
The Effect of Spray Parameters on the Survival of Bacteriophages


Abstract
:1. Introduction
2. Materials and Methods
2.1. Spray Machine Parameter Selection
2.2. PGS Diluted to 1% with Prepared Bottled Water
2.3. PGS Titration and Renumeration
SM Buffer Preparation
2.4. Luria Bertani (LB) Broth
2.5. Luria Bertani (LB) Agar Plates for Titration of PGS
2.6. Luria Bertani (LB) Top Agar Plates for Titration of PGS
2.7. Salmonella: Propagation of Culture
2.8. Overnight Culture for Glycerol Stocks
2.9. Phage Enumeration via Titration of PGS Samples
2.10. Phage Renumeration
2.11. PGS Diluted to 1% with Distilled Water
2.12. Statistical Analysis
3. Results
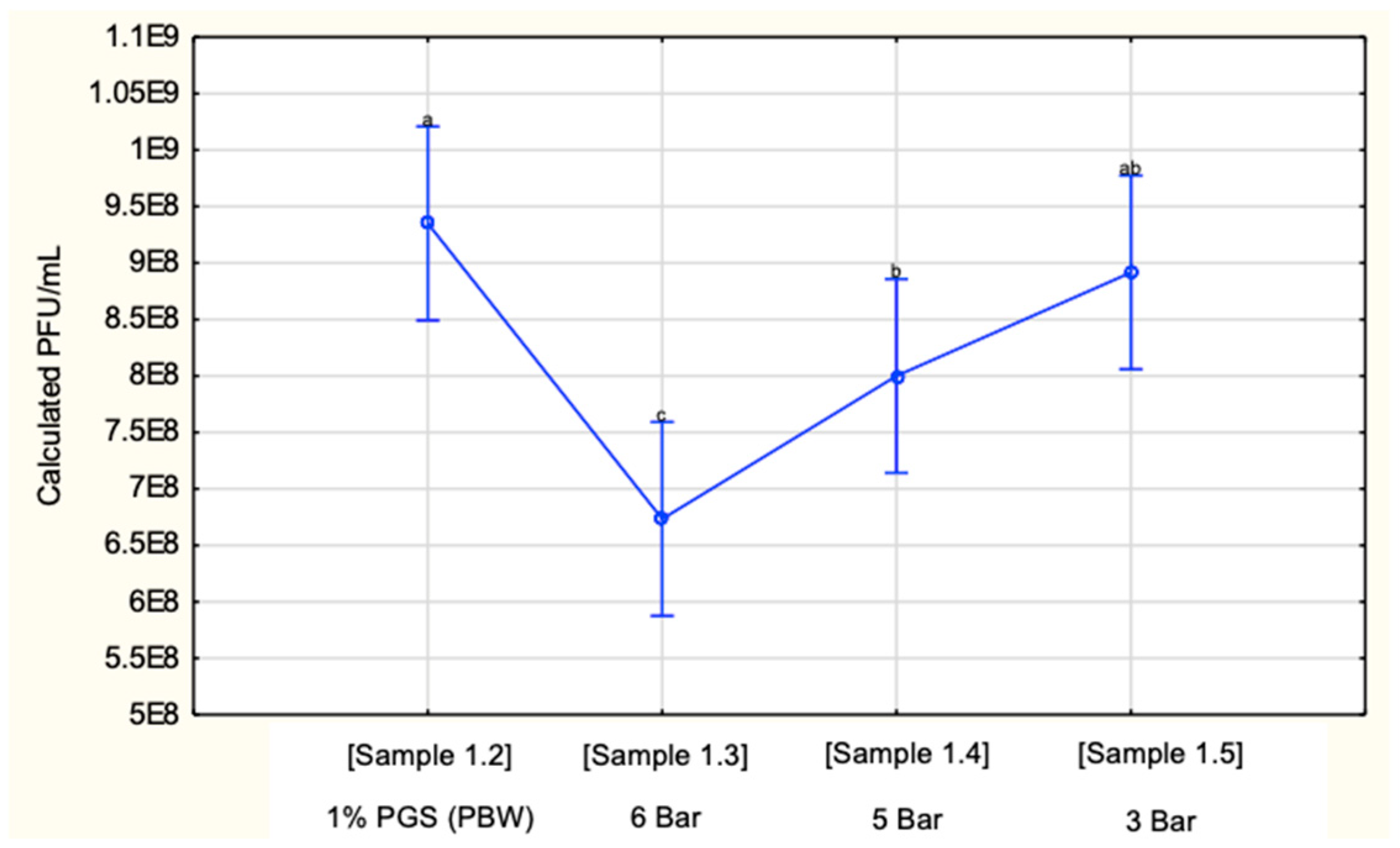
3.1. PGS Diluted to 1% with PBW
3.2. PGS Diluted to 1% with dH2O
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Hagens, S.; Loessner, M.J. Bacteriophage for biocontrol of foodborne pathogens: Calculations and considerations. Curr. Pharm. Biotechnol. 2010, 11, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Endersen, L.; Coffey, A. The use of bacteriophages for food safety. Curr. Opin. Food Sci. 2020, 36, 1–8. [Google Scholar] [CrossRef]
- Wessels, K.; Rip, D.; Gouws, P. Salmonella in Chicken Meat: Consumption, Outbreaks, Characteristics, Current Control Methods and the Potential of Bacteriophage Use. Foods. Foods 2021, 10, 1742. [Google Scholar] [CrossRef] [PubMed]
- Berry, E.M.; Dernini, S.; Burlingame, B.; Meybeck, A.; Conforti, P. Food security and sustainability: Can one exist without the other? Public Health Nutr. 2015, 18, 2293–2302. [Google Scholar] [CrossRef]
- Thapa, S.P.; Shrestha, S.; Anal, A.K. Addressing the antibiotic resistance and improving the food safety in food supply chain (farm-to-fork) in Southeast Asia. Food Control 2020, 108, 106809. [Google Scholar] [CrossRef]
- Shannon, R.; Radford, D.R.; Balamurugan, S. Impacts of food matrix on bacteriophage and endolysin antimicrobial efficacy and performance. Crit. Rev. Food Sci. Nutr. 2020, 60, 1631–1640. [Google Scholar] [CrossRef] [PubMed]
- Moye, Z.D.; Woolston, J.; Sulakvelidze, A. Bacteriophage applications for food production and processing. Viruses 2018, 10, 205. [Google Scholar] [CrossRef] [Green Version]
- De Melo, A.G.; Levesque, S.; Moineau, S. Phages as friends and enemies in food processing. Curr. Opin. Biotechnol. 2018, 49, 185–190. [Google Scholar] [CrossRef]
- Yeh, Y.; Purushothaman, P.; Gupta, N.; Ragnone, M.; Verma, S.C.; De Mello, A.S. Bacteriophage application on red meats and poultry: Effects on Salmonella population in final ground products. Meat Sci. 2017, 127, 30–34. [Google Scholar]
- Huang, C.; Shi, J.; Ma, W.; Li, Z.; Wang, J.; Li, J.; Wang, X. Isolation, characterization, and application of a novel specific Salmonella bacteriophage in different food matrices. Food Res. Int. 2018, 111, 631–641. [Google Scholar] [CrossRef]
- Ahmadi, H.; Barbut, S.; Lim, L.T.; Balamurugan, S. Examination of the use of bacteriophage as an additive and determining its best application method to control Listeria monocytogenes in a cooked-meat model system. Front. Microbiol. 2020, 11, 779. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. Food Additives Permitted for Direct Addition to Food for Human Consumption: Bacteriophage Preparation. 2006. Available online: http://edocket.access.gpo.gov/2006/E6-13621.htm (accessed on 22 May 2021).
- Health Canada. Policy on Listeria Monocytogenes in Ready-To-Eat Foods, Identification, Number: FD-FSNP 0071. 2011. Available online: http://www.hc-sc.gc.ca/fn-an/legislation/pol/policy_listeria_monocytogenes_2011-eng.php (accessed on 22 May 2021).
- Reinhard, R.G.; Kalinowski, R.M.; Bodnaruk, P.W.; Eifert, J.D.; Boyer, R.R.; Duncan, S.E.; Bailey, R.H. Practical application of bacteriophage in food manufacturing facilities for the control of Listeria sp. J. Food Saf. 2020, e12871. [Google Scholar] [CrossRef]
- Soffer, N.; Woolston, J.; Li, M.; Das, C.; Sulakvelidze, A. Bacteriophage preparation lytic for Shigella significantly reduces Shigella sonnei contamination in various foods. PLoS ONE 2017, 12, e0175256. [Google Scholar]
- Sukumaran, A.T.; Nannapaneni, R.; Kiess, A.; Sharma, C.S. Reduction of Salmonella on chicken meat and chicken skin by combined or sequential application of lytic bacteriophage with chemical antimicrobials. Int. J. Food Microbiol. 2015, 207, 8–15. [Google Scholar] [CrossRef]
- Jończyk-Matysiak, E.; Łodej, N.; Kula, D.; Owczarek, B.; Orwat, F.; Międzybrodzki, R.; Neuberg, J.; Bagińska, N.; Weber-Dąbrowska, B.; Górski, A. Factors determining phage stability/activity: Challenges in practical phage application. Expert Rev. Anti-Infect. Ther. 2019, 17, 583–606. [Google Scholar] [CrossRef]
- Fister, S.; Robben, C.; Witte, A.K.; Schoder, D.; Wagner, M.; Rossmanith, P. Influence of environmental factors on phage–bacteria interaction and on the efficacy and infectivity of phage P100. Front. Microbiol. 2016, 7, 1152. [Google Scholar] [CrossRef]
- Brié, A.; Gantzer, C.; Boudaud, N.; Bertrand, I. The impact of chlorine and heat on the infectivity and physicochemical properties of bacteriophage MS2. FEMS Microbiol. Ecol. 2018, 94, fiy106. [Google Scholar] [CrossRef]
- González-Menéndez, E.; Arroyo-López, F.N.; Martínez, B.; García, P.; Garrido-Fernández, A.; Rodríguez, A. Optimizing propagation of Staphylococcus aureus infecting bacteriophage vB_sauM-phiIPLA-RODI on Staphylococcus xylosus using response surface methodology. Viruses 2018, 10, 153. [Google Scholar] [CrossRef] [Green Version]
- Chaudhry, W.N.; Concepcion-Acevedo, J.; Park, T.; Andleeb, S.; Bull, J.J.; Levin, B.R. Synergy and order effects of antibiotics and phages in killing Pseudomonas aeruginosa biofilms. PLoS ONE 2017, 12, e0168615. [Google Scholar]
- Langlet, J.; Gaboriaud, F.; Gantzer, C. Effects of pH on plaque forming unit counts and aggregation of MS2 bacteriophage. J. Appl. Microbiol. 2007, 103, 1632–1638. [Google Scholar] [CrossRef]
- Mylon, S.E.; Rinciog, C.I.; Schmidt, N.; Gutierrez, L.; Wong, G.C.; Nguyen, T.H. Influence of salts and natural organic matter on the stability of bacteriophage MS2. Langmuir 2010, 26, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Wigginton, K.R.; Pecson, B.M.; Sigstam, T.; Bosshard, F.; Kohn, T. Virus inactivation mechanisms: Impact of disinfectants on virus function and structural integrity. Environ. Sci. Technol. 2012, 46, 12069–12078. [Google Scholar] [CrossRef] [PubMed]
- Sano, D.; Ohta, T.; Nakamura, A.; Nakagomi, T.; Nakagomi, O.; Okabe, S. Culture-independent evaluation of nonenveloped-virus infectivity reduced by free-chlorine disinfection. Appl. Environ. Microbiol. 2015, 81, 2819–2826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loison, P.; Majou, D.; Gelhaye, E.; Boudaud, N.; Gantzer, C. Impact of reducing and oxidizing agents on the infectivity of Qβ phage and the overall structure of its capsid. FEMS Microbiol. Ecol. 2016, 92, fiw153. [Google Scholar] [CrossRef] [Green Version]
- Bloomfield, S.F.; Smith-Burchnell, C.A.; Dalgleish, A.G. Evaluation of hypochlorite-releasing disinfectants against the human immunodeficiency virus (HIV). J. Hosp. Infect. 1990, 15, 273–278. [Google Scholar] [CrossRef]
- Mansour, N. Bacteriophages are a natural gift, could we pay further attention! J. Food Microbiol. 2017, 1, 1–22. [Google Scholar]
- Marti, R.; Zurfluh, K.; Hagens, S.; Pianezzi, J.; Klumpp, J.; Loessner, M.J. Long tail fibres of the novel broad-host-range T-even bacteriophage S 16 specifically recognize Salmonella OmpC. Mol. Microbiol. 2013, 87, 818–834. [Google Scholar] [CrossRef]
- Whichard, J.M.; Weigt, L.A.; Borris, D.J.; Li, L.L.; Zhang, Q.; Kapur, V.; Pierson, F.W.; Lingohr, E.J.; She, Y.M.; Kropinski, A.M.; et al. Complete genomic sequence of bacteriophage Felix O1. Viruses 2010, 2, 710–730. [Google Scholar] [CrossRef]
- Loh, B.; Kuhn, A.; Leptihn, S. The fascinating biology behind phage display: Filamentous phage assembly. Mol. Microbiol. 2019, 111, 1132–1138. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Concentration (ppm) |
|---|---|
| Salt | 0.007 |
| Potassium | 0.5 |
| Chloride | 4 |
| Calcium | 0.62 |
| Magnesium | 0.5 |
| Fluoride | 0.1 |
| Sulphate | 1 |
| Calcium carbonate | 0.8 |
| Nitrate | 0.89 |
| Sample Information | Dilution | Pump Pressure (Bar) |
|---|---|---|
| Sample 1.1 10 mL PGS stock | 100% | N/A |
| Sample 1.2 25 mL 1% PGS pre-spray | 1 % | N/A |
| Sample 1.3 25 mL 1% PGS post-spray | 1% | 6 |
| Sample 1.4 25 mL 1% PGS post-spray | 1% | 5 |
| Sample 1.5 25 mL 1% PGS post-spray | 1% | 3 |
| PBW | dH2O | ||
|---|---|---|---|
| Sample | % Survival | Sample | % Survival |
| 1.1 (stock) | 100 | 2.1 (stock) | 100.00 |
| 1.2 (1%) | 33.16 | 2.2 (1%) | 88.16 |
| 1.3 (6 Bar) | 23.88 | 2.3 (6 Bar) | 63.74 |
| 1.4 (5 Bar) | 28.37 | 2.4 (5 Bar) | 77.10 |
| 1.5 (3 Bar) | 31.62 | 2.5 (3 Bar) | 85.50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wessels, K.; Rip, D.; Gouws, P. The Effect of Spray Parameters on the Survival of Bacteriophages. Processes 2022, 10, 673. https://doi.org/10.3390/pr10040673
Wessels K, Rip D, Gouws P. The Effect of Spray Parameters on the Survival of Bacteriophages. Processes. 2022; 10(4):673. https://doi.org/10.3390/pr10040673
Chicago/Turabian StyleWessels, Kirsten, Diane Rip, and Pieter Gouws. 2022. "The Effect of Spray Parameters on the Survival of Bacteriophages" Processes 10, no. 4: 673. https://doi.org/10.3390/pr10040673
APA StyleWessels, K., Rip, D., & Gouws, P. (2022). The Effect of Spray Parameters on the Survival of Bacteriophages. Processes, 10(4), 673. https://doi.org/10.3390/pr10040673