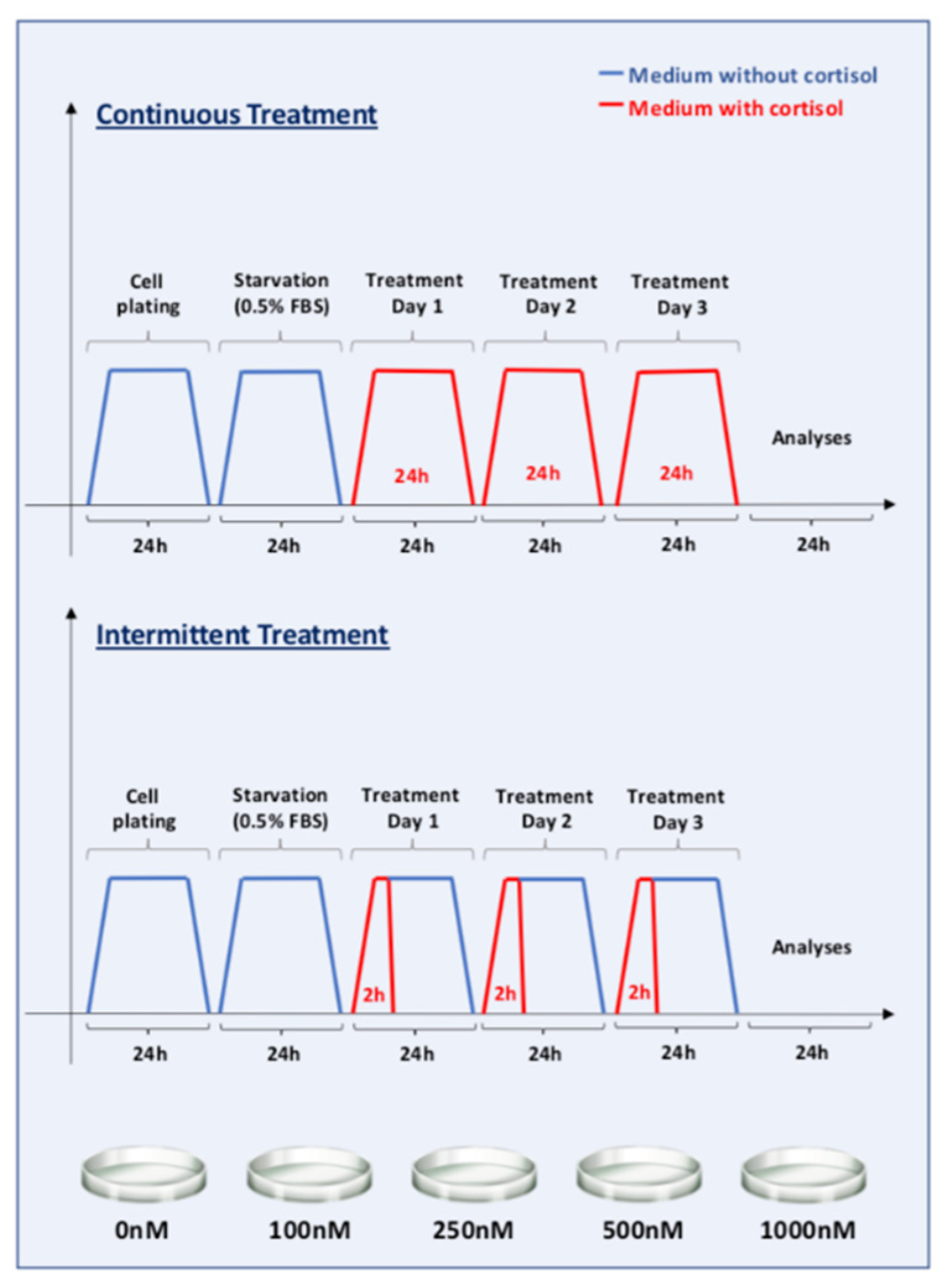
2.1. Cell Culture and Cortisol Treatment
2.2. RNA Extraction and RT-qPCR
2.3. MTT—Cell Proliferation Assays
2.4. Wound Healing Assay
2.5. Tube-Like Formation Assay
2.6. Statistical Analyses
3. Results
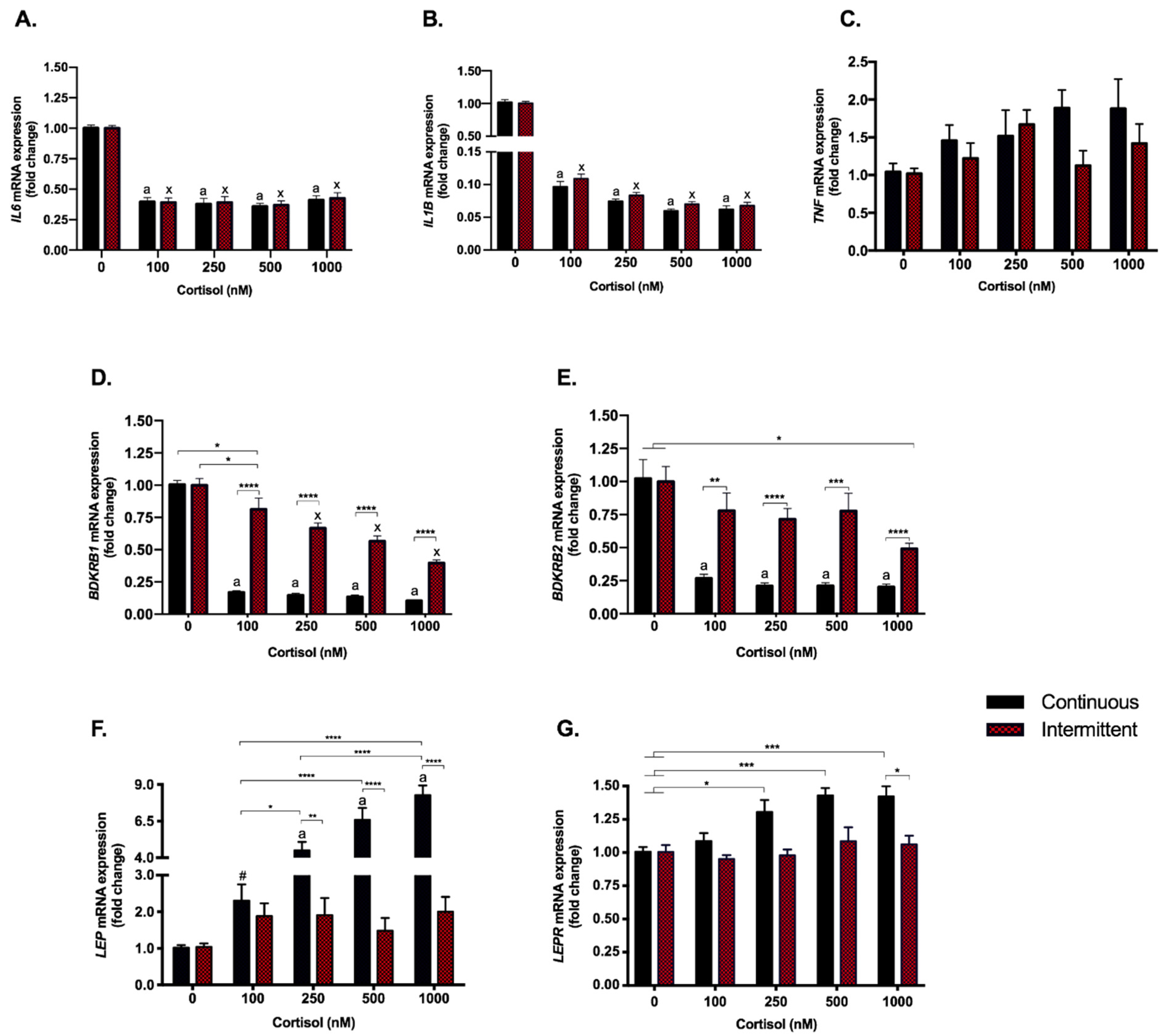
3.1. Continuous Treatment Represses NR3C1 Gene without Modulations in HSD11B2
3.2. The Anti-Inflammatory Effect of Cortisol Was Found to Be Similar with Both Treatments; However, Kinin Receptors Responded Differently to Each Exposure
3.3. Continuous Treatment Modulates Leptin (LEP) and Leptin Receptor (LEPR) Expression
3.4. Angiogenesis Pathway Genes Are Modulated by Continuous Exposure to Cortisol Even at Low Doses. PlGF and KDR (VEGFR-2) Are Also Repressed by the Highest Dose in the Intermittent Exposure
3.5. Cortisol Treatment Represses the Expression of Chemokine CXCL12 and Its Receptor
3.6. Cortisol Had No Impact on the Viability of the SGHPL-4 Cell Line
3.7. Cortisol Affects Cell Migration in a Dose-Dependent Manner
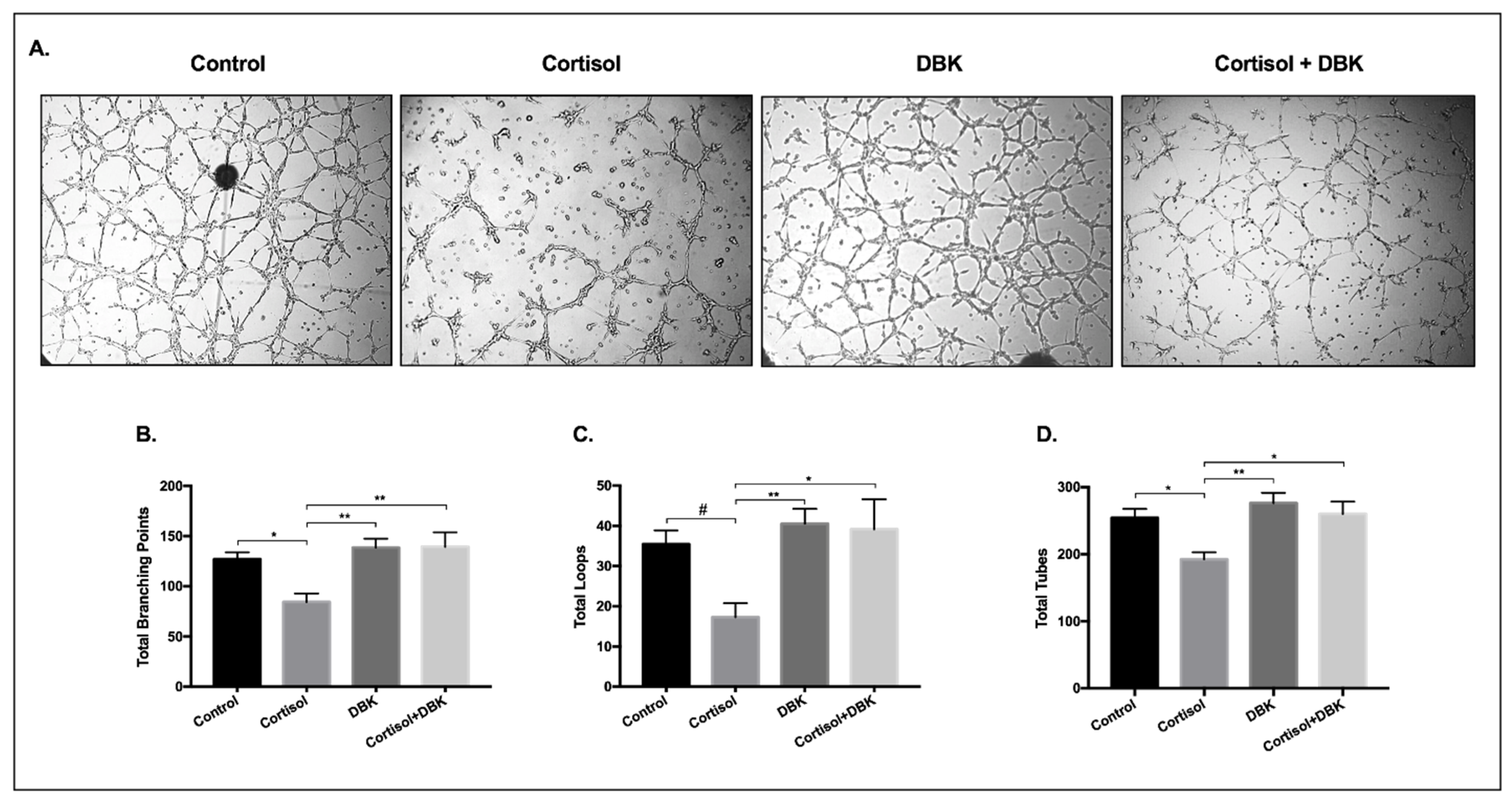
3.8. Cortisol Treatment Reduces the Formation of Tube-Like Structures and DBK Modulates This Response
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers |
|---|---|
| NR3C1 | For: 5′-CGA CCA ATG TAA ACA CAT GCT-3 Rev: 5′-CCG TCC TTA GGA ACT GAA GAG-3′ |
| HSD11B2 | For: 5′-TCA AGA CAG AGT CAG TGA GAA ACG-3′ Rev: 5′-GGA ACT GCC CAT GCA AGT G-3′ |
| 18S | For: 5′-ACA CGT TCC ACC TCA TCC TC-3′ Rev: 5′-CTT TGC CAT CAC TGC CAT TA-3′ |
| IL6 | For: 5′-TGG CTG AAA AAG ATG GAT GCT-3′ Rev: 5′-TCT GCA CAG CTC TGG CTT GT-3′ |
| IL1B | For: 5′-AGG GAC AGG ATA TGG AGC AAC AAG -3′ Rev: 5′-CAT CTT TCA ACA CGC AGG ACA GGT-3′ |
| TNF | For: 5′-TGG CCC AGG CAG TCA GA-3′ Rev: 5′-GGT TTG CTA CAA CAT GG GCT ACA-3′ |
| BDKRB1 (B1R) | For: 5′-CTG CAC AGA GTG CTG CCA ACA TT-3′ Rev: 5′-ACA CCA GAT CAG AGG CTG CCA GG-3′ |
| BDKRB2 (B2R) | For: 5′-TCT GGC TTC TGG GCT CCG AG-3′ Rev: 5′-AGC GGC ATG GGC ACT TCA GT-3′ |
| VEGFA | For: 5′-TACCTCCACCATGCCAAGTG -3′ Rev: 5′-GATGATTCTGCCCTCCTCCTT -3′ |
| PlGF | For: 5′-TGC TGC GGC GAT GAG AAT C-3′ Rev: 5′-CCC TTG GGT CTC CTC CTT TC-3′ |
| KDR | For: 5′-GCA TCT CAT CTG TTA CAG C-3′ Rev: 5′-CTT CAT CAA TCT TTA CCC C-3′ |
| sFLT1 | For: 5′-AAT CAG AGG TGA GCA CTG CAA C-3′ Rev: 5′-TGG TAC AAT CAT TCC TTG TGC TTT-3′ |
| MMP2 | For: 5′-CCC ACT GCG GTT TTC TCG AAT-3′ Rev: 5′-CAA AGG GGT ATC CAT CGC CAT-3′ |
| MMP9 | For: 5′-AGA CCT GGG CAG ATT CCA AAC-3′ Rev: 5′-CGG CAA GTC TTC CGA GTA GT-3′ |
| LEP | For: 5′-CAT TGG GGA ACC CTG TGC GGA TTC-3′ Rev: 5′-TGG CAG CTC TTA GAG AAG GCC AGC-3′ |
| LEPR | For: 5′-GTA AGA GGC TAG ATG GAC TGG GAT AT-3′ Rev: 5′-ATT CTC CAA AAT TCA GGT CCT CTC A-3′ |
| CXCL12 | For: 5′-ATT CTC AAC ACT CCA AAC TGT GC-3′ Rev: 5′-ACT TTA GCT TCG GGT CAA TGC-3′ |
| CXCR4 | For: 5′-GGG CAA TGG ATT GGT CAT CCT-3′ Rev: 5′-TGC AGC CTG TAC TTG TCC G-3′ |
| TGFB1 | For: 5′-ACC AAC TAT TGC TTC AGC TC-3′ Rev: 5′-TTA TGC TGG TTG TAC AGG G-3′ |
| E-cadherin | For: 5′-GCC ACA TAC ACT CTC TTC TC-3′ Rev: 5′-CCA TCA CAG AGG TTC CTG-3′ |



References
- Aufdenblatten, M.; Baumann, M.; Raio, L.; Dick, B.; Frey, B.M.; Schneider, H.; Surbek, D.; Hocher, B.; Mohaupt, M.G. Prematurity is related to high placental cortisol in preeclampsia. Pediatric Res. 2009, 65, 198–202. [Google Scholar] [CrossRef] [Green Version]
- Field, T.; Diego, M. Cortisol: The culprit prenatal stress variable. Int. J. Neurosci. 2008, 118, 1181. [Google Scholar] [CrossRef] [PubMed]
- Castellucci, M.; Schepe, M.; Scheffen, I.; Celona, A.; Kaufmann, P. The development of the human placental villous tree. Anat. Embryol. 1990, 181, 117–128. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E.; Charnock-Jones, D.S. The influence of the intrauterine environment on human placental development. Int. J. Dev. Biol. 2010, 54, 303–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pijnenborg, R.; Bland, J.M.; Robertson, W.B.; Brosens, I. Uteroplacental arterial changes related to interstitial trophoblast migration in early human pregnancy. Placenta 1983, 4, 397–413. [Google Scholar] [CrossRef]
- Turco, M.Y.; Gardner, L.; Kay, R.G.; Hamilton, R.S.; Prater, M.; Hollinshead, M.S.; McWhinnie, A.; Esposito, L.; Fernando, R.; Skelton, H.; et al. Trophoblast organoids as a model for maternal–fetal interactions during human placentation. Nature 2018, 564, 263–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeldovich, V.B.; Robbins, J.R.; Kapidzic, M.; Lauer, P.; Bakardjiev, A.I. Invasive extravillous trophoblasts restrict intracellular growth and spread of Listeria monocytogenes. PLoS Pathog. 2011, 7, e1002005. [Google Scholar] [CrossRef] [Green Version]
- Meekins, J.W.; Pijnenborg, R.; Hanssens, M.; MCFadyen, I.R.; van Asshe, A. A study of placental bed spiral arteries and trophoblast invasion in normal and severe pre-eclamptic pregnancies. BJOG Int. J. Obstet. Gynaecol. 1994, 101, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Ness, R.B.; Sibai, B.M. Shared and disparate components of the pathophysiologies of fetal growth restriction and preeclampsia. Am. J. Obstet. Gynecol. 2006, 195, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Broughton Pipkin, F.; Rubin, P.C. Pre-eclampsia—The ‘disease of theories’. Br. Med. Bull. 1994, 50, 381–396. [Google Scholar] [CrossRef]
- Choy, M.Y.; Manyonda, I.T. The phagocytic activity of human first trimester extravillous trophoblast. Hum. Reprod. 1998, 13, 2941–2949. [Google Scholar] [CrossRef] [Green Version]
- Choy, M.Y.; St Whitley, G.; Manyonda, I.T. Efficient, rapid and reliable establishment of human trophoblast cell lines using poly-L-ornithine. Early Pregnancy 2000, 4, 124–143. [Google Scholar]
- McCormick, J.; Whitley, G.S.J.; Le Bouteiller, P.; Cartwright, J.E. Soluble HLA-G regulates motility and invasion of the trophoblast-derived cell line SGHPL-4. Hum. Reprod. 2009, 24, 1339–1345. [Google Scholar] [CrossRef] [Green Version]
- Lasha, G.E.; Cartwright, J.E.; Whitley, G.S.J.; Trew, A.J.; Baker, P.N. The effects of angiogenic growth factors on extravillous trophoblast invasion and motility. Placenta 2000, 21, 593–594. [Google Scholar] [CrossRef]
- Angelova, M.; Machado, H.; Swan, K.; Morris, C.; Sullivan, D. Extravillous trophoblast migration and invasion assay. Bio-Protocol 2013, 3, 3–6. [Google Scholar] [CrossRef] [Green Version]
- Westwood, M.; Al-Saghir, K.; Finn-Sell, S.; Tan, C.; Cowley, E.; Berneau, S.; Adlam, D.; Johnstone, E.D. Vitamin D attenuates sphingosine-1-phosphate (S1P)-mediated inhibition of extravillous trophoblast migration. Placenta 2017, 60, 1–8. [Google Scholar] [CrossRef]
- Harris, L.K.; Clancy, O.H.; Myers, J.E.; Baker, P.N. Plasma from women with preeclampsia inhibits trophoblast invasion. Reprod. Sci. 2009, 16, 1082–1090. [Google Scholar] [CrossRef] [PubMed]
- Shiverick, K.T.; King, A.; Frank, H.G.; Whitley, G.S.J.; Cartwright, J.E.; Schneider, H. Cell culture models of human trophoblast II: Trophoblast cell lines—A workshop report. Placenta 2001, 22, 104–106. [Google Scholar] [CrossRef] [PubMed]
- Challis, J.R.G.; Sloboda, D.; Matthews, S.G.; Holloway, A.; Alfaidy, N.; Patel, F.A.; Whittle, W.; Fraser, M.; Moss, T.J.M.; Newnham, J. The fetal placental hypothalamic-pituitary-adrenal (HPA) axis, parturition and post natal health. Mol. Cell. Endocrinol. 2001, 185, 135–144. [Google Scholar] [CrossRef]
- McEwen, B.S. The neurobiology of stress: From serendipity to clinical relevance. Brain Res. 2000, 886, 172–189. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.Y.; Kim, E.; Choi, M.H. Technical and clinical aspects of cortisol as a biochemical marker of chronic stress. BMB Rep. 2015, 48, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Amdi, C.; Giblin, L.; Hennessy, A.A.; Ryan, T.; Stanton, C.; Stickland, N.C.; Lawlor, P.G. Feed allowance and maternal backfat levels during gestation influence maternal cortisol levels, milk fat composition and offspring growth. J. Nutr. Sci. 2013, 2, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, E.E.; Zack, E.; Battaglini, C.; Viru, M.; Viru, A.; Hackney, A.C. Exercise and circulating Cortisol levels: The intensity threshold effect. J. Endocrinol. Investig. 2008, 31, 587–591. [Google Scholar] [CrossRef]
- Michael, A.E.; Papageorghiou, A.T. Potential significance of physiological and pharmacological glucocorticoids in early pregnancy. Hum. Reprod. Update 2008, 14, 497–517. [Google Scholar] [CrossRef] [Green Version]
- Moreau, P.; Faure, O.; Lefebvre, S.; Ibrahim, E.G.; O’Brien, M.; Gourand, L.; Dausset, J.; Carosella, E.D.; Paul, P. Glucocorticoid hormones upregulate levels of HLA-G transcripts in trophoblasts. Transplant. Proc. 2001, 33, 2277–2280. [Google Scholar] [CrossRef]
- Hompes, T.; Vrieze, E.; Fieuws, S.; Simons, A.; Jaspers, L.; van Bussel, J.; Schops, G.; Gellens, E.; van Bree, R.; Verhaeghe, J.; et al. The influence of maternal cortisol and emotional state during pregnancy on fetal intrauterine growth. Pediatric Res. 2012, 72, 305–315. [Google Scholar] [CrossRef]
- Hansen, D.; Lou, H.C.; Olsen, J. Serious life events and congenital malformations: A national study with complete follow-up. Lancet 2000, 356, 875–880. [Google Scholar] [CrossRef]
- Obel, C.; Hedegaard, M.; Henriksen, T.B.; Secher, N.J.; Olsen, J.; Levine, S. Stress and salivary cortisol during pregnancy. Psychoneuroendocrinology 2005, 30, 647–656. [Google Scholar] [CrossRef]
- French, N.P.; Hagan, R.; Evans, S.F.; Godfrey, M.; Newnham, J.P. Repeated antenatal corticosteroids: Size at birth and subsequent development. Am. J. Obstet. Gynecol. 1999, 180, 114–121. [Google Scholar] [CrossRef]
- Gennari-Moser, C.; Khankin, E.V.; Schüller, S.; Escher, G.; Frey, B.M.; Portmann, C.B.; Baumann, M.U.; Lehmann, A.D.; Surbek, D.; Karumanchi, S.A.; et al. Regulation of placental growth by aldosterone and cortisol. Endocrinology 2011, 152, 263–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.; Witte, E.; McGee, D.; Knott, J.; Narang, K.; Racicot, K. Cortisol inhibits CSF2 and CSF3 via DNA methylation and inhibits invasion in first-trimester trophoblast cells. Am. J. Reprod. Immunol. 2017, 78, e12741. [Google Scholar] [CrossRef]
- Kisanga, E.P.; Tang, Z.; Guller, S.; Whirledge, S. Glucocorticoid signaling regulates cell invasion and migration in the human first-trimester trophoblast cell line Sw.71. Am. J. Reprod. Immunol. 2018, 80, e12974. [Google Scholar] [CrossRef]
- Funder, J.; Pearce, P.; Smith, R.; Smith, A. Mineralocorticoid action: Target tissue specificity is enzyme, not receptor, mediated. Science 1988, 242, 583–585. [Google Scholar] [CrossRef] [PubMed]
- McCalla, C.O.; Nacharaju, V.L.; Muneyyirci-Delale, O.; Glasgow, S.; Feldman, J.G. Placental 11β-hydroxysteroid dehydrogenase activity in normotensive and pre-eclamptic pregnancies. Steroids 1998, 63, 511–515. [Google Scholar] [CrossRef]
- Alfaidy, N.; Blot-Chabaud, M.; Robic, D.; Kenouch, S.; Bourbouze, R.; Bonvalet, J.P.; Farman, N. Characteristics and regulation of 11β-hydroxysteroid dehydrogenase of proximal and distal nephron. BBA Gen. Subj. 1995, 1243, 461–468. [Google Scholar] [CrossRef]
- Alfaidy, N.; Gupta, S.; DeMarco, C.; Caniggia, I.; Challis, J.R.G. Oxygen regulation of placental 11β-hydroxysteroid dehydrogenase 2: Physiological and pathological implications. J. Clin. Endocrinol. Metab. 2002, 87, 4797–4805. [Google Scholar] [CrossRef] [Green Version]
- Hanna, J.; Goldman-Wohl, D.; Hamani, Y.; Avraham, I.; Greenfield, C.; Natanson-Yaron, S.; Prus, D.; Cohen-Daniel, L.; Arnon, T.I.; Manaster, I.; et al. Decidual NK cells regulate key developmental processes at the human fetal-maternal interface. Nat. Med. 2006, 12, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Pollheimer, J.; Vondra, S.; Baltayeva, J.; Beristain, A.G.; Knöfler, M. Regulation of placental extravillous trophoblasts by the maternal uterine environment. Front. Immunol. 2018, 9, 2597. [Google Scholar] [CrossRef]
- Doherty, T.M.; Hu, A.; Salik, I. Physiology, Neonatal; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Yang, Q.; Wang, W.; Liu, C.; Wang, Y.; Sun, K. Compartmentalized localization of 11β-HSD 1 and 2 at the feto-maternal interface in the first trimester of human pregnancy. Placenta 2016, 46, 63–71. [Google Scholar] [CrossRef]
- Lee, M.J.; Wang, Z.; Yee, H.; Ma, Y.; Swenson, N.; Yang, L.; Kadner, S.S.; Baergen, R.N.; Logan, S.K.; Garabedian, M.J.; et al. Expression and regulation of glucocorticoid receptor in human placental villous fibroblasts. Endocrinology 2005, 146, 4619–4626. [Google Scholar] [CrossRef] [Green Version]
- Le Bellego, F.; Vaillancourt, C.; Lafond, J. Isolation and culture of term human cytotrophoblast cells and in vitro methods for studying human cytotrophoblast cells’ calcium uptake. Methods Mol. Biol. 2009, 550, 73–87. [Google Scholar] [CrossRef]
- Grada, A.; Otero-Vinas, M.; Prieto-Castrillo, F.; Obagi, Z.; Falanga, V. Research techniques made simple: Analysis of collective cell migration using the wound healing assay. J. Investig. Dermatol. 2017, 137, e11–e16. [Google Scholar] [CrossRef] [Green Version]
- Buckley, R.J.; Whitley, G.S.; Dumitriu, I.E.; Cartwright, J.E. Macrophage polarisation affects their regulation of trophoblast behaviour. Placenta 2016, 47, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Valdés, G.; Acuña, S.; Munizaga, A.; Soto, G.X.; Figueroa, C.D. Utero-placental cellular and nuclear expression of bradykinin B2 receptors in normal and preeclamptic pregnancies. Pregnancy Hypertens. 2016, 6, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Erices, R.; Corthorn, J.; Lisboa, F.; Valdés, G. Bradykinin promotes migration and invasion of human immortalized trophoblasts. Reprod. Biol. Endocrinol. 2011, 9, 97. [Google Scholar] [CrossRef] [Green Version]
- De Bosscher, K.; Vanden Berghe, W.; Haegeman, G. Mechanisms of anti-inflammatory action and of immunosuppression by glucocorticoids: Negative interference of activated glucocorticoid receptor with transcription factors. J. Neuroimmunol. 2000, 109, 16–22. [Google Scholar] [CrossRef]
- Wiley, J.W.; Higgins, G.A.; Athey, B.D. Stress and glucocorticoid receptor transcriptional programming in time and space: Implications for the brain-gut axis. Neurogastroenterol. Motil. 2016, 28, 12–25. [Google Scholar] [CrossRef]
- Diederich, S.; Quinkler, M.; Burkhardt, P.; Großmann, C.; Bähr, V.; Oelkers, W. 11β-Hydroxysteroid-dehydrogenase isoforms: Tissue distribution and implications for clinical medicine. Eur. J. Clin. Investig. 2000, 30 (Suppl. 3), 21–27. [Google Scholar] [CrossRef]
- Hayashi, R.; Wada, H.; Ito, K.; Adcock, I.M. Effects of glucocorticoids on gene transcription. Eur. J. Pharmacol. 2004, 500, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Koelsch, S.; Boehlig, A.; Hohenadel, M.; Nitsche, I.; Bauer, K.; Sack, U. The impact of acute stress on hormones and cytokines and how their recovery is affected by music-evoked positive mood. Sci. Rep. 2016, 6, 23008. [Google Scholar] [CrossRef] [Green Version]
- King, E.M.; Chivers, J.E.; Rider, C.F.; Minnich, A.; Giembycz, M.A.; Newton, R. Glucocorticoid repression of inflammatory gene expression shows differential responsiveness by transactivation- and transrepression-dependent mechanisms. PLoS ONE 2013, 8, e53936. [Google Scholar] [CrossRef] [PubMed]
- Seshagiri, P.B.; Vani, V.; Madhulika, P. Cytokines and blastocyst hatching. Am. J. Reprod. Immunol. 2016, 75, 208–217. [Google Scholar] [CrossRef] [Green Version]
- Bhoola, K.D.; Figueroa, C.D.; Worthy, K. Bioregulation of kinins: Kallikreins, kininogens, and kininases. Pharmacol. Rev. 1992, 44, 1–80. [Google Scholar]
- Marceau, F.; Hess, J.F.; Bachvarov, D.R. The B1 receptors for kinins. Pharmacol. Rev. 1998, 50, 357–386. [Google Scholar]
- Ikeda, Y.; Hayashi, I.; Kamoshita, E.; Yamazaki, A.; Endo, H.; Ishihara, K.; Yamashina, S.; Tsutsumi, Y.; Matsubara, H.; Majima, M. Host stromal bradykinin B2 receptor signaling facilitates tumor-associated angiogenesis and tumor growth. Cancer Res. 2004, 64, 5178–5185. [Google Scholar] [CrossRef] [Green Version]
- Hoegh, A.M.; Borup, R.; Nielsen, F.C.; Sørensen, S.; Hviid, T.V.F. Gene expression profiling of placentas affected by pre-eclampsia. J. Biomed. Biotechnol. 2010, 2010, 787545. [Google Scholar] [CrossRef]
- Yao, Y.Y.; Yin, H.; Shen, B.; Smith, R.S.; Liu, Y.; Gao, L.; Chao, L.; Chao, J. Tissue kallikrein promotes neovascularization and improves cardiac function by the Akt-glycogen synthase kinase-3β pathway. Cardiovasc. Res. 2008, 80, 354–364. [Google Scholar] [CrossRef] [Green Version]
- Thuringer, D.; Maulon, L.; Frelin, C. Rapid transactivation of the vascular endothelial growth factor receptor KDR/Flk-1 by the bradykinin B2 receptor contributes to endothelial nitric-oxide synthase activation in cardiac capillary endothelial cells. J. Biol. Chem. 2002, 277, 2028–2032. [Google Scholar] [CrossRef] [Green Version]
- Bader, M. Kallikrein-Kinin system in neovascularization. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 617–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzschoppe, A.; Fahlbusch, F.; Seidel, J.; Dörr, H.G.; Rascher, W.; Goecke, T.W.; Beckmann, M.W.; Schild, R.L.; Struwe, E.; Dötsch, J. Dexamethasone stimulates the expression of leptin and 11β-HSD2 in primary human placental trophoblastic cells. Eur. J. Obstet. Gynecol. Reprod. Biol. 2011, 156, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Masuzaki, H.; Ogawa, Y.; Sagawa, N.; Hosoda, K.; Matsumoto, T.; Mise, H.; Nishimura, H.; Yoshimasa, Y.; Tanaka, I.; Mori, T.; et al. Nonadipose tissue production of leptin: Leptin as a novel placenta-derived hormone in humans. Nat. Med. 1997, 3, 1029–1033. [Google Scholar] [CrossRef]
- Basak, S.; Duttaroy, A.K. Leptin induces tube formation in first-trimester extravillous trophoblast cells. Eur. J. Obstet. Gynecol. Reprod. Biol. 2012, 164, 24–29. [Google Scholar] [CrossRef]
- Shibuya, M. Vascular endothelial growth factor (VEGF) and its receptor (VEGFR) signaling in angiogenesis: A crucial target for anti- and pro-angiogenic therapies. Genes Cancer 2011, 2, 1097–1105. [Google Scholar] [CrossRef]
- Zeisler, H.; Llurba, E.; Chantraine, F.; Vatish, M.; Staff, A.C.; Sennström, M.; Olovsson, M.; Brennecke, S.P.; Stepan, H.; Allegranza, D.; et al. Predictive value of the sFlt-1:PlGF ratio in women with suspected preeclampsia. N. Engl. J. Med. 2016, 374, 13–22. [Google Scholar] [CrossRef]
- Andersen, L.B.; Golic, M.; Przybyl, L.; Sorensen, G.L.; Jørgensen, J.S.; Fruekilde, P.; von Versen-Höynck, F.; Herse, F.; Højskov, C.S.; Dechend, R.; et al. Vitamin D depletion does not affect key aspects of the preeclamptic phenotype in a transgenic rodent model for preeclampsia. J. Am. Soc. Hypertens. 2016, 10, 597–607.e1. [Google Scholar] [CrossRef]
- Lepercq, J.; Guerre-Millo, M.; André, J.; Caüzac, M.; Hauguel-De Mouzon, S. Leptin: A potential marker of placental insufficiency. Gynecol. Obstet. Investig. 2003, 55, 151–155. [Google Scholar] [CrossRef]
- Chen, J.; Khalil, R.A. Matrix metalloproteinases in normal pregnancy and preeclampsia. Physiol. Behav. 2017, 176, 87–165. [Google Scholar]
- Staun-Ram, E.; Goldman, S.; Gabarin, D.; Shalev, E. Expression and importance of matrix metalloproteinase 2 and 9 (MMP-2 and -9) in human trophoblast invasion. Reprod. Biol. Endocrinol. 2004, 2, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Ryu, J.S.; Dulay, A.; Segal, M.; Guller, S. Regulation of plasminogen activator inhibitor (PAI)-1 expression in a human trophoblast cell line by glucocorticoid (GC) and transforming growth factor (TGF)-β. Placenta 2002, 23, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.R.; Qiu, W.; Li, Y.X.; Zhang, Z.B.; Li, D.; Wang, Y.L. Dual effect of transforming growth factor β1 on cell adhesion and invasion in human placenta trophoblast cells. Reproduction 2006, 132, 333–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floridon, C.; Nielsen, O.; Holund, B.; Sunde, L.; Westergaard, J.G.; Thomsen, S.G.; Teisner, B. Localization of E-cadherin in villous, extravillous and vascular trophoblasts during intrauterine, ectopic and molar pregnancy. Mol. Hum. Reprod. 2000, 6, 943–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Li, X.; Zhao, Y.; Fang, C.; Lian, Y.; Gou, W.; Han, T.; Zhu, X. Insights into the mechanism of CXCL12-mediated signaling in trophoblast functions and placental angiogenesis. Acta Biochim. Biophys. Sin. 2015, 47, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, S.; Dinda, A.K.; Luthra, K. Differential expression of CXCR4 receptor in early and term human placenta. Placenta 2004, 25, 347–351. [Google Scholar] [CrossRef]
- Ren, L.; Liu, Y.Q.; Zhou, W.H.; Zhang, Y.Z. Trophoblast-derived chemokine CXCL12 promotes CXCR4 expression and invasion of human first-trimester decidual stromal cells. Hum. Reprod. 2012, 27, 366–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.H.; Du, M.R.; Dong, L.; Yu, J.; Li, D.J. Chemokine CXCL12 promotes the cross-talk between trophoblasts and decidual stromal cells in human first-trimester pregnancy. Hum. Reprod. 2008, 23, 2669–2679. [Google Scholar] [CrossRef] [Green Version]
- Jauniaux, E.; Watson, A.L.; Hempstock, J.; Bao, Y.P.; Skepper, J.N.; Burton, G.J. Onset of maternal arterial blood flow and placental oxidative stress: A possible factor in human early pregnancy failure. Am. J. Pathol. 2000, 157, 2111–2122. [Google Scholar] [CrossRef]
- Postovit, L.M.; Adams, M.A.; Graham, C.H. Does nitric oxide play a role in the aetiology of pre-eclampsia? Placenta 2001, 22, 51–55. [Google Scholar] [CrossRef]
- Caniggia, I.; Winter, J.; Lye, S.J.; Post, M. Oxygen and placental development during the first trimester: Implications for the pathophysiology of pre-eclampsia. Placenta 2000, 21, 25–30. [Google Scholar] [CrossRef]
- Knöfler, M.; Pollheimer, J. IFPA Award in Placentology Lecture: Molecular regulation of human trophoblast invasion. Placenta 2012, 33, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Reddy, T.E.; Pauli, F.; Sprouse, R.O.; Neff, N.F.; Newberry, K.M.; Garabedian, M.J.; Myers, R.M. Genomic determination of the glucocorticoid response reveals unexpected mechanisms of gene regulation. Genome Res. 2009, 19, 2163–2171. [Google Scholar] [CrossRef] [Green Version]
- John, S.; Johnson, T.A.; Sung, M.H.; Biddie, S.C.; Trump, S.; Koch-Paiz, C.A.; Davis, S.R.; Walker, R.; Meltzer, P.S.; Hager, G.L. Kinetic complexity of the global response to glucocorticoid receptor action. Endocrinology 2009, 150, 1766–1774. [Google Scholar] [CrossRef] [PubMed]
- Conradt, E.; Lester, B.M.; Appleton, A.A.; Armstrong, D.A.; Marsit, C.J. The roles of DNA methylation of NR3C1 and 11β-HSD2 and exposure to maternal mood disorder in utero on newborn neurobehavior. Epigenetics 2013, 8, 1321–1329. [Google Scholar] [CrossRef] [Green Version]
- Palma-Gudiel, H.; Córdova-Palomera, A.; Leza, J.C.; Fañanás, L. Glucocorticoid receptor gene (NR3C1) methylation processes as mediators of early adversity in stress-related disorders causality: A critical review. Neurosci. Biobehav. Rev. 2015, 55, 520–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagarajan, S.; Seddighzadeh, B.; Baccarelli, A.; Wise, L.A.; Williams, M.; Shields, A.E. Adverse maternal exposures, methylation of glucocorticoid-related genes and perinatal outcomes: A systematic review. Epigenomics 2016, 8, 925–944. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Húngaro, T.G.R.; Gregnani, M.F.; Alves-Silva, T.; Herse, F.; Alenina, N.; Bader, M.; Araújo, R.C. Cortisol Dose-Dependently Impairs Migration and Tube-like Formation in a Trophoblast Cell Line and Modulates Inflammatory and Angiogenic Genes. Biomedicines 2021, 9, 980. https://doi.org/10.3390/biomedicines9080980
Húngaro TGR, Gregnani MF, Alves-Silva T, Herse F, Alenina N, Bader M, Araújo RC. Cortisol Dose-Dependently Impairs Migration and Tube-like Formation in a Trophoblast Cell Line and Modulates Inflammatory and Angiogenic Genes. Biomedicines. 2021; 9(8):980. https://doi.org/10.3390/biomedicines9080980
Chicago/Turabian StyleHúngaro, Talita Guerreiro Rodrigues, Marcos F. Gregnani, Thaís Alves-Silva, Florian Herse, Natalia Alenina, Michael Bader, and Ronaldo C. Araújo. 2021. "Cortisol Dose-Dependently Impairs Migration and Tube-like Formation in a Trophoblast Cell Line and Modulates Inflammatory and Angiogenic Genes" Biomedicines 9, no. 8: 980. https://doi.org/10.3390/biomedicines9080980
APA StyleHúngaro, T. G. R., Gregnani, M. F., Alves-Silva, T., Herse, F., Alenina, N., Bader, M., & Araújo, R. C. (2021). Cortisol Dose-Dependently Impairs Migration and Tube-like Formation in a Trophoblast Cell Line and Modulates Inflammatory and Angiogenic Genes. Biomedicines, 9(8), 980. https://doi.org/10.3390/biomedicines9080980