
Differentiation Behaviour of Adipose-Derived Stromal Cells (ASCs) Seeded on Polyurethane-Fibrin Scaffolds In Vitro and In Vivo

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Studies
2.1.1. Isolation and Culture of Human Adipose-Derived Stem Cells (hASCs)
2.1.2. Fabrication of Polyurethane-Fibrin Composites
2.1.3. Scanning Electron Microscopy (SEM)
2.1.4. Chondrogenic Differentiation of PU-Fibrin Composites
- Group (DMEM): Control group maintained in expansion medium.
- 2.
- Group (BT): 50 ng/mL TGF-ß3 and 500 ng/mL BMP-6 (PromoCell GmbH, Heidelberg, Germany).
- 3.
- Group (BBT): 500 ng/mL BMP-6 for 7 days, followed by 50 ng/mL TGF-ß3 and 500 ng/mL BMP-6.
- 4.
- Group (TI): 50 ng/mL TGF-ß3 and 100 ng/mL IGF-1 (Sigma-Aldrich).
2.1.5. Histology and Immunohistochemistry
2.1.6. Real-Time Polymerase Chain Reaction (RT-PCR)
2.2. In Vivo Studies (New Zealand White Rabbits)
2.2.1. Harvesting of Adipose Tissue
2.2.2. Implantation of the PU-Fibrin Composites
2.2.3. Explantation of the PU-Fibrin Composites
2.2.4. Processing of the In Vivo Samples
2.3. Statistical Analyses
3. Results
3.1. In Vitro Studies
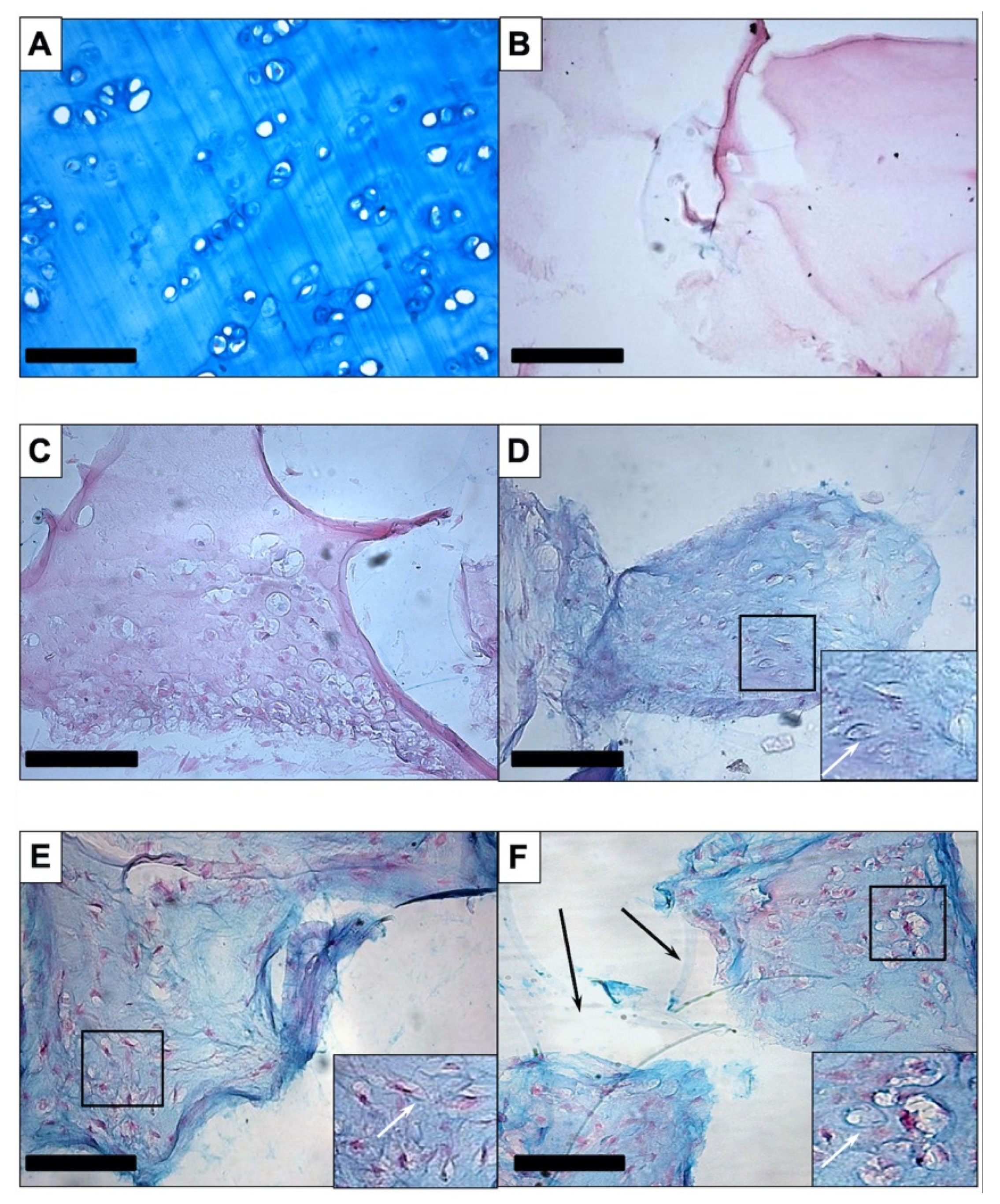
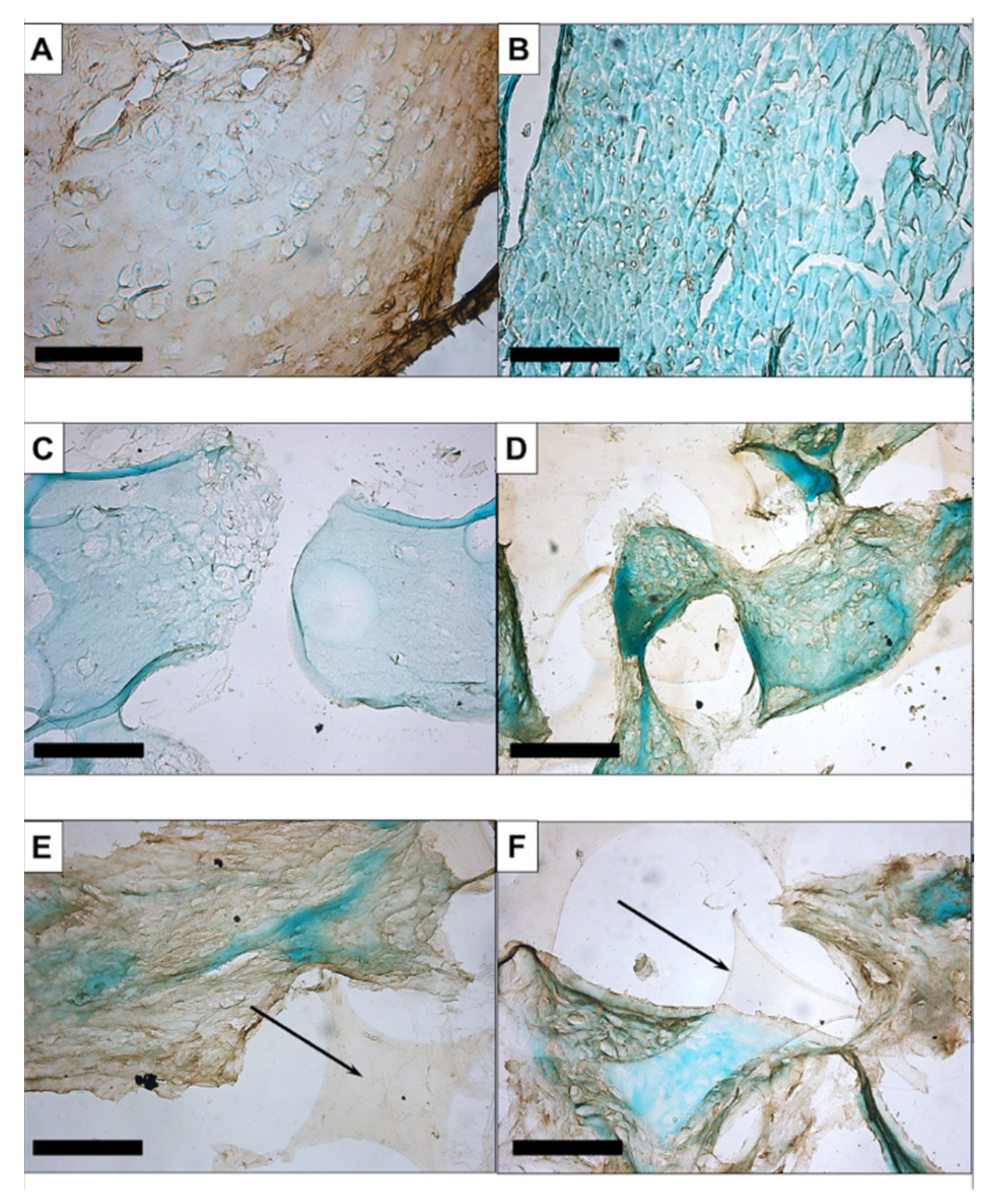
3.1.1. Histology and Immunohistochemistry
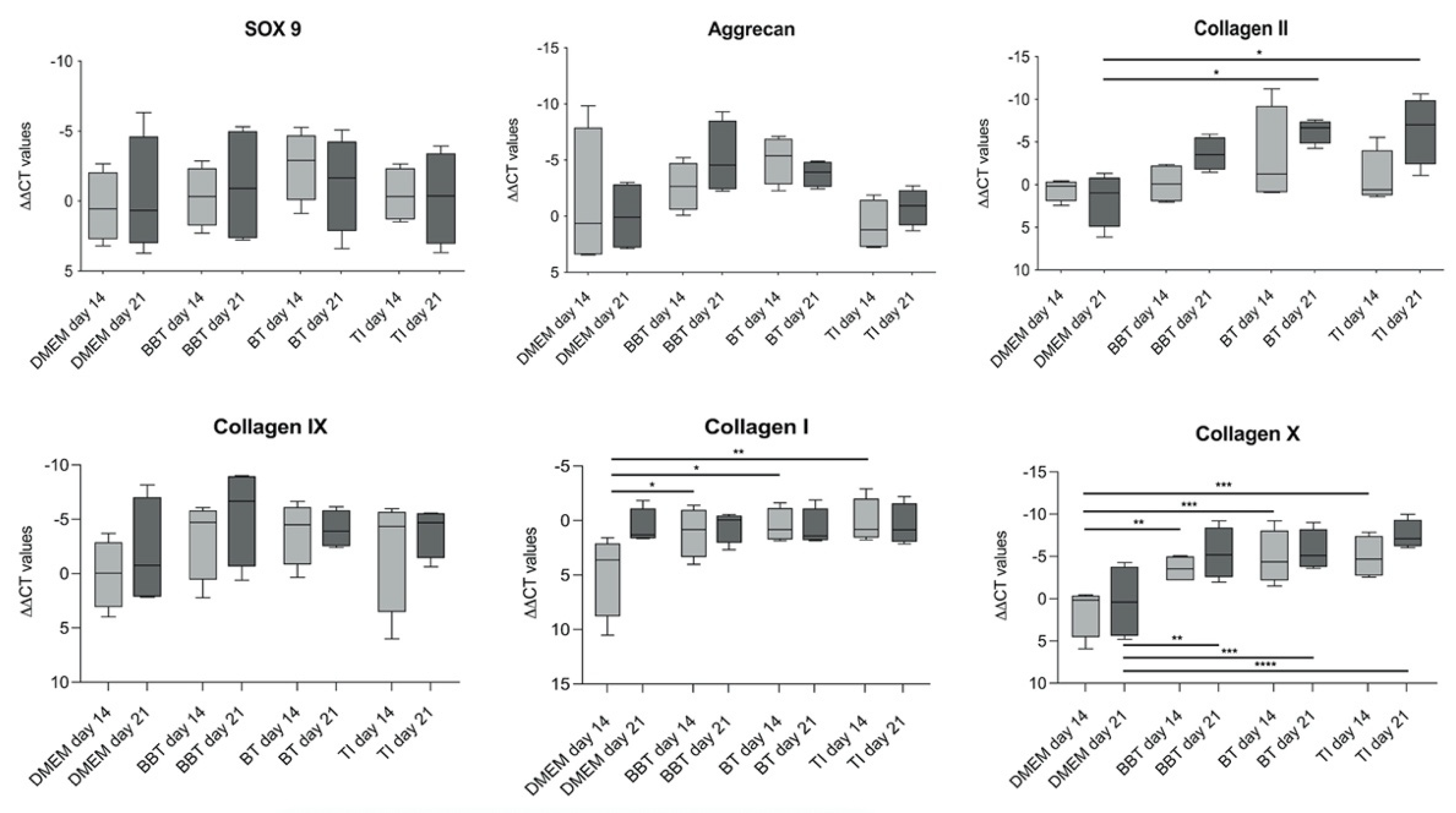
3.1.2. RT-PCR
3.1.3. Scanning Electron Microscopy (SEM)
3.2. In Vivo Experiments in New Zealand White Rabbits
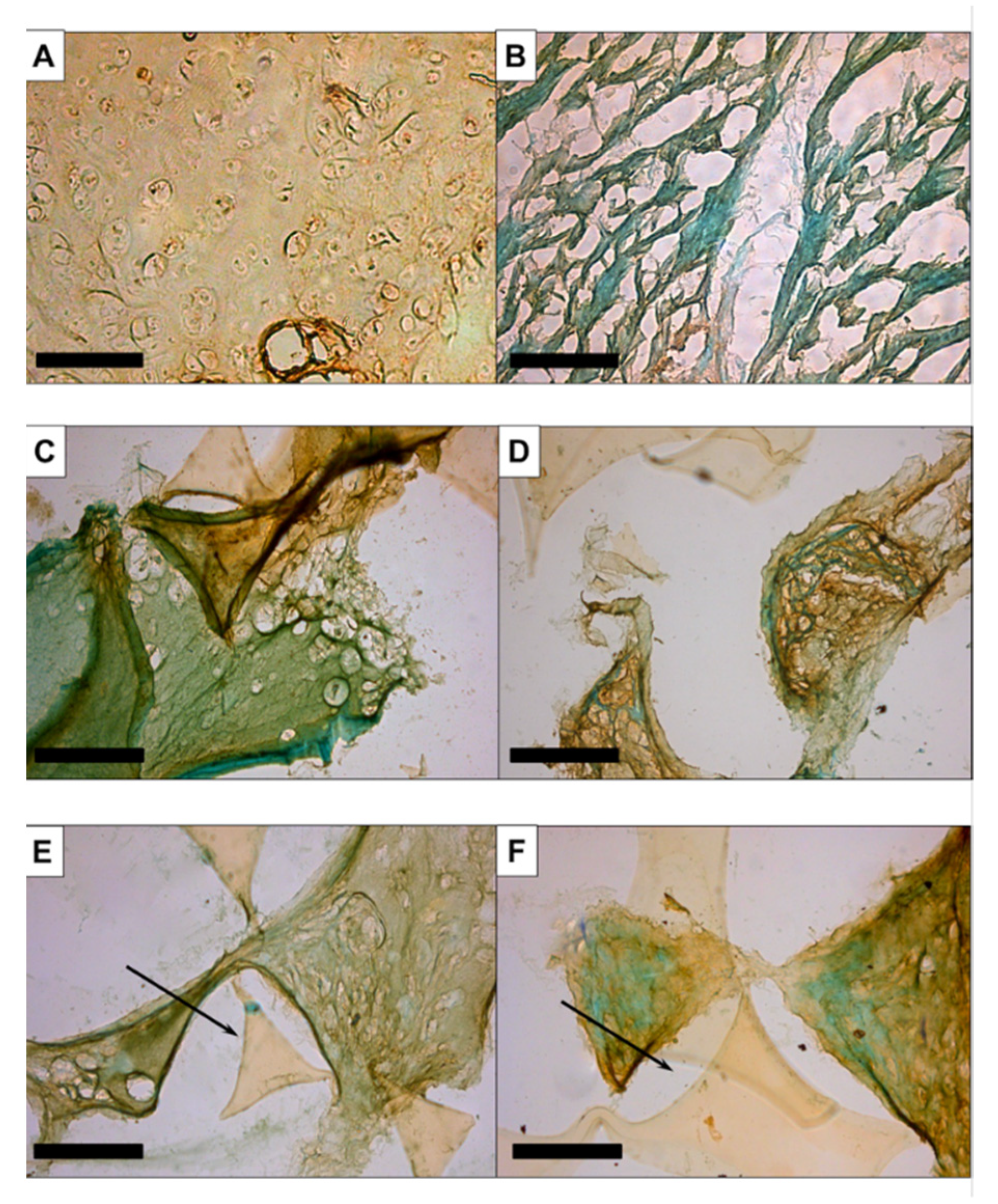
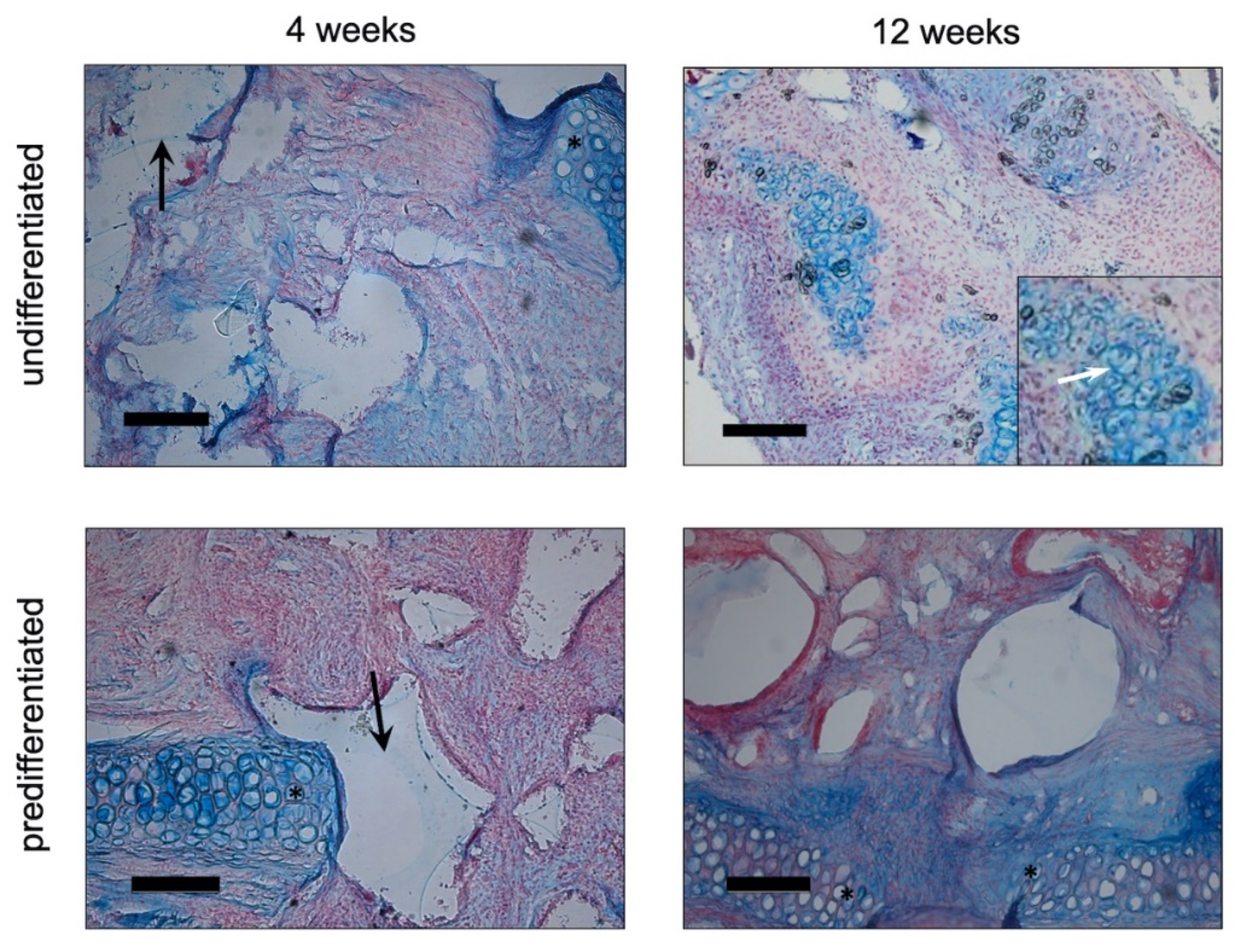
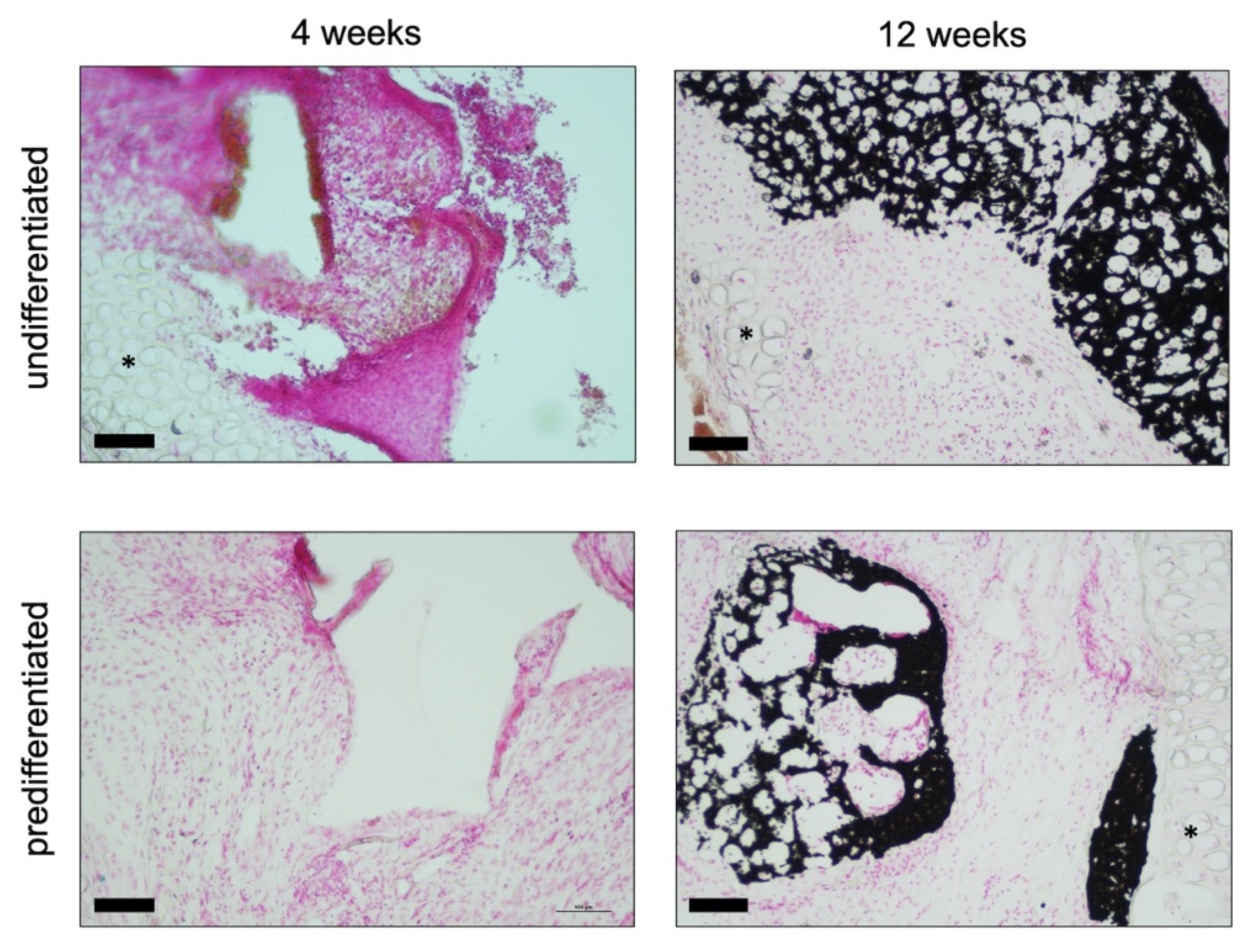
3.2.1. Histology
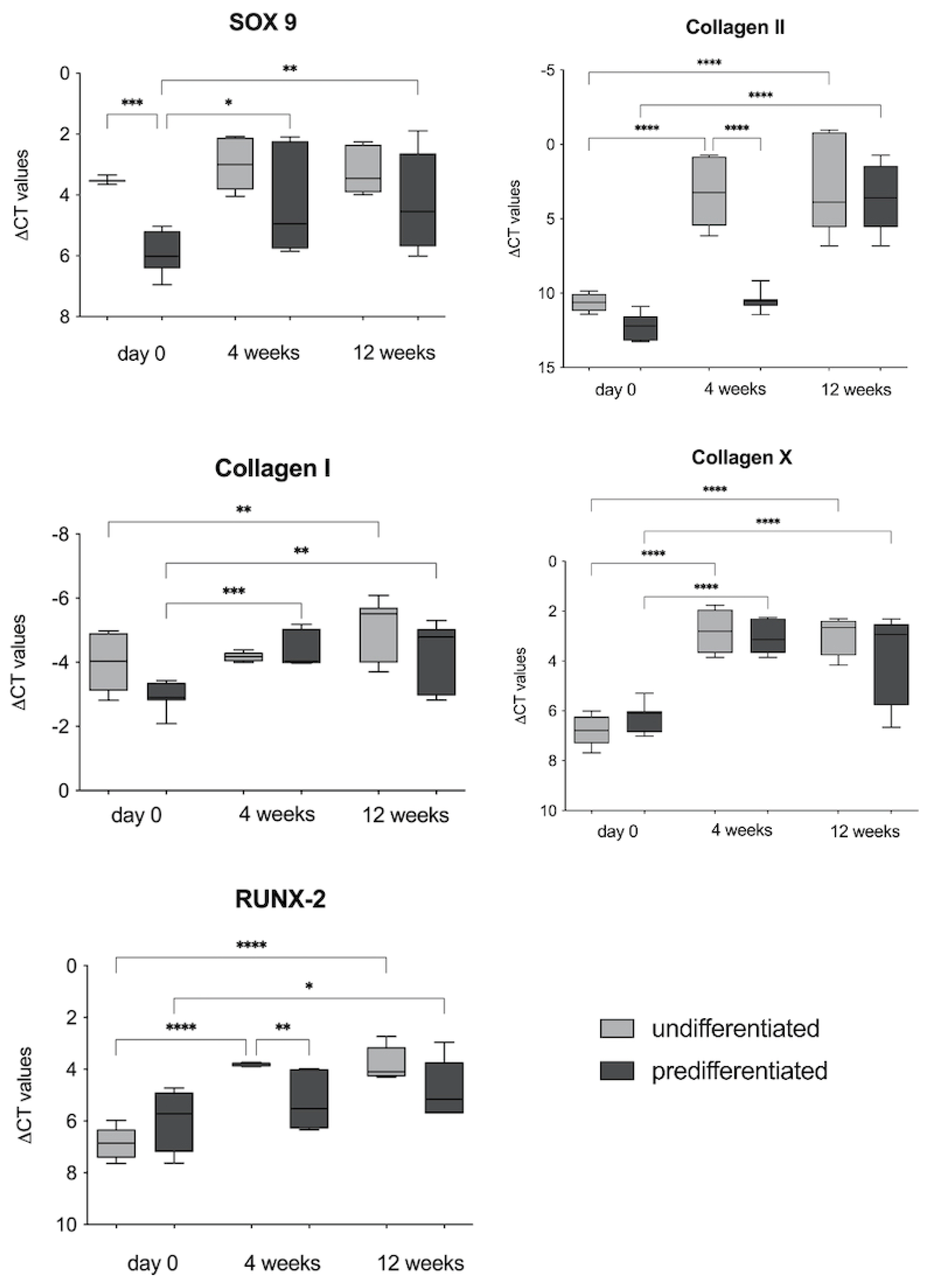
3.2.2. RT-PCR Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frölich, K.; Kleinsasser, N.; Rasp, G.; Staudenmaier, R. Navigation-assisted construction of an external ear canal using an autogenous foreskin graft. ORL J. Otorhinolaryngol. Relat. Spec. 2005, 67, 137–141. [Google Scholar] [CrossRef]
- Giacomini, P.G.; Ferraro, S.; Di Girolamo, S.; Ottaviani, F. Large nasal septal perforation repair by closed endoscopically assisted approach. Ann. Plast. Surg. 2011, 66, 633–636. [Google Scholar] [CrossRef]
- Detwiller, K.Y.; Schindler, J.S.; Schneider, D.S.; Lindau, R. Complex adult laryngotracheal reconstruction with a prefabricated flap: A case series. Head Neck 2013, 3, E376–E380. [Google Scholar] [CrossRef]
- Macchiarini, P.; Walles, T.; Biancosino, C.; Mertsching, H. First human transplantation of a bioengineered airway tissue. J. Thorac. Cardiovasc. Surg. 2004, 128, 638–641. [Google Scholar] [CrossRef] [PubMed]
- Von Bomhard, A.; Veit, J.; Bermueller, C.; Rotter, N.; Staudenmaier, R.; Storck, K.; The, H.N. Prefabrication of 3D cartilage contructs: Towards a tissue engineered auricle—A model tested in rabbits. PLoS ONE 2013, 8, e71667. [Google Scholar] [CrossRef] [PubMed]
- Fulco, I.; Miot, S.; Haug, M.D.; Barbero, A.; Wixmerten, A.; Feliciano, S.; Wolf, F.; Jundt, G.; Marsano, A.; Farhadi, J.; et al. Engineered autologous cartilage tissue for nasal reconstruction after tumour resection: An observational first-in-human trial. Lancet 2014, 384, 337–346. [Google Scholar] [CrossRef]
- Ansari, T.; Lange, P.; Southgate, A.; Greco, K.; Carvalho, C.; Partington, L.; Bullock, A.; MacNeil, S.; Lowdell, M.W.; Sibbons, P.D.; et al. Stem Cell-Based Tissue-Engineered Laryngeal Replacement. Stem Cells Transl. Med. 2017, 6, 677–687. [Google Scholar] [CrossRef]
- Herrmann, P.; Ansari, T.; Southgate, A.; Varanou Jenkins, A.; Partington, L.; Carvalho, C.; Janes, S.; Lowdell, M.; Sibbons, P.D.; Birchall, M.A. In vivo implantation of a tissue engineered stem cell seeded hemi-laryngeal replacement maintains airway, phonation, and swallowing in pigs. J. Tissue Eng. Regen. Med. 2019, 13, 1943–1954. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, G.; Cao, Y. Recent Progress in Cartilage Tissue Engineering—Our Experience and Future Directions. Engineering 2017, 3, 28–35. [Google Scholar] [CrossRef]
- Crawford, D.C.; DeBerardino, T.M.; Williams, R.J., 3rd. NeoCart, an autologous cartilage tissue implant, compared with microfracture for treatment of distal femoral cartilage lesions: An FDA phase-II prospective, randomized clinical trial after two years. J. Bone Jt. Surg. Am. 2012, 94, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Nöth, U.; Rackwitz, L.; Heymer, A.; Weber, M.; Baumann, B.; Steinert, A.; Schütze, N.; Jakob, F.; Eulert, J. Chondrogenic differentiation of human mesenchymal stem cells in collagen type I hydrogels. J. Biomed. Mater. Res. A 2007, 83, 626–635. [Google Scholar] [CrossRef] [PubMed]
- Bahrani, H.; Razmkhah, M.; Ashraf, M.J.; Tanideh, N.; Chenari, N.; Khademi, B.; Ghaderi, A. Differentiation of adipose-derived stem cells into ear auricle cartilage in rabbits. J. Laryngol. Otol. 2012, 126, 770–774. [Google Scholar] [CrossRef]
- Oedayrajsingh-Varma, M.J.; van Ham, S.M.; Knippenberg, M.; Helder, M.N.; Klein-Nulend, J.; Schouten, T.E.; Ritt, M.J.; van Milligen, F.J. Adipose tissue-derived mesenchymal stem cell yield and growth characteristics are affected by the tissue-harvesting procedure. Cytotherapy 2006, 8, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Housman, T.S.; Lawrence, N.; Mellen, B.G.; George, M.N.; Filippo, J.S.; Cerveny, K.A.; DeMarco, M.; Feldman, S.R.; Fleischer, A.B. The safety of liposuction: Results of a national survey. Dermatol. Surg. 2002, 28, 971–978. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, D.J.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Aust, L.; Devlin, B.; Foster, S.J.; Halvorsen, Y.D.; Hicok, K.; du Laney, T.; Sen, A.; Willingmyre, G.D.; Gimble, J.M. Yield of human adipose-derived adult stem cells from liposuction aspirates. Cytotherapy 2004, 6, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Strem, B.M.; Hicok, K.C.; Zhu, M.; Wulur, I.; Alfonso, Z.; Schreiber, R.E.; Fraser, J.K.; Hedrick, M.H. Multipotential differentiation of adipose tissue-derived stem cells. Keio J. Med. 2005, 54, 132–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- De Ugarte, D.A.; Morizono, K.; Elbarbary, A.; Alfonso, Z.; Zuk, P.A.; Zhu, M.; Dragoo, J.L.; Ashjian, P.; Thomas, B.; Benhaim, P.; et al. Comparison of multi-lineage cells from human adipose tissue and bone marrow. Cells Tissues Organs 2003, 174, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Guilak, F.; Awad, H.A.; Fermor, B.; Leddy, H.A.; Gimble, J.M. Adipose-derived adult stem cells for cartilage tissue engineering. Biorheology 2004, 41, 389–399. [Google Scholar] [PubMed]
- Bunnell, B.A.; Flaat, M.; Gagliardi, C.; Patel, B.; Ripoll, C. Adipose-derived stem cells: Isolation, expansion and differentiation. Methods 2008, 45, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Bourin, P.; Bunnell, B.A.; Casteilla, L.; Dominici, M.; Katz, A.J.; March, K.L.; Redl, H.; Rubin, J.P.; Yoshimura, K.; Gimble, J.M. Stromal cells from the adipose tissue-derived stromal vascular fraction and culture expanded adipose tissue-derived stromal/stem cells: A joint statement of the International Federation for Adipose Therapeutics and Science (IFATS) and the International Society for Cellular Therapy (ISCT). Cytotherapy 2013, 15, 641–648. [Google Scholar]
- Mooney, D.J.; Vandenburgh, H. Cell Delivery Mechanisms for Tissue Repair. Cell Stem Cell 2008, 2, 205–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, G.I. Regeneration of articular cartilage using adipose stem cells. J. Biomed. Mater. Res. A 2016, 104, 1830–1844. [Google Scholar] [CrossRef] [PubMed]
- Hennig, T.; Lorenz, H.; Thiel, A.; Goetzke, K.; Dickhut, A.; Geiger, F.; Richter, W. Reduced chondrogenic potential of adipose tissue derived stromal cells correlates with an altered TGFbeta receptor and BMP profile and is overcome by BMP-6. J. Cell Physiol. 2007, 211, 682–691. [Google Scholar] [CrossRef]
- Mehlhorn, A.T.; Niemeyer, P.; Kaschte, K.; Muller, L.; Finkenzeller, G.; Hartl, D.; Sudkamp, N.P.; Schmal, H. Differential effects of BMP-2 and TGF-β1 on chondrogenic differentiation of adipose derived stem cells. Cell Prolif. 2007, 4, 809–823. [Google Scholar] [CrossRef] [PubMed]
- Ude, C.C.; Chen, H.C.; Norhamdan, M.Y.; Azizi, B.M.; Aminuddin, B.S.; Ruszymah, B.H.I. The evaluation of cartilage differentiations using transforming growth factor beta3 alone and with combination of bone morphogenetic protein-6 on adult stem cells. Cell Tissue Bank 2017, 18, 355–367. [Google Scholar] [CrossRef]
- Froelich, K.; Setiawan, L.E.; Technau, A.; Tirado, M.R.; Hackenberg, S.; Hagen, R.; Staudenmaier, R.; Kleinsasser, N.H. Influence of different growth factors on chondrogenic differentiation of adipose-derived stem cells in polyurethane-fibrin composites. Int J. Artif. Organs 2012, 35, 1047–1060. [Google Scholar] [CrossRef]
- Diekman, B.O.; Rowland, C.R.; Lennon, D.P.; Caplan, A.I.; Guilak, F. Chondrogenesis of Adult Stem Cells from Adipose Tissue and Bone Marrow: Induction by Growth Factors and Cartilage-Derived Matrix. Tissue Eng. Part A 2010, 16, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Seitz, D.; Chevalier, Y.; Müller, P.E.; Jansson, V.; Klar, R.M. Synergistic interaction of hTGF-β 3 with hBMP-6 promotes articular cartilage formation in chitosan scaffolds with hADSCs: Implications for regenerative medicine. BMC Biotechnol. 2020, 20, 48. [Google Scholar] [CrossRef]
- Moradian Tehrani, R.; Mirzaei, H.; Verdi, J.; Sahebkar, A.; Noureddini, M.; Salehi, R.; Alani, B.; Kianmehr, M. Chondrogenic differentiation of human scalp adipose-derived stem cells in Polycaprolactone scaffold and using Freeze Thaw Freeze method. J. Cell Physiol. 2018, 233, 6705–6713. [Google Scholar] [CrossRef]
- Kim, H.J.; Im, G.I. Chondrogenic differentiation of adipose tissue-derived mesenchymal stem cells: Greater doses of growth factor are necessary. J. Orthop. Res. 2008, 27, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Goh, B.S.; Che Omar, S.N.; Ubaidah, M.A.; Saim, L.; Sulaiman, S.; Chua, K.H. Chondrogenesis of human adipose derived stem cells for future microtia repair using co-culture technique. Acta Otolaryngol. 2017, 137, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Kupcsik, L.; Yao, S.J.; Alini, M.; Stoddart, M.J. Chondrogenesis of human bone marrow mesenchymal stem cells in fibrin-polyurethane composites. Tissue Eng. Part A 2009, 15, 1729–1737. [Google Scholar] [CrossRef]
- Wittmann, K.; Storck, K.; Muhr, C.; Mayer, H.; Regn, S.; Staudenmaier, R.; Wiese, H.; Maier, G.; Bauer-Kreisel, P.; Blunk, T. Development of volume-stable adipose tissue constructs using polycaprolactone-based polyurethane scaffolds and fibrin hydrogels. J. Tissue Eng. Regen. Med. 2016, 10, E409–E418. [Google Scholar] [CrossRef] [PubMed]
- Eyrich, D.; Wiese, H.; Maier, G.; Skodacek, D.; Appel, B.; Sarhan, H.; Tessmar, J.; Staudenmaier, R.; Wenzel, M.M.; Goepferich, A.; et al. In vitro and in vivo cartilage engineering using a combination of chondrocyte-seeded long-term stable fibrin gels and polycaprolactone-based polyurethane scaffolds. Tissue Eng. 2007, 13, 2207–2218. [Google Scholar] [CrossRef]
- Grad, S.; Kupcsik, L.; Gorna, K.; Gogolewski, S.; Alini, M. The use of biodegradable polyurethane scaffolds for cartilage tissue engineering: Potential and limitations. Biomaterials 2003, 24, 5163–5171. [Google Scholar] [CrossRef]
- Lee, C.R.; Grad, S.; Gorna, K.; Gogolewski, S.; Goessl, A.; Alini, M. Fibrin-polyurethane composites for articular cartilage tissue engineering: A preliminary analysis. Tissue Eng. 2005, 11, 1562–1573. [Google Scholar] [CrossRef] [PubMed]
- Froelich, K.; Mickler, J.; Steusloff, G.; Technau, A.; Ramos Tirado, M.; Scherzed, A.; Hackenberg, S.; Radeloff, A.; Hagen, R.; Kleinsasser, N. Chromosomal aberrations and deoxyribonucleic acid single-strand breaks in adipose-derived stem cells during long-term expansion in vitro. Cytotherapy 2013, 15, 767–781. [Google Scholar] [CrossRef]
- Radeloff, K.; Ramos Tirado, M.; Haddad, D.; Breuer, K.; Müller, J.; Hochmuth, S.; Hackenberg, S.; Scherzad, A.; Kleinsasser, N.; Radeloff, A. Superparamagnetic Iron Oxide Particles (VSOPs) Show Genotoxic Effects but No Functional Impact on Human Adipose Tissue-Derived Stromal Cells (ASCs). Materials 2021, 14, 263. [Google Scholar] [CrossRef]
- Lev, R.; Spicer, S.S. Specific staining of sulphate groups with alcian blue at low pH. J. Histochem. Cytochem. 1964, 12, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ten Koppel, P.G.; van Osch, G.J.; Verwoerd, C.D.; Verwoerd-Verhoef, H.L. A new in vivo model for testing cartilage grafts and biomaterials: The ‘rabbit pinna punch-hole’ model. Biomaterials 2001, 22, 1407–1414. [Google Scholar] [CrossRef]
- Gentile, P.; Piccinno, M.S.; Calabrese, C. Characteristics and Potentiality of Human Adipose-Derived Stem Cells (hASCs) Obtained from Enzymatic Digestion of Fat Graft. Cells 2019, 8, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentile, P.; Scioli, M.G.; Bielli, A.; Orlandi, A.; Cervelli, V. Concise Review: The Use of Adipose-Derived Stromal Vascular Fraction Cells and Platelet Rich Plasma in Regenerative Plastic Surgery. Stem Cells 2017, 35, 117–134. [Google Scholar] [CrossRef]
- Gentile, P.; Calabrese, C.; De Angelis, B.; Pizzicannella, J.; Kothari, A.; Garcovich, S. Impact of the Different Preparation Methods to Obtain Human Adipose-Derived Stromal Vascular Fraction Cells (AD-SVFs) and Human Adipose-Derived Mesenchymal Stem Cells (AD-MSCs): Enzymatic Digestion Versus Mechanical Centrifugation. Int. J. Mol. Sci. 2019, 20, 5471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentile, P.; Kothari, A.; Casella, D.; Calabrese, C. Fat Graft Enhanced with Adipose-Derived Stem Cells in Aesthetic Breast Augmentation: Clinical, Histological, and Instrumental Evaluation. Aesthet. Surg. J. 2020, 40, 962–977. [Google Scholar] [CrossRef]
- Gentile, P.; De Angelis, B.; Pasin, M.; Cervelli, G.; Curcio, C.B.; Floris, M.; Di Pasquali, C.; Bocchini, I.; Balzani, A.; Nicoli, F.; et al. Adipose-derived stromal vascular fraction cells and platelet-rich plasma: Basic and clinical evaluation for cell-based therapies in patients with scars on the face. J. Craniofac. Surg. 2014, 25, 267–272. [Google Scholar] [CrossRef]
- Gentile, P.; Sterodimas, A.; Pizzicannella, J.; Dionisi, L.; De Fazio, D.; Calabrese, C.; Garcovich, S. Systematic Review: Allogenic Use of Stromal Vascular Fraction (SVF) and Decellularized Extracellular Matrices (ECM) as Advanced Therapy Medicinal Products (ATMP) in Tissue Regeneration. Int. J. Mol. Sci. 2020, 21, 4982. [Google Scholar] [CrossRef]
- Gentile, P.; Garcovich, S. Concise Review: Adipose-Derived Stem Cells (ASCs) and Adipocyte-Secreted Exosomal microRNA (A-SE-miR) Modulate Cancer Growth and proMote Wound Repair. J. Clin. Med. 2019, 8, 855. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, D.; Sato, K.; Gonda, K.; Takaki, Y.; Shigeura, T.; Sato, T.; Aiba-Kojima, E.; Lizuka, F.; Inoue, K.; Suga, H.; et al. Cell-assisted lipotransfer: Supportive use of human adipose-derived cells for soft tissue augmentation with lipoinjection. Tissue Eng. 2006, 12, 3375–3382. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, K.; Sato, K.; Aoi, N.; Kurita, M.; Inoue, K.; Suga, H.; Eto, H.; Kato, H.; Hirohi, T.; Harii, K. Cell-assisted lipotransfer for facial lipoatrophy: Efficacy of clinical use of adipose-derived stem cells. Dermatol. Surg. 2008, 34, 1178–1185. [Google Scholar] [CrossRef]
- Zielins, E.R.; Luan, A.; Brett, E.A.; Longaker, M.T.; Wan, D.C. Therapeutic applications of human adipose-derived stromal cells for soft tissue reconstruction. Discov. Med. 2015, 19, 245–253. [Google Scholar]
- Scioli, M.G.; Bielli, A.; Gentile, P.; Cervelli, V.; Orlandi, A. Combined treatment with platelet-rich plasma and insulin favours chondrogenic and osteogenic differentiation of human adipose-derived stem cells in three-dimensional collagen scaffolds. J. Tissue Eng. Regen. Med. 2017, 11, 2398–2410. [Google Scholar] [CrossRef]
- Bielli, A.; Scioli, M.G.; Gentile, P.; Cervelli, V.; Orlandi, A. Adipose-derived stem cells in cartilage regeneration: Current perspectives. Regen. Med. 2016, 11, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Im, G.I.; Shin, Y.W.; Lee, K.B. Do adipose tissue-derived mesenchymal stem cells have the same osteogenic and chondrogenic potential as bone marrow-derived cells? Osteoarthr. Cartil. 2005, 13, 845–853. [Google Scholar] [CrossRef] [Green Version]
- Pleumeekers, M.M.; Nimeskern, L.; Koevoet, J.L.M.; Karperien, M.; Stok, K.S.; van Osch, G.J.V.M. Trophic effects of adipose-tissue-derived and bone-marrow-derived mesenchymal stem cells enhance cartilage generation by chondrocytes in co-culture. PLoS ONE 2018, 13, e0190744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derks, M.; Sturm, T.; Haverich, A.; Hilfiker, A. Isolation and chondrogenic differentiation of porcine perichondrial progenitor cells for the purpose of cartilage tissue engineering. Cells Tissues Organs 2013, 198, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Otto, I.A.; Levato, R.; Webb, W.R.; Khan, I.M.; Breugem, C.C.; Malda, J. Progenitor cells in auricular cartilage demonstrate cartilage-forming capacity in 3D hydrogel culture. Eur. Cell Mater. 2018, 35, 132–150. [Google Scholar] [CrossRef]
- Kobayashi, S.; Takebe, T.; Zheng, Y.W.; Mizuno, M.; Yabuki, Y.; Maegawa, J.; Taniguchi, H. Presence of cartilage stem/progenitor cells in adult mice auricular perichondrium. PLoS ONE 2011, 6, e26393. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Takebe, T.; Inui, M.; Iwai, S.; Kan, H.; Zheng, Y.W.; Maegawa, J.; Taniguchi, H. Reconstruction of human elastic cartilage by a CD44+ CD90+ stem cell in the ear perichondrium. Proc. Natl. Acad. Sci. USA 2011, 108, 14479–14484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gvaramia, D.; Kern, J.; Jakob, Y.; Zenobi-Wong, M.; Rotter, N. Regenerative Potential of Perichondrium: A Tissue Engineering Perspective. Tissue Eng. Part B Rev. 2021. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Park, H.Y.; Choi, K.U.; Choi, S.W.; Kim, S.D.; Kong, S.K.; Cho, K.S. Auricular Cartilage Regeneration with Adipose-Derived Stem Cells in Rabbits. Mediat. Inflamm. 2018, 2018, 4267158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrell, E.; Both, S.K.; Odörfer, K.I.; Koevoet, W.; Kops, N.; O’Brien, F.J.; Baatenburg de Jong, R.J.; Verhaar, J.A.; Cuijpers, V.; Jansen, J.; et al. In-vivo generation of bone via endochondral ossification by in-vitro chondrogenic priming of adult human and rat mesenchymal stem cells. BMC Musculoskelet. Disord. 2011, 12, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, K.; Felimban, R.; Traianedes, K.; Moulton, S.E.; Wallace, G.G.; Chung, J.; Quigley, A.; Choong, P.F.; Myers, D.E. Chondrogenesis of Infrapatellar Fat Pad Derived Adipose Stem Cells in 3D Printed Chitosan Scaffold. PLoS ONE 2014, 9, e99410. [Google Scholar] [CrossRef]
- Osinga, R.; Di Maggio, N.; Todorov, A.; Allafi, N.; Barbero, A.; Laurent, F.; Schaefer, D.J.; Martin, I.; Scherberich, A. Generation of a Bone Organ by Human Adipose-Derived Stromal Cells Through Endochondral Ossification. Stem Cells Transl. Med. 2016, 5, 1090–1097. [Google Scholar] [CrossRef]
- Kuhlmann, C.; Schenck, T.L.; Tluczynski, K.; Aszodi, A.; Metzger, P.; Giunta, R.; Wiggenhauser, P.S. Experimental approach to nasal septal cartilage regeneration with adipose tissue-derived stem cells and decellularized porcine septal cartilage. Xenotransplantation 2021, 28, e12660. [Google Scholar] [CrossRef]
- Sriwatananukulkit, O.; Tawonsawatruk, T.; Rattanapinyopituk, K.; Luangwattanawilai, T.; Srikaew, N.; Hemstapat, R. Scaffold-Free Cartilage Construct from Infrapatellar Fat Pad Stem Cells for Cartilage Restoration. Tissue Eng. Part A 2020. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radeloff, K.; Weiss, D.; Hagen, R.; Kleinsasser, N.; Radeloff, A. Differentiation Behaviour of Adipose-Derived Stromal Cells (ASCs) Seeded on Polyurethane-Fibrin Scaffolds In Vitro and In Vivo. Biomedicines 2021, 9, 982. https://doi.org/10.3390/biomedicines9080982
Radeloff K, Weiss D, Hagen R, Kleinsasser N, Radeloff A. Differentiation Behaviour of Adipose-Derived Stromal Cells (ASCs) Seeded on Polyurethane-Fibrin Scaffolds In Vitro and In Vivo. Biomedicines. 2021; 9(8):982. https://doi.org/10.3390/biomedicines9080982
Chicago/Turabian StyleRadeloff, Katrin, Dorothee Weiss, Rudolf Hagen, Norbert Kleinsasser, and Andreas Radeloff. 2021. "Differentiation Behaviour of Adipose-Derived Stromal Cells (ASCs) Seeded on Polyurethane-Fibrin Scaffolds In Vitro and In Vivo" Biomedicines 9, no. 8: 982. https://doi.org/10.3390/biomedicines9080982
APA StyleRadeloff, K., Weiss, D., Hagen, R., Kleinsasser, N., & Radeloff, A. (2021). Differentiation Behaviour of Adipose-Derived Stromal Cells (ASCs) Seeded on Polyurethane-Fibrin Scaffolds In Vitro and In Vivo. Biomedicines, 9(8), 982. https://doi.org/10.3390/biomedicines9080982