
Effect of Microparticles on Fungal Fermentation for Fermentation-Based Product Productions




Abstract
:1. Introduction
2. Advantages and Disadvantages of Fungal Fermentation
3. Fungal Cell Growth Characteristics (Morphology) and Effects on the Product Formation
4. Conventional Methods for Controlling Fungal Cell Growth in Liquid Fermentation
5. Microparticles for Controlling Fungal Cell Growth in Liquid Fermentation
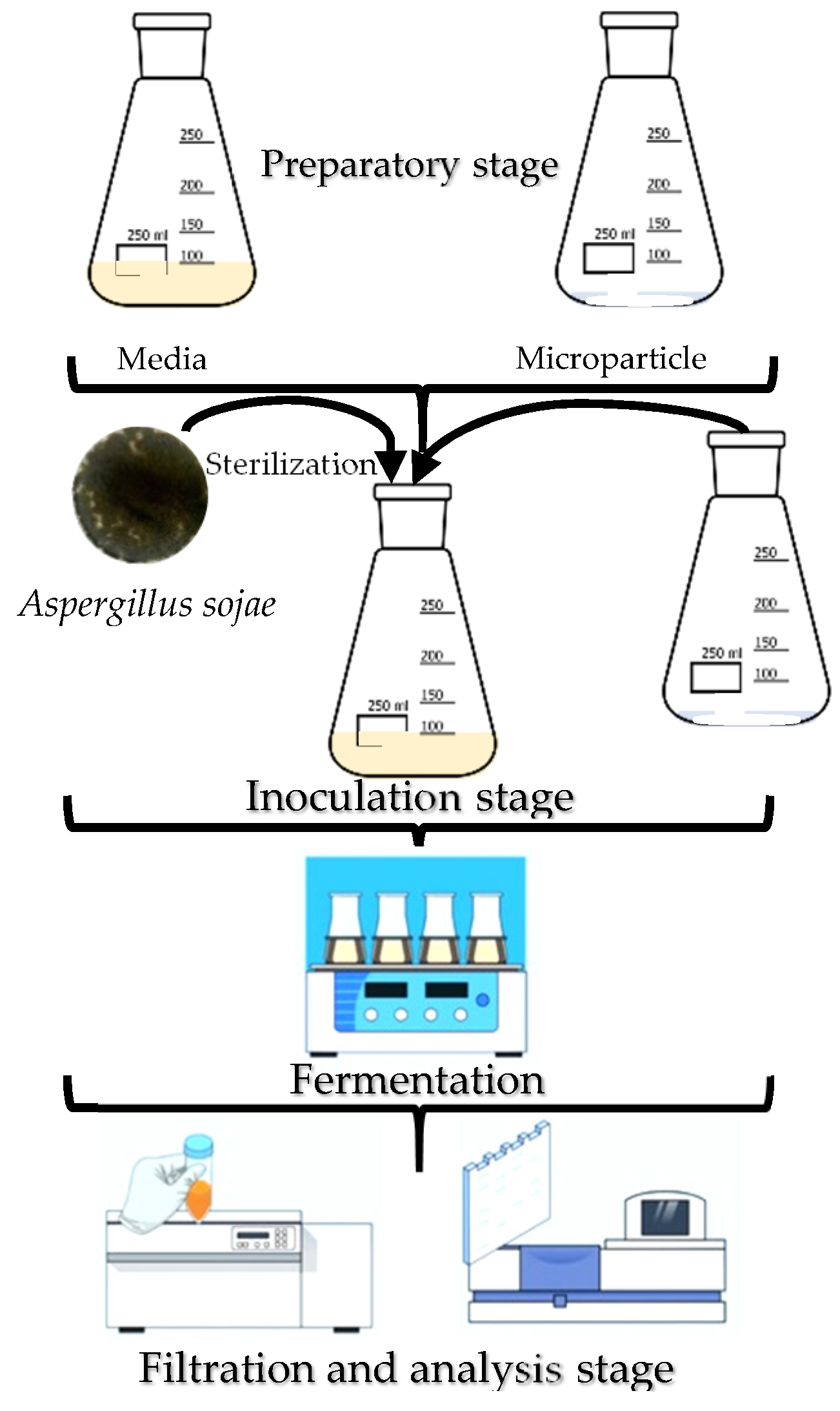
5.1. Microparticles
5.2. Mechanism of Microparticles in Controlling Fungal Cell Growth
6. Applications of Microparticles in Liquid Fermentation for the Productions of Value-Added Products
6.1. Enzyme Production
6.2. Organic Acid
6.3. Pigments
6.4. Lipids
6.5. Polysaccharides
7. Conclusions and Future Trends
Funding
Acknowledgments
Conflicts of Interest
References
- Cairns, T.C.; Nai, C.; Meyer, V. How a fungus shapes biotechnology: 100 years of Aspergillus niger research. Fungal Biol. Biotechnol. 2018, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavanagh, K. Fungal fermentation systems and products. In Fungi: Biology and Applications; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; p. 125. [Google Scholar] [CrossRef]
- Walker, G.M.; White, N.A. Introduction to fungal physiology. In Fungi: Biology and Applications; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 1–35. [Google Scholar] [CrossRef] [Green Version]
- Papagianni, M. Fungal morphology and metabolite production in submerged mycelial processes. Biotechnol. Adv. 2004, 22, 189–259. [Google Scholar] [CrossRef] [PubMed]
- Amore, A.; Giacobbe, S.; Faraco, V. Regulation of cellulase and hemicellulase gene expression in fungi. Curr. Genom. 2013, 14, 230–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkinson, B.; Daoud, I.S. Microbial floes and flocculation in fermentation process engineering. In Advances in Biochemical Engineering; Springer: Berlin/Heidelberg, Germany, 1976; Volume 4, pp. 41–124. [Google Scholar] [CrossRef]
- Yatmaz, E.; Karahalil, E.; Germec, M.; Ilgin, M.; Turhan, I. Controlling filamentous fungi morphology with microparticles to enhanced β-mannanase production. Bioprocess Biosyst. Eng. 2016, 39, 1391–1399. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, A.; Boruta, T.; Bizukojć, M. Performance of fungal microparticle-enhanced cultivations in stirred tank bioreactors depends on species and number of process stages. Biochem. Eng. J. 2020, 161, 107696. [Google Scholar] [CrossRef]
- Coban, H.B.; Demirci, A.; Turhan, I. Microparticle-enhanced Aspergillus ficuum phytase production and evaluation of fungal morphology in submerged fermentation. Bioprocess Biosyst. Eng. 2015, 38, 1075–1080. [Google Scholar] [CrossRef]
- Kaup, B.; Ehrich, K.; Pescheck, M.; Schrader, J. Microparticle-enhanced cultivation of filamentous microorganisms: Increased chloroperoxidase formation by Caldariomyces fumago as an example. Biotechnol. Bioeng. 2008, 99, 491–498. [Google Scholar] [CrossRef]
- Hölker, U.; Höfer, M.; Lenz, J. Biotechnological advantages of laboratory-scale solid-state fermentation with fungi. Appl. Microbiol. Biotechnol. 2004, 64, 175–186. [Google Scholar] [CrossRef]
- El-Enshasy, H.A. Filamentous fungal cultures–process characteristics, products, and applications. In Bioprocessing for Value-Added Products from Renewable Resources; Elsevier: Amsterdam, The Netherlands, 2007; pp. 225–261. [Google Scholar] [CrossRef]
- Iram, A.; Cekmecelioglu, D.; Demirci, A. Screening of bacterial and fungal strains for cellulase and xylanase production using distillers’ dried grains with solubles (DDGS) as the main feedstock. Biomass Convers. Biorefinery 2020, 11, 1955–1964. [Google Scholar] [CrossRef]
- Patakova, P. Monascus secondary metabolites: Production and biological activity. J. Ind. Microbiol. Biotechnol. 2013, 40, 169–181. [Google Scholar] [CrossRef]
- Jun, H.; Kieselbach, T.; Jönsson, L.J. Enzyme production by filamentous fungi: Analysis of the secretome of Trichoderma reesei grown on unconventional carbon source. Microb. Cell Factories 2011, 10, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, G.B.; de Sousa Francisco Filho, Á.; da Silva Rodrigues, J.R.; de Souza, R.R. Cellulase production by Aspergillus niger using urban lignocellulosic waste as substrate: Evaluation of different cultivation strategies. J. Environ. Manag. 2022, 305, 114431. [Google Scholar] [CrossRef] [PubMed]
- Muñiz-Márquez, D.B.; Teixeira, J.A.; Mussatto, S.I.; Contreras-Esquivel, J.C.; Rodríguez-Herrera, R.; Aguilar, C.N. Fructo-oligosaccharides (FOS) production by fungal submerged culture using aguamiel as a low-cost by-product. LWT 2019, 102, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Tong, Z.; Tong, Y.; Wang, D.; Shi, Y. Whole maize flour and isolated maize starch for production of citric acid by Aspergillus niger: A review. Starch-Stärke 2021, 2000014. [Google Scholar] [CrossRef]
- Ire, F.S.; Chima, I.J.; Ezebuiro, V. Enhanced xylanase production from UV-mutated Aspergillus niger grown on corn cob and sawdust. Biocatal. Agric. Biotechnol. 2021, 31, 101869. [Google Scholar] [CrossRef]
- Ismail, S.A.; Nour, S.A.; Hassan, A.A. Valorization of corn cobs for xylanase production by Aspergillus flavus AW1 and its application in the production of antioxidant oligosaccharides and removal of food stain. Biocatal. Agric. Biotechnol. 2022, 41, 102311. [Google Scholar] [CrossRef]
- Ravichandra, K.; Balaji, R.; Devarapalli, K.; Cheemalamarri, C.; Poda, S.; Banoth, L.; Pinnamaneni, S.R.; Prakasham, R.S. Impact of structure and composition of different sorghum xylans as substrates on production of xylanase enzyme by Aspergillus fumigatus RSP-8. Ind. Crops Prod. 2022, 188, 115660. [Google Scholar] [CrossRef]
- Chergui, D.; Akretche-Kelfat, S.; Lamoudi, L.; Al-Rshaidat, M.; Boudjelal, F.; Ait-Amar, H. Optimization of citric acid production by Aspergillus niger using two downgraded Algerian date varieties. Saudi J. Biol. Sci. 2021, 28, 7134–7141. [Google Scholar] [CrossRef]
- Nema, A.; Patnala, S.H.; Mandari, V.; Kota, S.; Devarai, S.K. Production and optimization of lipase using Aspergillus niger MTCC 872 by solid-state fermentation. Bull. Natl. Res. Cent. 2019, 43, 82. [Google Scholar] [CrossRef] [Green Version]
- Iram, A.; Cekmecelioglu, D.; Demirci, A. Salt and nitrogen amendment and optimization for cellulase and xylanase production using dilute acid hydrolysate of distillers’ dried grains with solubles (DDGS) as the feedstock. Bioprocess Biosyst. Eng. 2022, 45, 527–540. [Google Scholar] [CrossRef]
- de Souza, J.B.; Michelin, M.; Amâncio, F.L.R.; Brazil, O.A.V.; Maria de Lourdes, T.M.; Ruzene, D.S.; Silva, D.P.; Mendonça, M.D.C.; López, J.A. Sunflower stalk as a carbon source inductive for fungal xylanase production. Ind. Crops Prod. 2020, 153, 112368. [Google Scholar] [CrossRef]
- Carvalho, A.K.F.; Bento, H.B.S.; Reis, C.E.R.; De Castro, H.F. Sustainable enzymatic approaches in a fungal lipid biorefinery based in sugarcane bagasse hydrolysate as carbon source. Bioresour. Technol. 2019, 276, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Gutarra, M.L.E.; Godoy, M.G.; Maugeri, F.; Rodrigues, M.I.; Freire, D.M.G.; Castilho, L.R. Production of an acidic and thermostable lipase of the mesophilic fungus Penicillium simplicissimum by solid-state fermentation. Bioresour. Technol. 2009, 100, 5249–5254. [Google Scholar] [CrossRef] [PubMed]
- Martin, N.; de Souza, S.R.; da Silva, R.; Gomes, E. Pectinase production by fungal strains in solid-state fermentation using agro-industrial bioproduct. Braz. Arch. Biol. Technol. 2004, 47, 813–819. [Google Scholar] [CrossRef]
- Nwe, N.; Chandrkrachang, S.; Stevens, W.F.; Maw, T.; Tan, T.K.; Khor, E.; Wong, S.M. Production of fungal chitosan by solid state and submerged fermentation. Carbohydr. Polym. 2002, 49, 235–237. [Google Scholar] [CrossRef]
- Colla, L.M.; Primaz, A.L.; Benedetti, S.; Loss, R.A.; de Lima, M.; Reinehr, C.O.; Bertolin, T.E.; Costa, J.A.V. Surface response methodology for the optimization of lipase production under submerged fermentation by filamentous fungi. Braz. J. Microbiol. 2016, 47, 461–467. [Google Scholar] [CrossRef]
- Wang, H.; Kaur, G.; Pensupa, N.; Uisan, K.; Du, C.; Yang, X.; Lin, C.S.K. Textile waste valorization using submerged filamentous fungal fermentation. Process Saf. Environ. Prot. 2018, 118, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Gmoser, R.; Sintca, C.; Taherzadeh, M.J.; Lennartsson, P.R. Combining submerged and solid state fermentation to convert waste bread into protein and pigment using the edible filamentous fungus N. intermedia. Waste Manag. 2019, 97, 63–70. [Google Scholar] [CrossRef]
- Viniegra-González, G.; Favela-Torres, E.; Aguilar, C.N.; de Jesus Rómero-Gomez, S.; Dıaz-Godınez, G.; Augur, C. Advantages of fungal enzyme production in solid state over liquid fermentation systems. Biochem. Eng. J. 2003, 13, 157–167. [Google Scholar] [CrossRef]
- Yang, J.; Jiao, R.H.; Yao, L.Y.; Han, W.B.; Lu, Y.H.; Tan, R.X. Control of fungal morphology for improved production of a novel antimicrobial alkaloid by marine-derived fungus Curvularia sp. IFB-Z10 under submerged fermentation. Process Biochem. 2016, 51, 185–194. [Google Scholar] [CrossRef]
- Veiter, L.; Rajamanickam, V.; Herwig, C. The filamentous fungal pellet—relationship between morphology and productivity. Appl. Microbiol. Biotechnol. 2018, 102, 2997–3006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karahalil, E.; Coban, H.B.; Turhan, I. A current approach to the control of filamentous fungal growth in media: Microparticle enhanced cultivation technique. Crit. Rev. Biotechnol. 2019, 39, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, A.; Antecka, A.; Owczarz, P.; Bizukojć, M. Inulinolytic activity of broths of Aspergillus niger ATCC 204447 cultivated in shake flasks and stirred tank bioreactor. Eng. Life Sci. 2017, 17, 1006–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimm, L.H.; Kelly, S.; Krull, R.; Hempel, D.C. Morphology and productivity of filamentous fungi. Appl. Microbiol. Biotechnol. 2005, 69, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Krull, R.; Wucherpfennig, T.; Esfandabadi, M.E.; Walisko, R.; Melzer, G.; Hempel, D.C.; Kampen, I.; Kwade, A.; Wittmann, C. Characterization and control of fungal morphology for improved production performance in biotechnology. J. Biotechnol. 2013, 163, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liao, W.; Chen, S. Study of pellet formation of filamentous fungi Rhizopus oryzae using a multiple logistic regression model. Biotechnol. Bioeng. 2008, 99, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Bizukojc, M.; Ledakowicz, S. Physiological, morphological and kinetic aspects of lovastatin biosynthesis by Aspergillus terreus. Biotechnol. J. 2009, 4, 647–664. [Google Scholar] [CrossRef] [Green Version]
- Grimm, L.H.; Kelly, S.; Völkerding, I.I.; Krull, R.; Hempel, D.C. Influence of mechanical stress and surface interaction on the aggregation of Aspergillus niger conidia. Biotechnol. Bioeng. 2005, 92, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhou, J.; Ding, Q.; Luo, Q.; Liu, L. Morphology engineering of Aspergillus oryzae for l-malate production. Biotechnol. Bioeng. 2019, 116, 2662–2673. [Google Scholar] [CrossRef]
- Lu, H.; Li, C.; Tang, W.; Wang, Z.; Xia, J.; Zhang, S.; Zhuang, Y.; Chu, J.; Noorman, H. Dependence of fungal characteristics on seed morphology and shear stress in bioreactors. Bioprocess Biosyst. Eng. 2015, 38, 917–928. [Google Scholar] [CrossRef]
- Lin, P.J.; Scholz, A.; Krull, R. Effect of volumetric power input by aeration and agitation on pellet morphology and product formation of Aspergillus niger. Biochem. Eng. J. 2010, 49, 213–220. [Google Scholar] [CrossRef]
- Cairns, T.C.; Zheng, X.; Zheng, P.; Sun, J.; Meyer, V. Moulding the mould: Understanding and reprogramming filamentous fungal growth and morphogenesis for next generation cell factories. Biotechnol. Biofuels 2019, 12, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Driouch, H.; Roth, A.; Dersch, P.; Wittmann, C. Optimized bioprocess for production of fructofuranosidase by recombinant Aspergillus niger. Appl. Microbiol. Biotechnol. 2010, 87, 2011–2024. [Google Scholar] [CrossRef] [PubMed]
- Driouch, H.; Sommer, B.; Wittmann, C. Morphology engineering of Aspergillus niger for improved enzyme production. Biotechnol. Bioeng. 2010, 105, 1058–1068. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Zeng, J.; Yu, X.; Dong, T.; Chen, S. Improved lipid accumulation by morphology engineering of oleaginous fungus Mortierella Isabellina. Biotechnol. Bioeng. 2014, 111, 1758–1766. [Google Scholar] [CrossRef] [PubMed]
- Coban, H.B.; Demirci, A.; Turhan, I. Enhanced Aspergillus ficuum phytase production in fed-batch and continuous fermentations in the presence of talcum microparticles. Bioprocess Biosyst. Eng. 2015, 38, 1431–1436. [Google Scholar] [CrossRef] [PubMed]
- Coban, H.B.; Demirci, A. Enhancement and modeling of microparticle-added Rhizopus oryzae lactic acid production. Bioprocess Biosyst. Eng. 2016, 39, 323–330. [Google Scholar] [CrossRef]
- Yatmaz, E.; Germec, M.; Karahalil, E.; Turhan, I. Enhancing β-mannanase production by controlling fungal morphology in the bioreactor with microparticle addition. Food Bioprod. Process. 2020, 121, 123–130. [Google Scholar] [CrossRef]
- Gürler, H.N.; Erkan, S.B.; Ozcan, A.; Yılmazer, C.; Karahalil, E.; Germec, M.; Yatmaz, E.; Ogel, Z.B.; Turhan, I. Scale-up processing with different microparticle agent for β-mannanase production in a large-scale stirred tank bioreactor. Proc. J. Food Process. Preserv. 2021, 45, e14915. [Google Scholar] [CrossRef]
- Germec, M.; Yatmaz, E.; Karahalil, E.; Turhan, I. Effect of different fermentation strategies on β-mannanase production in fed-batch bioreactor system. 3 Biotech 2017, 7, 77. [Google Scholar] [CrossRef] [Green Version]
- Singh, B. Engineering fungal morphology for enhanced production of hydrolytic enzymes by Aspergillus oryzae SBS50 using microparticles. 3 Biotech 2018, 8, 283. [Google Scholar] [CrossRef] [PubMed]
- Germec, M.; Karahalil, E.; Yatmaz, E.; Tari, C.; Turhan, I. Effect of process parameters and microparticle addition on polygalacturonase activity and fungal morphology of Aspergillus sojae. Biomass Convers. Biorefinery 2021, 12, 5329–5344. [Google Scholar] [CrossRef]
- Du, L.; Gao, B.; Liang, J.F.; Wang, Y.; Xiao, Y.; Zhu, D. Microparticle-enhanced Chaetomium globosum DX-THS3 β-d-glucuronidase production by controlled fungal morphology in submerged fermentation. 3 Biotech 2020, 10, 100. [Google Scholar] [CrossRef] [PubMed]
- Niu, K.; Wu, X.P.; Hu, X.L.; Zou, S.P.; Hu, Z.C.; Liu, Z.Q.; Zheng, Y.G. Effects of methyl oleate and microparticle-enhanced cultivation on echinocandin B fermentation titer. Bioprocess Biosyst. Eng. 2020, 43, 2009–2015. [Google Scholar] [CrossRef]
- Yue, C.; Xu, H.; Yu, Y.; Yu, X.; Yu, M.; Zhang, C.; You, Q.; Xia, S.; Ding, Z.; Fu, H. Improvement of natamycin production by controlling the morphology of Streptomyces gilvosporeus Z8 with microparticle talc in seed preculture. J. Chem. Technol. Biotechnol. 2021, 96, 1533–1542. [Google Scholar] [CrossRef]
- Huang, J.; Guan, H.W.; Huang, Y.Y.; Lai, K.S.; Chen, H.Y.; Xue, H.; Zhang, B.B. Evaluating the effects of microparticle addition on mycelial morphology, natural yellow pigments productivity, and key genes regulation in submerged fermentation of Monascus purpureus. Biotechnol. Bioeng. 2021, 118, 2503–2513. [Google Scholar] [CrossRef]
- Tong, L.L.; Wang, Y.; Du, Y.H.; Yuan, L.; Liu, M.Z.; Mu, X.Y.; Chen, Z.L.; Zhang, Y.D.; He, S.J.; Li, X.J.; et al. Transcriptomic Analysis of Morphology Regulatory Mechanisms of Microparticles to Paraisaria dubia in Submerged Fermentation. Appl. Biochem. Biotechnol. 2022, 194, 4333–4347. [Google Scholar] [CrossRef]
- Tong, L.L.; Wang, Y.; Yuan, L.; Liu, M.Z.; Du, Y.H.; Mu, X.Y.; Yang, Q.H.; Wei, S.X.; Li, J.Y.; Wang, M.; et al. Enhancement of polysaccharides production using microparticle enhanced technology by Paraisaria dubia. Microb. Cell Factories 2022, 21, 12. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.; Chen, Y.; Xiao, N. Effects of metal oxide nanoparticles (TiO2, Al2O3, SiO2 and ZnO) on waste activated sludge anaerobic digestion. Bioresour. Technol. 2011, 102, 10305–10311. [Google Scholar] [CrossRef]
- Dong, M.; Wang, S.; Xu, F.; Li, Q.; Li, W. Addition of aluminum oxide microparticles to Trichoderma viride My preculture enhances cellulase production and influences fungal morphology. Eng. Life Sci. 2018, 18, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Boruta, T.; Antecka, A. Co-cultivation of filamentous microorganisms in the presence of aluminum oxide microparticles. Appl. Microbiol. Biotechnol. 2022, 106, 5459–5477. [Google Scholar] [CrossRef] [PubMed]
- Antecka, A.; Blatkiewicz, M.; Bizukojć, M.; Ledakowicz, S. Morphology engineering of basidiomycetes for improved laccase biosynthesis. Biotechnol. Lett. 2016, 38, 667–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Jin, Y.; Wang, Z.; Xing, D.; Ma, C.; Ding, J.; Ren, N. Enhanced photo-fermentative hydrogen production of Rhodopseudomonas sp. nov. strain A7 by the addition of TiO2, ZnO and SiC nanoparticles. Int. J. Hydrogen Energy 2017, 42, 18279–18287. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Z.; He, X.; Zheng, L.; Cheng, S.; Li, Z. Diminished inhibitory impact of ZnO nanoparticles on anaerobic fermentation by the presence of TiO2 nanoparticles: Phenomenon and mechanism. Sci. Total Environ. 2019, 647, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhang, Z.; Tian, L.; Zhang, L.; Cheng, S.; Li, Z.; Cang, D. Mechanistic investigation of toxicological change in ZnO and TiO2 multi-nanomaterial systems during anaerobic digestion and the microorganism response. Biochem. Eng. J. 2019, 147, 62–71. [Google Scholar] [CrossRef]
- Zhang, T.; Jiang, D.; Li, Y.; Zhang, H.; Zhang, Z.; Petracchini, F.; Paolini, V.; Zhang, Y.; Yue, T.; Zhang, Q. Study of the interrelationship between nano-TiO2 addition and photo-fermentative bio-hydrogen production of corn straw. Bioresour. Technol. 2021, 338, 125549. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Wei, W.; Ni, B.J. Revealing the mechanism of zinc oxide nanoparticles facilitating hydrogen production in alkaline anaerobic fermentation of waste activated sludge. J. Clean. Prod. 2021, 328, 129580. [Google Scholar] [CrossRef]
- Jin, B.; Yuan, Y.; Zhou, P.; Niu, J.; Niu, J.; Dai, J.; Li, N.; Tao, H.; Ma, Z.; Zhang, J.; et al. Effects of zinc oxide nanoparticles on sludge anaerobic fermentation: Phenomenon and mechanism. J. Environ. Sci. Health Part A Toxic/Hazard. Subst. Environ. Eng. 2020, 55, 1094–1103. [Google Scholar] [CrossRef]
- Driouch, H.; Hänsch, R.; Wucherpfennig, T.; Krull, R.; Wittmann, C. Improved enzyme production by bio-pellets of Aspergillus niger: Targeted morphology engineering using titanate microparticles. Biotechnol. Bioeng. 2012, 109, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Hu, T.; Liu, H.; Xie, H.; Tian, X.; Wu, Z. Biosynthesis and polyketide oxidation of Monascus red pigments in an integrated fermentation system with microparticles and surfactants. Food Chem. 2022, 394, 133545. [Google Scholar] [CrossRef]
- Etschmann, M.M.W.; Huth, I.; Walisko, R.; Schuster, J.; Krull, R.; Holtmann, D.; Schrader, J. Improving 2-phenylethanol and 6-pentyl-α-pyroneproduction with fungi by microparticle-enhancedcultivation (MPEC). Yeast 2015, 32, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Karahalil, E.; Demirel, F.; Evcan, E.; Germeç, M.; Tari, C.; Turhan, I. Microparticle-enhanced polygalacturonase production by wild type Aspergillus sojae. 3 Biotech 2017, 7, 361. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, M.; Rückert, C.; Gläser, L.; Beganovic, S.; Luzhetskyy, A.; Kalinowski, J.; Wittmann, C. Microparticles enhance the formation of seven major classes of natural products in native and metabolically engineered actinobacteria through accelerated morphological development. Biotechnol. Bioeng. 2021, 118, 3076–3093. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Carbon Source | Fungal Strain(S) | Value-Added Products | References |
|---|---|---|---|
| Spent hydrolysates | Trichoderma reesei Rut C-30 | Enzymes | [15] |
| Mixture of dry leaves | Aspergillus niger | Cellulase | [16] |
| Aguamiel | Aspergillus niger GH1, Aspergillus niger PSH and Aspergillus oryzae DIA-MF | Fructooligosaccharides (FOS) | [17] |
| Whole maize flour | Aspergillus niger | Citric acid | [18] |
| Corn cob and sawdust | Aspergillus niger | Xylanase | [19] |
| Corn cobs | Aspergillus flavus AW1 | Xylanase | [20] |
| Sorghum xylans | Aspergillus fumigatus RSP-8 | Xylanase | [21] |
| Algerian date varieties | Aspergillus niger ATCC 16888 | Citric acid | [22] |
| Rice husk, cottonseed cake, and red gram husk | Aspergillus niger MTCC 872 | Lipases | [23] |
| Distillers dried grains with solubles (DDGS) | Aspergillus niger (NRRL 330) and (NRRL 567) | Cellulases and hemicellulases | [24] |
| Sunflower stalks | Aspergillus sp. | Xylanase | [25] |
| Sugarcane bagasse | Mucor circinelloides | Single cell oil | [26] |
| Babassu cake | Penicillium simplicissimum | Lipase | [27] |
| Agro-industrial bioproducts | Moniliella SB9 and Penicillium sp. EGC5 | Polygalcturonase and pectin lyase | [28] |
| Sweet potato extract | Gongronella butleri USDB 0201 | Chitosan | [29] |
| Wheat and soybean bran | Aspergillus niger and Aspergillus flavus | Lipase | [30] |
| Textile waste | Trichoderma reesei ATCC 24449 | Cellulase | [31] |
| Waste streams from ethanol and bread | Neurospora intermedia | Ethanol, biomass and a feed product | [32] |
| Microparticle | Microorganism | Product | Reference |
|---|---|---|---|
| Talc | C. fumago | Chloroperoxidase | [10] |
| A. niger | Fructofuranosidase | [47] | |
| Enzyme Production | [48] | ||
| Inulinolytic activity | [37] | ||
| M. isabellina | Lipid production | [49] | |
| A. ficuum | Phytase | [9,50] | |
| R. oryzae | Lactic acid | [51] | |
| A. sojae | β-mannanase | [7,52,53,54] | |
| Hydrolytic enzymes | [55] | ||
| Polygalacturonase | [56] | ||
| C. globosum | β-d-glucuronidase | [57] | |
| A. nidulans | Echinocandin B | [58] | |
| S. gilvosporeus | Natamycin | [59] | |
| M. purpureus | Yellow pigments | [60] | |
| P. dubia | Morphology regulatory | [61] | |
| Polysaccharides | [62] | ||
| Al-oxide | C. fumago | Chloroperoxidase | [10] |
| A. niger | Enzyme Production | [48] | |
| A. ficuum | Phytase | [9,50] | |
| R. oryzae | Lactic acid | [51] | |
| A. sojae | β-mannanase | [7,52,53,54] | |
| Hydrolytic enzymes | [55] | ||
| Polygalacturonase | [56] | ||
| M. purpureus | Yellow pigments | [60] | |
| P. dubia | Morphology regulatory | [61] | |
| Activated sludge anaerobic digestion | [63] | ||
| Trichoderma viride | Cellulase | [64] | |
| C. globosum | β-d-glucuronidase | [57] | |
| A. terreus co-culture | Lovastatin | [65] | |
| Penicillin G | [65] | ||
| Oxytetracycline | [65] | ||
| Laccase | [65] | ||
| C. unicolor | Laccase | [66] | |
| P. sapidus | Laccase | [66] | |
| SiO2 | M. purpureus | Yellow pigments | [60] |
| Activated sludge anaerobic digestion | [63] | ||
| TiO2 | M. purpureus | Yellow pigments | [60] |
| Activated sludge anaerobic digestion | [39] | ||
| Rhodopseudomonas sp. | Hydrogen production | [67] | |
| Sludge anaerobic fermentation | [68,69] | ||
| HAU-M1 | Hydrogen production | [70] | |
| ZnO | Activated sludge anaerobic digestion | [63] | |
| Rhodopseudomonas sp. | Hydrogen production | [67,71] | |
| Sludge anaerobic fermentation | [68,69,72] | ||
| Titanium silicone oxide | A. niger | Enzyme Production | [73] |
| OSA-SNa | M. purpureus | Monascus red pigments | [74] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iram, A.; Özcan, A.; Yatmaz, E.; Turhan, İ.; Demirci, A. Effect of Microparticles on Fungal Fermentation for Fermentation-Based Product Productions. Processes 2022, 10, 2681. https://doi.org/10.3390/pr10122681
Iram A, Özcan A, Yatmaz E, Turhan İ, Demirci A. Effect of Microparticles on Fungal Fermentation for Fermentation-Based Product Productions. Processes. 2022; 10(12):2681. https://doi.org/10.3390/pr10122681
Chicago/Turabian StyleIram, Attia, Ali Özcan, Ercan Yatmaz, İrfan Turhan, and Ali Demirci. 2022. "Effect of Microparticles on Fungal Fermentation for Fermentation-Based Product Productions" Processes 10, no. 12: 2681. https://doi.org/10.3390/pr10122681
APA StyleIram, A., Özcan, A., Yatmaz, E., Turhan, İ., & Demirci, A. (2022). Effect of Microparticles on Fungal Fermentation for Fermentation-Based Product Productions. Processes, 10(12), 2681. https://doi.org/10.3390/pr10122681